
Abstract
Early identification of the potentially salvageable penumbra is critical for the determination of therapeutic intervention strategies in acute focal cerebral ischemia. This study differentiates the ischemic penumbra from the core on the basis of the dynamics of lactate formation. This was tested in a rat model of focal cerebral ischemia by infusion of [1-13C]-glucose, using lactate-edited magnetic resonance spectroscopic imaging techniques. The authors detected essentially no enrichment of lactate with 13C-label from the infused 13C-glucose in the ischemic core. However, in borderzone areas, 13C was incorporated into lactate, which could point toward compromised but potentially viable tissue. The authors' findings suggest that this combination of 13C-glucose infusion with the proposed magnetic resonance methods may aid in differentiating the penumbra from the core in cerebral ischemia.
In the acute stages of focal cerebral ischemia, the brain lesion encompasses an irreversibly injured core and a peripheral zone, where tissue is at risk but potentially viable (i.e., the penumbra) (see for review Hossmann, 1994). Without therapeutic intervention, the penumbra eventually will be recruited into the process of infarction. Therefore, early identification of this area is of crucial importance.
Recently, magnetic resonance imaging (MRI) and spectroscopy methods have demonstrated their sensitivity to pathophysiologic alterations in brain ischemia as early as in the hyperacute stages (see for review Fisher et al., 1995). Diffusion-weighted MRI allows the detection of the ischemia-induced acute reduction of the apparent diffusion coefficient (ADC) of tissue water, which can be associated with the development of cytotoxic edema (Moseley et al., 1990). 1H-NMR spectroscopic imaging (MRSI) provides a means to spatially assess the metabolic status of ischemic tissue from the detection of metabolites such as the neuronal marker N-acetylaspartate and lactate, the end-product of anaerobic glycolysis (Higuchi et al., 1993). The immediate uncoupling of glycolysis from oxidative phosphorylation during ischemia provides a basis for early differentiation between the core and the penumbra. In the ischemic core, where perfusion has dropped dramatically, the lactate pool is expected to essentially fully originate from glucose that was present at the time of onset of ischemia. In areas with residual perfusion, ongoing lactate formation and lactate washout can be expected to occur, suggesting a dynamic lactate pool. In a previous study using localized 1H-MRS, 13C-enriched lactate was detected in the brain of a chronic stroke patient after [1-13C]-glucose infusion (Rothman et al., 1991).
This study evaluates the potential of MRSI of lactate to distinguish the penumbra from the ischemic core in acute focal ischemic brain tissue. We specifically tested the implicit hypothesis that regions in which the ADC of water continues to decline sustain a significant increase of lactate. To that end, we combined diffusion-weighted MRI with measurements of the distribution and time course of the incorporation of 13C-label into lactate during infusion with [1-13C]-glucose, by use of 1H-MRSI, tailored for the exclusive detection of lactate.
METHODS
Male Fisher rats (225 to 325 g) (n = 8) were anesthetized by a subcutaneous injection of a mixture of 0.55 mL/kg fentanyl citrate (0.315 mg/mL) and fluanisone (10 mg/mL), and 0.55 mL/kg midazolam. After endotracheal intubation, animals were mechanically ventilated with O2/N2O (1:2). Body temperature was maintained at 37.5 ± 0.5°C by means of a feedback-controlled heating pad. The left femoral artery was cannulated for continuous recording of blood pressure and blood sampling. Plasma glucose levels and blood gas tensions were frequently measured. The left femoral vein was cannulated for infusion with D-[1-13C]-glucose (Omicron, Biochemicals, Inc., South Bend, IN, U.S.A.) (6.7 g/kg in 5 mL of saline). Permanent focal cerebral ischemia was induced by unilateral tandem occlusion of the common carotid artery and the middle cerebral artery, as described by Brint and others (1988). Infusion with [1-13C]-glucose (Patlak and Pettigrew, 1976) was started immediately after middle cerebral artery occlusion (MCAO), after which the rats were positioned in the magnet.
The MRI measurements were performed in a 4.7-T SISCO/Varian NMR spectrometer (Palo Alto, CA, U.S.A.) using a 1H/13C surface coil (1H: circular [1.2-cm diameter]; 13C: butterfly [1 × 2 cm loop dimensions]). During the MRI protocol, anesthesia was maintained by ventilation with 2% isoflurane. Diffusion-weighted MRI (spin-echo sequence; repetition time 2 seconds; echo time 100 milliseconds; one [b = 1325 s/mm2, n = 4] or three b values [b = 150 to 1600 s/mm2, n = 4]) (diffusion-sensitization parallel to the long axis of the brain); two acquisitions; field-of-view 3 × 3 cm2; 64 × 64 data matrix) of a 1.5-mm coronal slice was done to identify the ischemic area (approximately 0.5 and 3 hours after MCAO). Where multiple b values were used, calculation of quantitative brain maps of the water ADC was done by monoexponential fitting. The distribution of [3-12C]- and [3-13C]-lactate was measured with a homonuclear lactate-edited two-dimensional 1H-MRSI (adiabatic gradient-enhanced multiple quantum-coherence filtering sequence [De Graaf et al., 1995]; repetition time 1.5 seconds; echo time (1/J) 144 ms; four acquisitions; 16 × 16 data matrix; field-of-view 18 × 18 or 24 × 24 mm2; 1.5- or 2.0-mm slice [nominal voxel sizes 1.9 to 4.5 μL]). Homonuclear lactate-edited MRSI was repeated every 30 minutes from approximately 0.5 to 3 hours after MCAO. In two cases, we verified the production and distribution of [3-13C]-lactate with heteronuclear 1H-observed/13C-edited MRSI (adiabatic gradient-enhanced heteronuclear multiple quantum-coherence filtering sequence [De Graaf et al., 1995]; repetition time 2.0 seconds; echo time (1/J) 8 ms; eight acquisitions; 16 × 16 data matrix; field-of-view 24 × 24 mm2; 2.0-mm slice) with decoupling during acquisition at 4 hours after MCAO.
Data are presented as mean ± SD. Statistical analysis was done by one-way analysis of variance with repeated measures. Groups were compared by use of a paired Student's t-test, with Bonferroni correction when appropriate. P < 0.05 was considered significant.
RESULTS AND DISCUSSION
Intravenous infusion of [1-13C]-glucose resulted in a significant increase of plasma glucose from 6.6 ± 0.5 mmol/L to 34.5 ± 7.2 mmol/L (P < 0.001) within 10 minutes. Plasma glucose remained significantly elevated during the following 3 hours (19.8 ± 3.0 mmol/L after 3 hours, P < 0.001).
Middle cerebral artery occlusion induced a focal cerebral ischemic lesion, which was delineated as a clear ipsilateral hyperintense area on diffusion-weighted images (Fig. 1A). Calculated maps of the tissue water ADC revealed normal ADC values (0.7 to 0.8 × 10−3 mm2/s) in contralateral cortices. The ADC, as measured from volumes-of-interest (4.5 μL) positioned in the periphery of the lesion (i.e., the primary motor cortex) and the ischemic core (i.e., the primary somatosensory cortex), revealed a significant decline from 0.47 ± 0.05 to 0.38 ± 0.04 × 10−3 mm2/s (P < 0.05) in the borderzone between 0.5 and 3 hours after MCAO, whereas in the core the ADC already was dramatically reduced after 0.5 hours (0.37 ± 0.06 × 10−3 mm2/s) and showed no further change after 3 hours (0.38 ± 0.04 × 10−3 mm2/s). Since a reduction of the tissue water ADC is typically associated with the formation of cytotoxic edema (Moseley et al., 1990), our results suggest that the development of cytotoxic edema proceeded in the peripheral areas, while already maximal in the core.
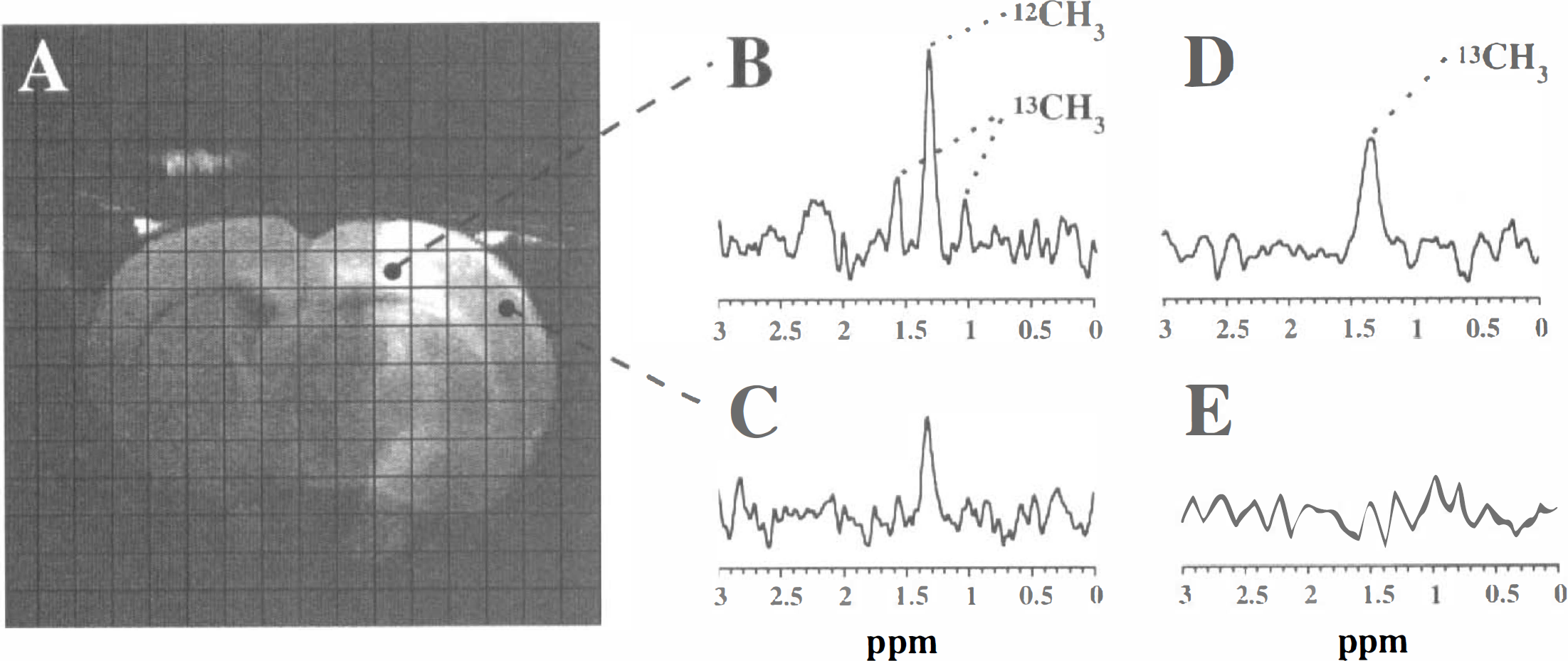
Homonuclear lactate-edited MRSI revealed exclusively lactate peaks, which were detected only in voxels within the ischemic region. Figure 1B and C show spectra from voxels in the ischemic borderzone and core, respectively. Typically, at the boundaries of the lesion, the center signal from [3-12C]-lactate methyl protons at 1.33 ppm was clearly flanked by [3-13C]-lactate methyl proton signals, which were split into a doublet by heteronuclear spin-spin coupling (J ∼ 125 Hz). In the deeper central areas, 13C-lactate satellite peaks were rarely detected. Fig. 2A illustrates that fractional enrichment of lactate with 13C-label was the highest at the boundaries of the lesion. Thus, although tissue was compromised in the borderzones, these regions postischemically received glucose (i.e., residual perfusion), which was metabolized into lactate.

The presence or absence of 13C-lactate satellite peaks, as observed with homonuclear lactate-edited MRSI, was invariably confirmed with heteronuclear 1H-observed/13C-edited MRSI from the presence or absence of signal from the 13C-lactate methyl protons (Fig. 1D and E).
In the borderzone, the lactate concentration increased dynamically and was significantly elevated after 2 to 3 hours compared with the situation after 0.5 to 1 hours (Fig. 2B). In the core, the modest lactate concentration increase was not significant (Fig. 2B). The 13C-lactate fraction in the borderzone already was 30% to 40% after 0.5 to 1 hours and remained at this level up to 3 hours (Fig. 2B). Since theoretically the maximal fractional enrichment is 50% (see Rothman et al., 1991), our results suggest that the lactate present in these areas was almost completely derived from blood glucose during infusion.
In this study, the start of [1-13C]-glucose infusion coincided with the onset of ischemia. By initiating the [1-13C]-glucose infusions at varying time points and by alternating infusions with [1-13C]-glucose and [1-12C]-glucose in pulse-chase-type experiments, we expect to gain new insights in the sequence of metabolic changes in both permanent and transient focal cerebral ischemia. Importantly, since hyperglycemia may seriously affect outcome in cerebral ischemia, future studies also should assess the effect of elevated glucose levels on both metabolic impairment and tissue injury.
In conclusion, this study has shown that homonuclear- and heteronuclear-edited MRSI combined with infusion of [1-13C]-glucose may provide a means to distinguish irreversibly damaged and potentially viable ischemic tissue at a metabolic level. These techniques, in combination with other MRI modalities such as perfusion- and diffusion-weighted MRI, thereby would provide a powerful tool to differentiate the penumbra from the core in cerebral ischemia.
Footnotes
Acknowledgments
The authors thank Gerard van Vliet and Yanping Luo for expert technical assistance.