
Abstract
Global warming and extreme weather events pose a significant threat to global biodiversity, with rising water temperatures exerting a profound influence on fish conservation and fishery development. In this study, we used zebrafish as a model organism to explore the impact of a heat acclimation period on their survival rates. The results demonstrated that a 2-month heat acclimation period almost completely mitigated heat stress-induced mortality in zebrafish. Subsequent analysis of the surviving zebrafish revealed a predominance of hepatic mitochondria in a fission state. Remarkably, a short-term fasting regimen, which induced hepatic mitochondrial fission, mirrored the outcomes of the protective effect of heat acclimation and augmented animal survival under heat stress. Conversely, treatment with a mitochondrial fission inhibitor within the fasting group attenuated the elevated survival rate. Furthermore, zebrafish embryos subjected to brief heat acclimation also exhibited increased heat resistance, a trait diminished by a chemical intervention inhibiting mitochondrial fission. This suggests a shared mechanism for heat resistance between embryos and adult zebrafish. These findings underscore the potential use of inducing mitochondrial fission to enhance heat resistance in zebrafish, offering promise for fish biodiversity conservation in the face of global warming.
Introduction
Global warming, characterized by the progressive increase in the Earth’s surface temperature over the past century primarily due to the emission of greenhouse gases from fossil fuel use, 1 presents an impending challenge. Current projections anticipate a 5°C temperature rise by the end of this century,2,3 with the climate nearing an irreversible tipping point. 4 Climate change has adversely affected biodiversity, impacting ecosystem structure and function.5,6 In addition, climate-driven alterations in fishery production and fish species composition pose substantial economic losses. 7 In addition to the gradual temperature increase, short-term extreme heat events, known as heatwaves, are becoming more prevalent during summers, directly endangering fish populations. 8
Heat stress significantly influences fish physiology, increasing cortisol levels and perturbing normal physiological functions.9–11 It leads to the overproduction of reactive oxygen species (ROS), resulting in oxidative stress and damage to biological macromolecules.12,13 Furthermore, heat stress compromises the immune system by upregulating proinflammatory cytokine expression14–16 and, under extreme conditions, can activate apoptosis-related pathways, leading to cell death.8,16 Consequently, fish have developed mechanisms to enhance heat resistance. The antioxidant system is activated to produce more antioxidant enzymes, protecting cells from ROS-induced damage caused by heat stress.17,18 In addition, molecular chaperones, such as heat–shock proteins, play a vital role in restoring the structure of heat-damaged proteins.17,19 However, these protective mechanisms have limitations, as sustained high temperatures or acute heat stress can lead to irreversible fish body damage.17,18
Heat acclimation is known to enhance heat resistance to elevated temperatures, with previous studies examining its effects on gene expression, immunity, and heat rate.20–23 Nevertheless, the detailed mechanism underlying heat resistance resulting from heat acclimation remains incompletely understood. A recent report investigates the role of mitochondrial fission in long-term heat tolerance in Arabidopsis, highlighting the discovery of the SLOH 5 mutant hypersensitive to long-term heat stress owing to massively elongated mitochondria, indicating the significance of mitochondrial dynamics in plant heat stress response. 24 In addition, dynamin-related protein 1 (DRP-1)-dependent mitochondrial fission and autophagosome biogenesis can enhance Caenorhabditis elegans’ resilience to acute heat stress by promoting mitochondrial rebuilding. 25 Whether a similar mechanism exists in fish undergoing heat acclimation and protecting them from heat stress remains to be elucidated.
This study used zebrafish to investigate the impact of heat acclimation on their survival rates. We found that heat acclimation entirely prevented heat stress-induced mortality in zebrafish. Notably, heat acclimation was associated with a significant shift in mitochondrial morphology toward a fission state in the zebrafish liver. Consequently, short-term fasting recapitulated mitochondrial fission phenotype by heat acclimation and significantly increased the survival rate of zebrafish exposed to heat stress. Moreover, interventions with chemicals inhibiting mitochondrial fission markedly reduced fish survival, highlighting the critical role of mitochondrial dynamics in regulating heat resistance in zebrafish.
Materials and Methods
Ethical approval
The zebrafish experiments conducted in this study were approved by the ethics committee for animal experiments at Wonkwang University (approval code: WKU21-25) and adhered to animal experiment guidelines.
Zebrafish experiments
Zebrafish (Danio rerio) of the Tg(Xla.Eef1a1:mlsEGFP) transgenic line were used to study mitochondrial morphology in the liver, 26 whereas wild-type zebrafish were used for other experiments. Standard protocols were followed for raising and maintaining zebrafish in a controlled environment with a 14-h light/10-h dark cycle at 28.5°C. 27 Embryos were collected, developed in egg water, and staged according to standard procedures. 28 During experimental treatments, adult fish were housed in transparent 2-L tanks with a maximum of five fish per tank. Unless otherwise specified, fish in each group received fresh artemia as feed twice a day. A thermostable water bath (ThermoFisher Scientific, Waltham, MA, USA) was used for tank placement and temperature maintenance, as indicated in each experiment. All experiments were conducted at least twice.
Heat acclimation and heat tolerance in adult zebrafish
Adult zebrafish at 3.5 months postfertilization (mpf) were randomly assigned to either the heat acclimation or the control group. The heat acclimation condition involved exposure to 33°C (moderate heat stress) for 2 months, based on preliminary data and previous reports. 29 The control group was maintained at 28.5°C. After the 2-month temperature exposure, zebrafish were assessed for physiological parameters and then randomly divided into two groups: one for a heat tolerance test at 39°C and the other for examining mitochondrial morphology or performing quantitative polymerase chain reaction (qPCR).
Chloroquine treatment
Adult zebrafish at 3.5 mpf were randomly allocated to three groups: control (fed), fast, and fast + chloroquine (CQ) group. CQ was obtained from Sigma-Aldrich (St. Louis, MO, USA). All fish were maintained at 28.5°C. The fed group received fresh artemia twice daily, whereas the fasted group abstained from food for 2 days. The fast + CQ group was administered 0.5 mM CQ for 2 days. After 2 days of treatment, fish were either sacrificed for cryosectioning or subjected to heat tolerance tests.
Short-term heat acclimation in embryos
Embryos from a natural mating were divided into four experimental groups and exposed to 33°C heat stress for different durations from 0 to 4, 1 to 4, 2 to 4, or 3 to 4 days postfertilization (dpf). Sibling embryos developed at 28.5°C served as the control group. At 4 dpf, embryos were processed for cryosectioning or subjected to a heat tolerance test at 39°C.
Inhibition of mitochondrial fission in embryos
Random allocation of embryos was performed into one of three experimental groups: control, heat acclimation, and heat acclimation with the addition of Mdivi-1 (Sigma-Aldrich), a mitochondrial division inhibitor. From 2 to 4 dpf, the heat acclimation group was subjected to an elevated temperature of 33°C. The heat acclimation + Mdivi-1 group received a treatment of 5 μM Mdivi-1 and was concurrently exposed to the elevated temperature of 33°C from 2 to 4 dpf. The control group was maintained at a temperature of 28.5°C without any intervention. At the conclusion of the 4 dpf period, larvae were either sacrificed for subsequent cryosectioning or used in a heat tolerance assessment conducted at 39°C.
Measurement of body mass index
Standard body length (SL), defined as the distance spanning from the tip of the snout to the caudal peduncle, was quantified using a ruler. Body weight (W) was measured using the GX-400 milligram scale (A&D Weighing, Japan). Body mass index (BMI) was calculated using the formula: BMI = W (mg) / SL (cm)2.
Blood glucose measurement
The blood glucose measurement was performed in accordance with previously established protocols. 30 The fish were subjected to anesthesia in ice-cold water, 8 hours subsequent to their most recent feeding, and blood glucose levels were assessed using one-touch ultra-test strips (Roche, Basel, Switzerland).
RNA extraction and qPCR
Total RNA was extracted from the liver tissue of zebrafish specimens using Trizol (Ambion, ThermoFisher Scientific). Subsequently, first-strand cDNA synthesis was carried out using the Roche kit, and qPCR was conducted using equipment from Agilent Technologies (Santa Clara, CA, USA), in accordance with previously described procedures. 29 The reference gene used was β-actin. The primer sequences used in this study are provided in Table 1.
Sequence Information of Primers
qPCR, quantitative polymerase chain reaction.
Cryosectioning and imaging
Cryosectioning procedures were executed in accordance with previously documented protocols. 31 Briefly, anesthetized zebrafish were subjected to overnight fixation in a 4% paraformaldehyde solution. Subsequently, they were embedded in 5% sucrose and 1.5% agarose gel, followed by incubation in a 30% sucrose solution overnight at 4°C. The samples were then exposed in a liquid nitrogen environment to 2-methylbutane (Junsei Chemical, Tokyo, Japan) to form final frozen blocks. A cryomicrotome HX525 NX (ThermoFisher Scientific) was used to section the cryopreserved blocks. These sliced specimens were affixed onto ultra-frost microscope slides, air-dried for 30 min, and subsequently used for imaging. The mitochondrial morphology within liver sections was visualized using a confocal microscope (Olympus IX81 Fluoview FV1000, Olympus, Tokyo, Japan) at the Core Facility for Supporting Analysis and Imaging of Biomedical Materials at Wonkwang University, which receives support from the National Research Facilities and Equipment Center.
Heat tolerance assessment
Each zebrafish group was subjected to an elevated temperature of 39°C, and the cumulative hourly mortality was meticulously recorded. Deceased zebrafish were promptly removed from the tank and handled in accordance with established animal experimentation guidelines. Survival rate comparisons among distinct groups were analyzed using GraphPad Prism 8 software (GraphPad, Boston, MA, USA).
Quantification of cells with fragmented mitochondria
Images were taken by Olympus FluoView 1000 confocal laser scanning system (Olympus), and the imaging was edited by Photoshop CS6 (Adobe). For determining fragmented mitochondria, ImageJ software (National Institutes of Health) was used. In a given cell of an image field, the number of mitochondria with length <3 μm was manually determined. Cells that did not contain mitochondria with >3 μm were categorized as cells with “fragmented” mitochondria. The percentage of cells with fragmented mitochondria was determined by observing at least 13 cells/sample, each from a separate image field and from at least three independent experiments. *indicates p < 0.05, **p < 0.01, and ***p < 0.001.
Statistical analyses
All reported values are presented as the mean ± standard deviation, except in the case of qPCR results, for which standard errors of the mean were used. Levels of RNA expression were normalized relative to β-actin expression, with the gene expression in the control group being designated as 1.0. Statistical significance was determined via the Student’s unpaired t-test using GraphPad Prism 8 software, with a predefined significance threshold of p < 0.05, p < 0.01, or p < 0.001.
Results
Physiological alterations in zebrafish induced by heat acclimation
The phenomenon of heat acclimation has consistently demonstrated its efficacy in enhancing thermal tolerance across diverse species.32–34 In a prior investigation, zebrafish accustomed to their standard habitat temperature of 28.5°C exhibited rapid mortality when subjected to an elevated temperature of 39°C. 29 In the current study, we imposed a heat acclimation regimen on zebrafish at 33°C for a duration of 2 months. As a consequence of heat acclimation, the heat-acclimated group exhibited no significant increase in W or BMI compared with the control group (Fig. 1A, B). In addition, the heat-acclimated zebrafish displayed a marked reduction in blood glucose levels compared to the control group (Fig. 1C). Importantly, the heat-acclimated zebrafish displayed remarkable enhancement in their ability to endure high temperatures, notably enduring a 5-h exposure at 39°C, whereas the control group died during the heat tolerance assessment (n = 20 per group, 2 replicates). Given that heat-induced fish mortality is frequently associated with hepatic damage, 8 and considering the pivotal role of mitochondrial function in high-temperature acclimation in other fish species such as salmon, 35 it is plausible that the physiological changes observed following heat acclimation may be linked to alterations within the mitochondrial population of the zebrafish liver.
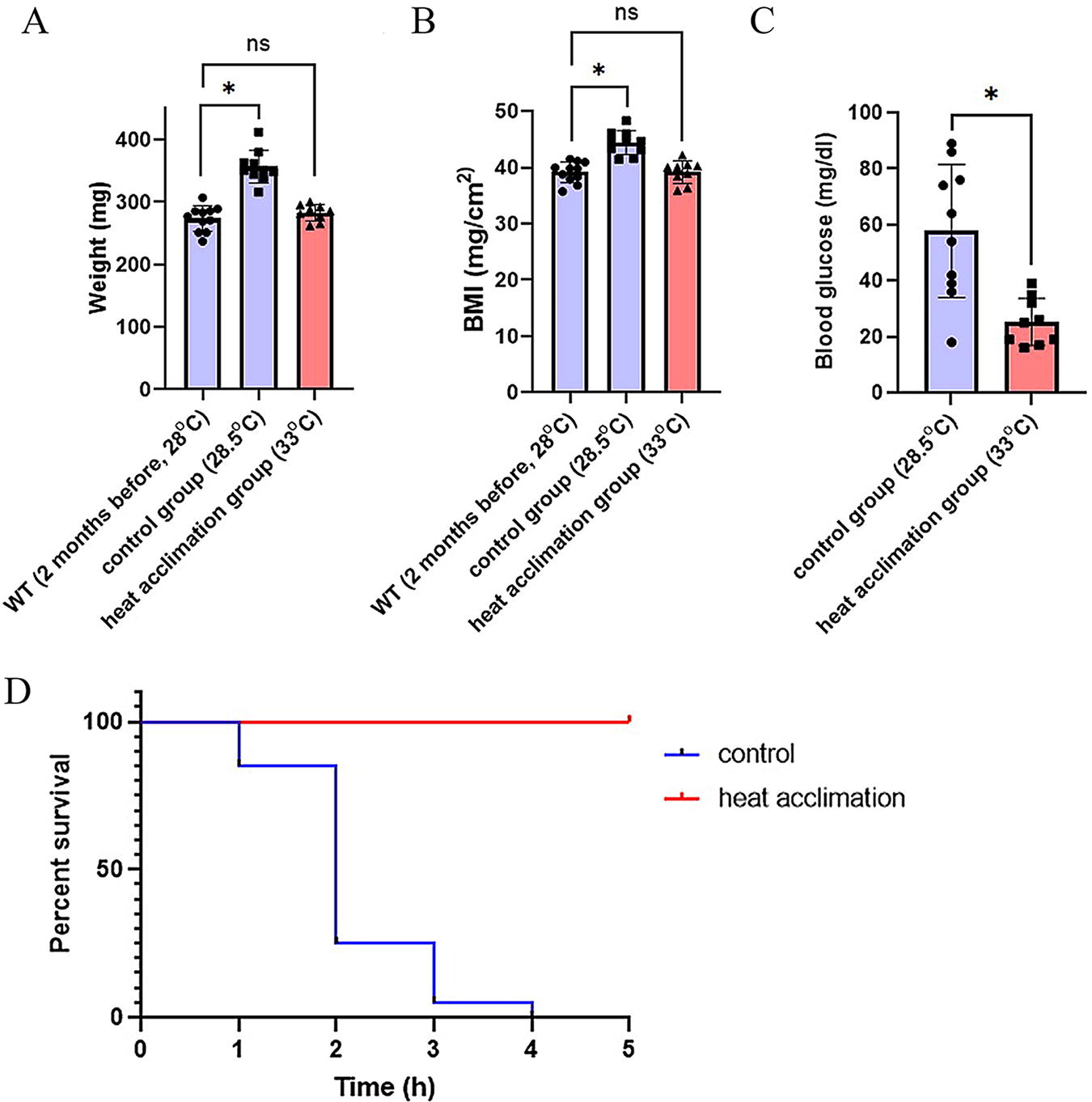
Heat acclimation causes physiological changes in zebrafish.
Heat acclimation-induced gene expression changes pertaining to mitochondrial function and mitochondrial fission in zebrafish liver
To explore the impact of heat acclimation on hepatic mitochondria, we assessed the mRNA expression profiles of genes associated with mitochondrial function. Specifically, we examined the expression levels of atp5f1b and ndufa9a, critical genes involved in mitochondrial oxidative phosphorylation (OXPHOS), as well as cpt1ab, a key player in mitochondrial fatty acid beta-oxidation. Strikingly, the heat-acclimated zebrafish exhibited a significant downregulation in the expression of these genes relative to the control group (Fig. 2A). To directly visualize morphological changes in liver mitochondria, we used a transgenic zebrafish strain expressing Green Fluorescent Protein (GFP) targeted exclusively to mitochondria, denoted as Tg(Xla.Eef1a1:mlsEGFP). Notably, heat acclimation resulted in the formation of smaller, more discrete hepatic mitochondria, indicative of the fission state (Fig. 2B).

Heat acclimation reduces the expression of functional mitochondrial genes and induces mitochondrial fission.
CQ-mediated inhibition of mitochondrial fission diminished heat tolerance in zebrafish
The alteration of gene expression associated with mitochondrial function and changes in mitochondrial morphology following heat acclimation suggest the potential involvement of mitochondrial dynamics in facilitating thermal tolerance. We postulated that mitochondrial fission might confer improved heat tolerance in zebrafish. Given the established capacity of glucose deprivation to induce mitochondrial fission, 36 we conducted an assessment of mitochondrial morphology in hepatic tissues from adult Tg(Xla.Eef1a1:mlsEGFP) zebrafish after a 2-day fasting period. The utilization of mitochondrial-targeted GFP revealed a notable fragmentation of mitochondria in fasted liver tissues, indicative of increased mitochondrial fission (Fig. 3A), mirroring the mitochondrial morphology observed in heat-acclimated zebrafish (Fig. 2B). CQ has been previously reported to promote the transition from fragmented to tubular mitochondria through the activation of adenosine 5′-monophosphate-activated protein kinase. 37 In our experimental approach, we administered 0.5 mM CQ and observed its effective inhibition of fasting-induced mitochondrial fission (Fig. 3A). Importantly, heat tolerance assessments demonstrated that fasting conferred enhanced heat resistance in zebrafish, and CQ treatment significantly attenuated the fasting-induced improvement in heat resistance (Fig. 3B), thereby corroborating the association between hepatic mitochondrial fission and heat resistance.

Fasting increases heat resistance and induces mitochondrial fission in adult zebrafish.
Short-term heat acclimation-induced mitochondrial fission enhances heat resistance in zebrafish embryos
A prior investigation had suggested that fish require a relatively long period to undergo heat acclimation. 38 To investigate whether hepatic mitochondria could be induced into a state of heat acclimation in a shorter time frame, we used zebrafish embryos ranging from 0 to 3 dpf and subjected them to incubation at 33°C until 4 dpf. Subsequently, we assessed their survival rates and hepatic mitochondrial morphology at 4 dpf. Zebrafish embryos at 0 dpf (2-cell stage) did not survive to 4 dpf when exposed to 33°C and were therefore excluded from this study, as depicted in Figure 4A. Conversely, embryos at 1 dpf or later developmental stages, when incubated at 33°C until 4 dpf, exhibited higher than 90% survival rate. Moreover, heat tolerance tests at 39°C demonstrated improved heat resistance in all groups of zebrafish embryos subjected to short-term heat acclimation (Fig. 4B). Notably, the 1–4 dpf and 2–4 dpf heat acclimation groups displayed increased survival rates compared with the 3–4 dpf group. In the 1–4 dpf and 2–4 dpf heat acclimation groups, the augmented survival was closely associated with increased mitochondrial fission (Fig. 4C), indicating a robust connection between fish survival and mitochondrial fission induced by heat acclimation.

Short-term heat acclimation induces mitochondrial fission and promotes heat resistance in zebrafish embryos.
To further substantiate the causative link between alterations in mitochondrial morphology and heat acclimation, we employed Mdivi-1, a specific inhibitor of Drp-1, to impede mitochondrial fission.35,39 Control embryos were maintained at 28.5°C, whereas embryos in the heat acclimation group were subjected to incubation at 33°C from 2 to 4 dpf, with or without Mdivi-1 treatment. Our observations revealed that, in comparison with the heat acclimation group, mitochondria in the heat acclimation + Mdivi-1 group exhibited a fused state (Fig. 4D). Furthermore, a heat tolerance test at 39°C disclosed lower survival rates in the heat acclimation + Mdivi-1 group compared with the heat acclimation group (Fig. 4E), signifying a diminished capacity for heat resistance by Mdivi-1 treatment. Collectively, hepatic mitochondrial fission via heat acclimation emerges as a pivotal factor in mediating heat resistance in zebrafish.
Discussion
The deleterious effects of heat stress, notably acute heat stroke, can result in substantial physiological impairment, including fatality, in fish. Therefore, elucidating the underlying mechanisms of heat resistance in fish is of paramount importance for the development of effective strategies to mitigate the impact of global warming on these organisms.
In this study, we used zebrafish as a model organism to investigate the potential mitigating effects of heat acclimation on the elevated mortality associated with heat stress. Our findings indicate that heat acclimation induces hepatic mitochondrial fission, leading to an enhancement of heat resistance in zebrafish. Furthermore, we observed that fasting-induced hepatic mitochondrial fission also confers increased heat resistance in zebrafish. Interestingly, intervention with a mitochondrial fission inhibitor attenuated heat resistance in both adult and larval zebrafish. These results suggest that promoting mitochondrial fission may constitute a viable approach to augment heat resistance in fish.
For the duration of heat acclimation in zebrafish, we assessed hepatic mitochondrial fission as the readout for successful heat acclimation and found at least 1-month period would be necessary to induce >90% hepatic mitochondria in their fission state (Supplementary Fig. S1). Our analysis using 2-month heat acclimation regimen would thus be enough to reliably establish heat-acclimatized zebrafish.
Although the exact mechanisms underlying the improved survival following mitochondrial fission under heat stress were not directly explored in this study, it is plausible to speculate that the observed elevation in autophagosome formation, a phenomenon associated with fission following acute heat stress, 25 may contribute to the heightened survival of zebrafish. Indeed, we found a significant reduction in the mitochondrial DNA contents in embryos undergone heat acclimation from 2 to 4 dpf, supporting dynamic reconstruction of mitochondrial contents during heat acclimation (Supplementary Fig. S2). Related to the observed changes in the mitochondrial morphology, we also found a significant changes in the gene expression directly associated with mitochondrial fusion and fission. In particular, we found increased expression of mitofusin 1b (mfn1b), opa1 mitochondrial dynamin like GTPase (opa1), fission mitochondrial 1 (fis1), and dynamin 1-like (dnm1l) in embryonic livers subjected to heat acclimation, whereas Mdivi-1 treatment reverted expression of these genes to the control levels (Supplementary Fig. S3), indicating dynamic changes in the mitochondrial morphologies during heat acclimation. However, since not only the expression of genes but posttranslational modifications of the proteins involved in mitochondrial dynamics are also critical, our analysis on the gene expression could provide only partial interpretation on mitochondrial dynamics during heat acclimation and heat stress.
Our findings suggest that fasting could serve as an effective and cost-efficient strategy for fish to overcome heat waves (Fig. 3). In addition, the use of compounds that facilitate mitochondrial fission may represent a promising intervention to rapidly enhance heat resistance, thereby advancing fish conservation efforts. It is worth noting that Mdivi-1, an inhibitor of mitochondrial fission, only partially suppressed the heat resistance induced by short-term heat acclimation (Fig. 4E). This partial effect could potentially be attributed to suboptimal Mdivi-1 concentrations in our experimental setup, as complete inhibition of mitochondrial fission may not have been achieved. However, we cannot exclude the possibility of additional mechanisms that may synergistically contribute to heat acclimation alongside mitochondrial fission. In this context, although our focus in this study was primarily on hepatic mitochondria, the mitochondrial morphology in other tissues may also exert an influence on heat resistance and therefore merits investigation in future studies. In our preliminary analysis on the embryonic heart tissue undergone heat acclimation, we did not observe noticeable change in the mitochondrial morphologies between control and heat-acclimated groups (Supplementary Fig. S4). Nonetheless, the potential involvement of metabolic tissues such as skeletal muscle and adipose tissue needs to be further analyzed.
In conclusion, we propose that induction of hepatic mitochondrial fission holds promise as a potential strategy to bolster heat resistance in zebrafish, offering insights into the prevention and treatment of heat stress. In the face of the unprecedented era of global warming and the increasing frequency of heat waves, this study provides novel mechanistic insights into heat resistance and adaptation, particularly concerning hepatic mitochondria. These findings have the potential to significantly contribute to the conservation and sustainable development of fish populations.
Footnotes
Acknowledgments
The authors thank members of Dr. Choe laboratory for providing constructive comments and sharing materials. This study was supported by Wonkwang University in 2023.
Authors’ Contributions
Y.M. performed the research, analyzed results, wrote and revised the article. L.L. performed the research and analyzed results. K.H. contributed to conceptualization, provided the reagents, and analyzed data. I.-K.N. provided the reagents. S.-H.K. wrote and revised the article. S.-K.C. contributed to conceptualization, experimental design, supervision, writing, and funding acquisition.
Disclosure Statement
The authors declare no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This article was supported by Wonkwang University in 2023.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.