
Abstract
Objective
The overarching objective was to evaluate whether workload sensory-domain specificity could be identified through electroencephalogram (EEG) recordings during simulated rotary-wing operations.
Background
Rotary-wing aviators experience workload from different sensory domains, although predominantly through auditory and visual domains. Development of real-time monitoring tools using psychophysiological indices, such as EEG recordings, could enable identification of aviator overload in real time.
Method
Two studies were completed, both of which recorded EEG, task performance, and self-report data. In Study 1, 16 individuals completed a basic auditory and a basic visual laboratory task where workload was manipulated. In Study 2, 23 Army aviators completed simulated aviation flights where workload was manipulated within auditory and visual sensory domains.
Results
Results from Study 1 found differences in frontal alpha activity during the auditory task, and that alpha and beta activities were associated with perceived workload. Frontal theta activity was found to differ during the visual task while frontal alpha was associated with perceived workload. Study 2 found support for frontal beta activity and the ratio of beta to alpha + theta to differentiate level of workload within the auditory domain.
Conclusion
There is likely a role of frontal alpha and beta activities in response to workload manipulations within the auditory domain; however, this role becomes more equivocal when examined in a multifaceted flight scenario.
Application
Results from this study provide a basis for understanding changes in EEG activity when workload is manipulated in sensory domains that can be used in furthering the development of real-time monitoring tools.
Keywords
Background
With increasing application of automated systems, accurately assessing cognitive workload is essential for understanding human–machine interactions (HMI; Tsang & Vidulich, 2006) and designing systems compatible with operator capabilities. Multiple resource theory (MRT; Wickens, 1980, 1984, 2002, 2008), a framework for understanding workload and associated human performance outcomes, postulates that cognitive resources are parsed into separate dimensions. Broadly, these dimensions can be considered as sensory domains: cognitive, motor, visual, and auditory. Ultimately, performance outcomes requiring multitasking are dictated by the degree to which separate tasks utilize resources from the same sensory domain.
Wickens (2008) asserted MRT has neurophysiological plausibility, whereby specialized cortical networks of the brain correspond to different resource domains. The cognitive neuroarchitecture 4CAPS (Cortical Capacity-Constrained Concurrent Activation-based Production System; Just & Varma, 2007) and associated functional neuroimaging studies (e.g., Just et al., 1996, 2001) have shown different cortical networks are activated with changes in qualitatively different task demands (i.e., verbal vs. spatial). The connection between MRT and separate cortical networks has implications for assessing cognitive workload during operational conditions and driving adaptive automation (AA) systems through real-time physiological monitoring, thereby supporting HMI. Specifically, workload assessment may be improved with physiological measures that correspond to qualitatively different sources of resource demands. For example, certain electroencephalographic bandwidths (i.e., alpha, beta, theta) may be more sensitive to demand fluctuations within a specific sensory domain. This could enable domain specificity within neurophysiological metrics, and translate to AA systems driven by physiological measures or, at minimum, use of neurophysiological metrics in assessing workload during the design process. The present work addresses whether such a process is possible by examining the degrees to which physiological measures covary with performance and perceived domain-specific workload.
Electroencephalography
Electroencephalogram (EEG) has frequently been used to quantify workload (e.g., Hockey et al., 2009; Hogervorst et al., 2014) with support for practical use of EEG metrics in improving HMI through workload detection (Aricò, Borghini, Di Flumeri, Colosimo, Pozzi, et al., 2016) and in driving AA (Aricò, Borghini, Di Flumeri, Colosimo, Bonelli, et al., 2016). A defining feature of workload is activation in frontal brain regions (e.g., Aricό et al., 2017; Gevins & Smith, 2007; So et al., 2017), although certain features in the parietal region are also indicative of workload variations (e.g., Berka et al., 2007; Smith et al., 2001). Selecting one region to measure (i.e., frontal) minimizes required electrode sites, thus increasing usability in applied contexts (So et al., 2017). Theta (4–8 Hz), alpha (9–13 Hz), and beta (14–30 Hz) frequency bands are commonly examined in workload manipulations, where patterns have included increased theta and decreased alpha power spectral density (PSD) with increased task demands (Borghini et al., 2014). Increased power within beta corresponds with overload (Borghini et al., 2014; Matthews et al., 2017).
Evaluating changes in power within frequency bands enables examination of relationships between behavior and neural oscillations, rhythmic fluctuations in excitability of neural populations believed to play a “gate and filter” role for incoming stimuli (Schroeder & Lakatos, 2009; Zoefel & Van Rullen, 2017). Filtering occurs as fluctuations within different bands reflect either inhibitory or excitatory processes, namely activating or suppressing activity within a brain region. Fluctuations within bands influence the responsiveness of a brain region through moderation of neural firing rates (Haegens et al., 2011; Jensen et al., 2012), which impacts the processing of information relative to that specific region. For example, increased theta power and suppression of alpha power in frontal regions is suggestive of cognitive overload and resource depletion (Borghini et al., 2014). Although there are well-documented patterns of physiological responses that accompany changes in task demands (e.g., Borghini et al., 2014), these patterns are typically explored with global changes in task parameters, not separate sensory domains (i.e., visual vs. auditory). Physiological measures of workload may depend on the qualitative nature of the resource demands placed on the operator (Hankins & Wilson, 1998; Ryu & Myung, 2005).
EEG in Differentiating Sources of Workload
Neural oscillations play a significant role in attentional selection and stimulus processing across sensory domains (e.g., Arnal, 2012; Galán & Beal, 2012; Lakatos et al., 2008; Stefanics et al., 2010). Moreover, recent work shows differences between sensory domains in terms of the oscillatory mechanisms of stimulus selection (Thorne & Debener, 2014), particularly prevalent between the auditory and visual domains within the frontal region (Zoefel & Van Rullen, 2017). Given that the frontal region has been associated with response to workload manipulations, closer examination of frequency oscillations may give insight to domains being overtaxed.
Rotary-wing aviators are exposed to tasks of differing sensory domains when flying, predominantly, the visual domain given the large amounts of visual information to process (Webb et al., 2010). Additionally, Army rotary-wing aviators rely on auditory sources of information to complete missions, such as the integration of radio calls from onboard crew, ground stations, and other aircraft.
Differentiating domain-specific sources of resource utilization within rotary-wing aviation is critical for the advancement of real-time operator state monitoring and neuroergonomic evaluations of new systems. One solution to minimize workload within the auditory domain is spatial audio, which entails augmenting sounds with virtually mapped spatial locations around the operator. Spatially mapped radio transmissions have significantly improved target radio call detection accuracy and increased cerebral blood flow (CBF) velocity, measured by transcranial Doppler sonography, compared to monaurally delivered radio transmissions (Shaw et al., 2013). With the development of domain-specific cueing systems, such as spatial audio, more refined physiological measures of workload could improve diagnosing sources of increased workload. However, relying on physiological measures to differentiate high and low workload, without differentiating the source of workload when evaluating these metrics, assumes that the metrics are effective regardless of workload sources. This is in opposition to what Wickens (2008) postulated with the neurophysiological underpinnings of MRT.
Two studies were completed to assess whether measuring EEG can identify workload relating to the auditory and visual sensory domains. Study 1 examined EEG indices relative to workload level during an auditory discrimination task and a visual search task. Study 2 evaluated whether similar EEG patterns occurred while completing flight tasks designed to tax each domain using a Black-Hawk simulator. The results reported here are a subset of a larger study (Feltman et al., 2019) and focus only on frontal scalp locations.
Study 1
Methods
Participants
Sixteen participants (three women; M age = 31.33 years, SD = 9.72) were recruited from the Fort Rucker, AL, area. Exclusion criteria included disorders affecting cognitive function (depression, anxiety) and hearing and visual impairment. Participants were required to sleep at least 6 hr, refrain from taking over-the-counter medications which may induce drowsiness, refrain from any alcohol or sedatives 24 hr, and refrain from nicotine use 2 hr and caffeine 16 hr prior to the study (confirmed by self-report). These criteria were enforced to reduce variability in physiological recordings (e.g., Gilbert et al., 2000; Kähkönen et al., 2003).
Materials
Subjective workload
The NASA Task Load Index (TLX; Hart & Staveland, 1988) was used to measure subjective workload. Using a 100-point scale, participants rate mental demand, physical demand, temporal demand, performance, effort, and frustration subscales and then provide assessments of which factors contributed to workload through pairwise comparisons. The ratings and weights were used to calculate weighted total scores.
Electroencephalography
EEG data were recorded using the Advanced Brain Monitoring (ABM) B-Alert X-24® wireless wet electrode system (256 Hz sampling rate) with 20 electrodes placed according to the International 10–20 system. Frontal sites examined included: Fp1, Fp2, F7, F3, Fz, F4, and F8. The EEG signal was referenced to linked mastoid electrodes. A 0.5–65 Hz bandpass filter was applied to the signal. Notch filters (50, 60, 100, 120 Hz) were used to sharply attenuate environmental noise not reduced by the bandpass filter. PSD values were computed using the automated algorithms provided through the B-Alert Live® data acquisition software (ABM: Advanced Brain Monitoring, Inc, 2009). Prior to computing PSD values, artifacts (spikes, excursions, amplifier saturations, electromyography, and eye blinks) were identified and removed using algorithms developed by ABM (ABM: Advanced Brain Monitoring, Inc, 2009; Berka et al., 2004). The software fast Fourier transformed the raw EEG signal data and calculated the amplitudes of the sinusoidal components for designated frequency bins. Frequency domain variables were based on the PSD derived after application of a 50% overlapping window and Kaiser window. Each window size was defined as one epoch containing 1 s of data (256 decontaminated samples). The software then provided PSD values ranging from 1 to 40 Hz for each EEG channel that were logged to obtain a Gaussian distribution. Selected relative 1 Hz bins were averaged to create the EEG bands used in analyses (theta: 4–8 Hz; alpha: 9–13 Hz; beta: 14–30 Hz). PSD values were examined from frontal channels with values averaged across each task. An engagement index of the ratio of beta values to alpha + theta values (Freeman et al., 2000) was also examined.
Auditory discrimination task
Participants listened to two different pure sine-wave tones (one low frequency, 1,000 Hz; one high frequency, 2,000 Hz) presented binaurally through headphones. The low-frequency tone occurred on 80% of trials, while the high-frequency tone occurred on 20% of trials and served as the target in all workload conditions. Each tone was presented for 100 ms with a 10 ms rise and fall time. Inter-stimulus intervals randomly varied between 800 and 1,200 ms. Participants responded to targets in each condition by pressing the spacebar. Three workload conditions were completed, each presenting 200 sequential stimuli. Condition 1 was a control condition, a simple odd ball detection task (Kramer & Weber, 2000; Trejo et al., 1995) which required participants to respond as quickly as possible when a target was detected (infrequent 2,000 Hz tones) and inhibiting responses on nontarget trials (frequent 1,000 Hz tones). Condition 2 was a low workload condition, a counting forward task where participants responded when a target was heard, simultaneously counting the number of target occurrences and verbally report this total at the end of the task. Finally, Condition 3 was a high workload condition, a counting backward task which required participants to respond to the target and then subtract 1 day from the current date for each target detected (i.e., counting in reverse order) and verbally report the date calculated at the end of the session. At the start of the task, participants adjusted the volume to a comfortable level, ranging from 80 to 90 dB. A 3-min break was given between each condition. Reaction times (RT) and accuracies were recorded. Completion of all three conditions required approximately 30 min (10 min per condition). These workload manipulations have been previously validated (Horat et al., 2016).
Visual search task
For the visual domain, participants performed a visual search task at low- and high-workload levels. During the low-workload condition, each trial consisted of four letters presented in a vertical column (display duration = 2,000 ms) and participants determined whether the target letter (“X”) was present. If the letter X was present, participants pressed the spacebar and made no response if the letter X was not present. Distractor letters included E, T, S, D, U, W, Q, Z, I, O, and B. Between trials, a blank screen was presented for 150 ms. During the high-workload condition, four columns with four letters each were presented. Participants identified if the letter X was present by pressing a key that was spatially mapped to the column in which the X was displayed. Accuracy and RT were recorded. Task parameters were previously validated (Cassenti & Kelley, 2006, Cassenti & Kelley, 2008; Colle, 2010). Total completion time was 30 min.
Procedure
The U.S. Army Medical Research and Materiel Command Office of Research Protections Institutional Review Board reviewed and approved the protocol for both studies. All procedures were conducted according to institutional ethical standards. Participants provided written informed consent before participation. Participants were fitted with the EEG leads and then provided instruction on each task, including practice trials. Participants then completed three iterations of the auditory task (control, low, and high workloads) and two of the visual task (low and high workloads). The order of each task block (auditory, visual) was counterbalanced, as well as the workload of the visual task, and workload randomized for the auditory task. Following each task iteration, participants completed the TLX (five times total). All procedures were completed in a single, independent session.
Statistical Analysis and Quality Control
Distributions of performance and TLX weighted total scores were evaluated for normality; outliers exceeding three standard deviations from the mean were excluded from analyses. To determine the efficacy of the workload manipulations on performance (accuracy and RTs), perceived workload (TLX scores), and EEG activity, repeated-measures analyses of variance (ANOVAs) and paired samples t-tests were used. Analyses for the auditory and visual tasks were conducted independently. Finally, correlational analyses were used to evaluate the concurrent validity of the EEG as indices of workload (relative to the TLX scores). Statistical significance was set at α = .05 and η p 2 is reported as the effect size for omnibus ANOVAs.
Results
Performance
There was no effect of the workload manipulation on the number of correct responses during the auditory task, F(2, 30) = 0.21, p = .81, but responses were slightly slower in the control condition than the low-workload condition, F(2, 30) = 3.62, p = .051, ηp 2 = 0.31 (Table 1). For the visual search task, there was a significant effect of the workload manipulation on the total number of correct responses, F(1, 13) = 13.30, p = .003, ηp 2 = 0.51, and on RTs, F(1, 13) = 95.88, p = .001, ηp 2 = 0.88. The total number of correct responses was lower and RTs slower in the high-workload condition compared to the low-workload condition (Table 2).
Auditory Task: Descriptive Statistics Including Means and Standard Errors of the Mean (SE)
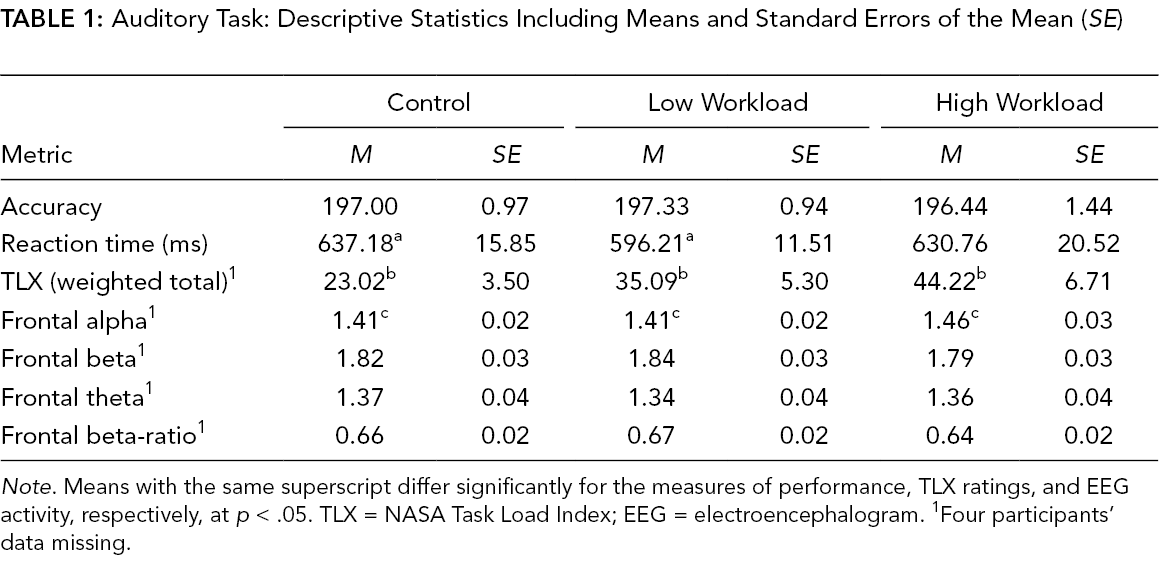
Note. Means with the same superscript differ significantly for the measures of performance, TLX ratings, and EEG activity, respectively, at p < .05. TLX = NASA Task Load Index; EEG = electroencephalogram. 1Four participants’ data missing.
Visual Task: Descriptive Statistics Including Means and Standard Errors of the Mean (SE)

Note. Means with an asterisk differ significantly at p < .05. TLX = NASA Task Load Index. aTwo participants’ data missing.
Workload
There was a significant effect of workload on the TLX scores for both tasks: auditory, F(2, 26) = 6.44, p = .001, ηp 2 = 0.31 (Table 1); visual, F(1, 13) = 22.27, p < .001, ηp 2 = 0.61 (Table 2). As seen in Table 1, follow-up comparisons revealed significant differences between the TLX ratings for all three auditory task workload conditions.
EEG
There was an effect of workload on frontal alpha activity during the auditory task, such that activity increased with more difficult workload levels, F(2, 22) = 5.67, p = .010, ηp 2 = 0.34. No significant effects of workload emerged with frontal beta, F(2, 22) = 2.68, p = .126, or theta activity, F(2, 22) = 4.63, p = .09 (Table 1). There was an effect on frontal theta activity during the visual task, where activity decreased with higher workload, t(13) = 2.43, p = .030, d = .54 (Table 2). There were no effects of alpha or beta during the visual task.
Correlation coefficients between TLX scores and EEG across all conditions were calculated. Perceived workload on the auditory task correlated with changes in frontal alpha activity, r(11) = .65, p = .030, and beta activity, r(11) = −.64, p = .03, such that alpha activity increased whereas beta activity decreased with workload. On the visual task, frontal alpha activity and perceived workload were significantly correlated, r(14) = .41, p = .024, such that activity increased as perceived workload increased.
Discussion
These initial results suggest a role of frontal alpha and beta activity during an auditory discrimination task, and that the degree of perceived workload was related to differences in spectral power within these bands. During the visual task, changes to frontal theta activity due to workload were found, while alpha was associated with perceived workload. We then investigated these patterns in an ecologically valid environment using flight simulation.
Study 2
Methods
Participants
Twenty-three male U.S. Army rated, rotary-wing pilots were recruited from the Fort Rucker, AL, area (M age = 36.13 years, SD = 4.99). Reported average flight time in the last year ranged from 12.50 to 480 hr (M = 198.37, SD = 115.40). Exclusion criteria were consistent with Study 1.
Materials
The NASA-TLX and EEG were administered and recorded, respectively, using the same methodology as Study 1.
Simulated flight
Flight performance was evaluated in a full-motion Black-Hawk flight simulator composed of a compartment containing a cockpit, instructor/operator station, observer station, and a six degree-of-freedom motion system. Aircraft/simulator state parameters were collected at a 60 Hz capture rate. Four flight scenarios, designed by in-house research pilots, were validated for workload manipulations using three pilots naïve to the study and scenarios. Two scenarios (low and high workloads) emphasized the auditory domain and two (low and high workloads) emphasized the visual domain.
During the auditory domain scenarios, workload was manipulated with a radio calls task that required participants to fly from one airport to another (approximately 10 min apart) and respond to several air traffic control calls en route. In the low-workload condition, 100% of calls were ownship (eight calls total). In the high-workload condition, 40% of calls were ownship, while the remaining were distractors (34 total). Participants were given a specific call sign prior to the flight and were instructed that radio calls would not be repeated. Day view conditions were employed (Figure 1). The proportion of correct radio calls was used as an outcome measure.

Day view of cockpit within the Black-Hawk Simulator.
During the visual domain scenarios, visibility was used to manipulate visual workload while participants conducted a terrain flight from one airport to another. Participants followed the contours of a valley while maintaining an altitude at or below 200 ft above ground level. The low-workload condition required participants to fly during the daytime (Figure 1) and the high-workload condition required participants to fly during nighttime (Figure 2).

Night view of cockpit within the Black-Hawk Simulator.
For all flights, winds were calm and participants were given target altitude and airspeeds to maintain while actively flying the scenarios (i.e., controlling flight surfaces and navigating). Routes were preprogrammed into the flight global positional system. Deviations from these target parameters were used as outcome measures. Specified target altitude and airspeed parameters did not change once instructed by the research pilot.
Procedure
First, participants were fitted with the EEG and then escorted to the simulator where they completed familiarization flights. A research pilot verified proficiency with required flight maneuvers. Participants then completed high- and low-workload conditions of the two flight scenarios (approx. 12 min each; four flights total). Order of flight scenarios were randomized by sensory domain (auditory, visual) and within each domain, counterbalanced by workload level (Table 3). The TLX was completed at the end of each flight.
Flight Scenario Descriptions

Note. Day view was used for both auditory domain flights.
Statistical Analysis and Quality Control
All procedures were identical to those for Study 1 with the exception of workload manipulation being assessed only with paired-samples t-tests.
Results
Performance
Flight performance varied by workload conditions in both sets of domain scenarios. Specifically, airspeed deviations during the auditory flight were greater during the high-workload compared to low-workload condition, t(19) = 5.68, p < .001, d = 1.27 (Table 4). Similarly, performance on the radio calls task was worse in the high- compared to low-workload condition, t(22) = −3.04, p = .006, d = .63 (Table 4). Significant differences in both altitude and airspeed deviations were found for the visual flight. Altitude deviations, t(19) = 3.46, p = .003, d = .77, and airspeed deviations, t(19) = 2.16, p = .044, d = .48, were greater during the high-workload compared to the low-workload condition (Table 5).
Auditory Flight: Descriptive Statistics Including Means and Standard Deviation of the Mean (SD)
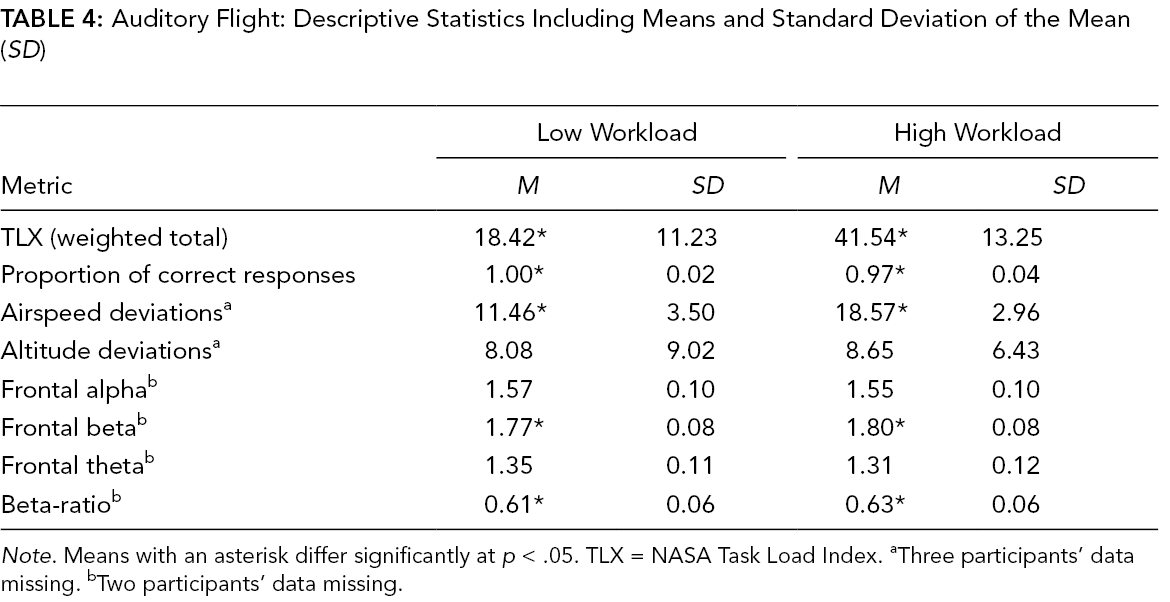
Note. Means with an asterisk differ significantly at p < .05. TLX = NASA Task Load Index. aThree participants’ data missing. bTwo participants’ data missing.
Visual Flight: Descriptive Statistics Including Means and Standard Deviation of the Mean (SD)
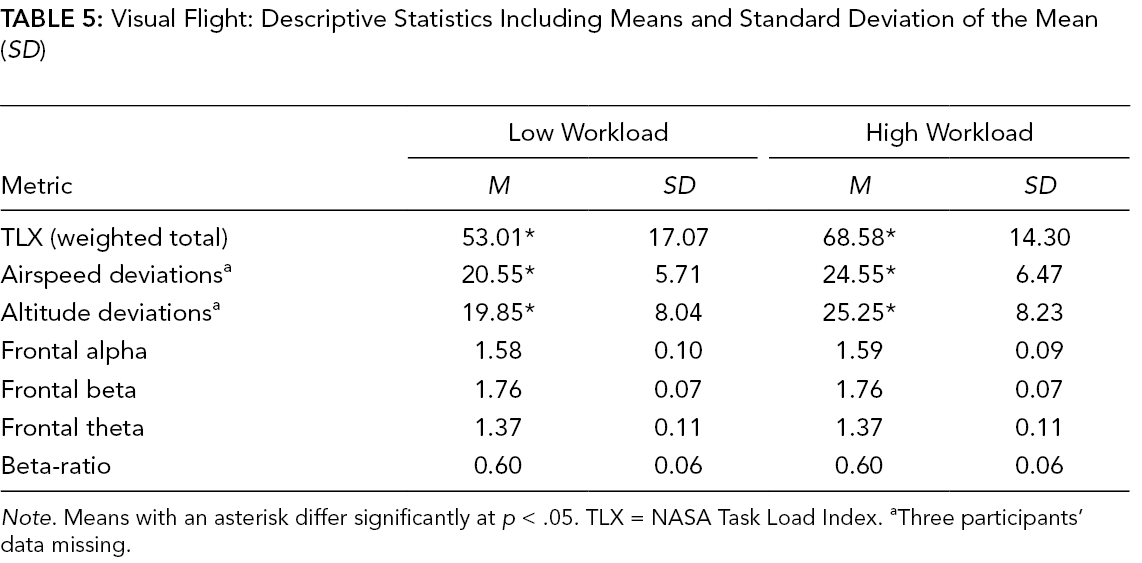
Note. Means with an asterisk differ significantly at p < .05. TLX = NASA Task Load Index. aThree participants’ data missing.
Workload
TLX scores significantly differed between workload conditions. Workload was rated significantly higher in the high-workload condition compared to the low-workload condition for both the auditory domain during the auditory flight, t(22) = −7.49, p < .001, d = 1.56 (Table 4), and the visual domain during the visual flight, t(22) = −5.93, p < .001, d = 1.24 (Table 5).
EEG
EEG outcome metrics varied between high- and low-workload conditions for the auditory flights. Both frontal beta values, t(20) = −2.55, p = .018, d = .56, and frontal beta-ratio values, t(20) = −2.58, p = .02, d = .56, were sensitive to workload. Each was significantly higher during the high-workload compared to low-workload condition (Table 4). No significant differences were found for theta or alpha values. There were no differences in EEG metrics for the visual flights.
There were no significant correlations found between performance measures and TLX ratings nor EEG metrics and TLX ratings. Significant correlations were found for the visual flight high-workload condition between airspeed deviations and overall TLX ratings, r(22) = .50, p = .02, such that airspeed deviations increased with higher workload ratings.
Because of the wide range of flight experience, pilots with less experience may have confounded results. Therefore, analyses were repeated excluding participants with less than 100 hr (n = 4) of flight time within the past year. Results of the hypothesis tests were the same as reported above for flight performance and TLX measures (i.e., statistically significant results remained significant and nonstatistically significant results remained nonsignificant). In contrast, results of EEG analyses were no longer statistically significant (p-values > .05). However, effect sizes were small to medium (d = 0.33–0.43), indicating that the hypothesis tests were likely underpowered. Indeed, other studies (e.g., Matthews et al., 2015) reported similar effect sizes with a much larger sample size.
Discussion
The results suggest frontal beta values and the beta-ratio differentiate workload within the auditory domain. There was no support for frontal EEG metrics to differentiate workload within the visual domain.
General Discussion
The goal of these studies was to determine whether different EEG oscillation patterns occurred based on sensory domain of workload, which could be used to aid in the design of AA systems. For each study, analyses focused on frontal EEG features which are the most conducive for applied settings. In Study 1, support was found for frontal activity differentiating workload: as workload became more difficult, alpha increased (auditory task) and theta decreased (visual task). However, different frequency bands were sensitive to workload in Study 2 and only during the auditory task, with higher beta and beta-ratio values in the high-workload condition.
The differing results between Study 1 and Study 2 may be due to the differences in task complexity. The tasks in Study 1 strictly relied on auditory and visual stimuli, while the simulated flights incorporated several sources of workload (e.g., maintaining flight parameters based on the simulation’s displays), in addition to the experimental manipulations of the auditory calls and day/night visual condition. The complexity of the flights likely resulted in different EEG activation patterns than the Study 1 tasks. Several functional neuroimaging studies (e.g., Just & Varma, 2007; Mandrick et al., 2016; Mirelman et al., 2014) have demonstrated that dual-tasking results in more distributed cortical activation, potentially explaining why different EEG patterns were observed. Interestingly, no differences were found during the visual flights in EEG metrics, despite confirmation of workload manipulation with performance metrics and subjective ratings. This suggests that one measure of workload (i.e., EEG) may not be enough to reliably differentiate workload. Our findings are similar to Matthews et al. (2015) who reported divergent results when examining multiple psychophysiological indices of cognitive workload. Specifically, they concluded that multiple neurocognitive responses likely occur when responding to workload, which could produce differences in responses across individuals. Further studies are needed to refine the diagnosticity of workload using psychophysiological indices, including different recording sites.
There remains plausibility in using EEG to identify differences in sensory domain taxed. For example, changes in alpha activity during the first study within the auditory task are consistent with past literature postulating that alpha oscillations may play a role in auditory inhibition and stimulus processing (Strauß et al., 2014). Specifically, increased alpha activity in the frontal region is thought to aid in processing auditory information by reducing the interference of nonrelevant information. Schrauf et al. (2011) demonstrated that alpha band activity significantly increased when an auditory distractor task occurred during a driving task. However, changes in alpha activity during Study 2 when auditory workload was manipulated through radio calls were not significant. Rather, the results of Study 2 are more in line with research regarding increased beta values within operational environments, including flight (Di Stasi et al., 2015; Dussault et al., 2004) and shipping operations (Orlandi & Brooks, 2018). In contrast to the findings here, a review by Borghini et al. (2014) found that flight and driving demands were associated with consistent changes in the theta band. It may be the case that the theta band reflects global increases in task demands associated with the integration of several sources of information (Klimesch, 1999; Sauseng et al., 2005, 2010). Given that the beta-ratio was sensitive to workload manipulations, an index incorporating multiple frequency bands is likely more sensitive to workload changes than a single band.
Limitations and Future Research
There were limitations to this study that should be addressed. First, only frontal EEG sites were examined. While studies have shown changes in frontal EEG variables in response to workload manipulations, researchers have also advocated that changes in processing demands activate distributed cortical networks depending on the type of task (Just & Varma, 2007). Therefore, the full range of potential neurophysiological responses associated with multiple cortical networks was not captured. Despite this, our analyses did reveal differences in EEG patterns across the sensory-domain manipulations of workload. Second, Study 1 tasks were singular tasks. While the tasks in Study 1 were selected to capture activation patterns in response to specific domains without the possibility of confounds due to multiple domains engaged, MRT, as a whole, is the most germane under multitask situations. Therefore, while support for neurophysiological underpinnings of discreet tasks related to resource domain were found, to fully understand these implications for MRT, future work should carefully manipulate multitasking by including different sensory domains to establish better mapping onto operational tasks, while also examining other physiological measures. For example, measures of CBF taken from several cortical areas could be used to further understand the measurement of workload from several domains when combined with EEG. Functional near infrared spectroscopy (fNIRS) is a portable neuroimaging technique that provides cost-effective cerebral hemodynamic measures (i.e., oxygenated and deoxygenated hemoglobin). Increased workload has been associated with increases in oxygenated hemoglobin, particularly within frontal regions (e.g., Bunce et al., 2011; Herff et al., 2013). Combining cerebral hemodynamic information with EEG may provide greater diagnosticity of workload across sensory domains given the added features. Liu et al. (2017) found that combining fNIRS and EEG to classify workload states significantly outperformed EEG or fNIRS alone. The feasibility of incorporating both EEG and fNIRS recordings into operational environments should be explored. Additionally, dry electrode technologies may offer a more fieldable option than wet electrode systems (Di Flumeri et al., 2019).
Finally, another limitation to this study is that two different sets of participants were used for each study. Had the same participants completed each study, individual differences as a possible reason for different EEG outcomes found between studies would have been eliminated (Johnson et al., 2011). Moreover, comparisons could have been made on the individual level between each study. Future studies are recommended to take this approach to increase interpretability of findings.
Conclusions
Initial evidence for a difference in EEG oscillations based on workload manipulation within sensory domain at the basic task level was found to support the tenet of domain-specificity of cognitive resources within the MRT. According to the multiple resource model of attentional resources, there are several, qualitatively separate resource pools. Moreover, Wickens (2008) asserted that the multiple resource model should have “neurophysiological plausibility,” meaning that distinct cortical networks underlie qualitatively different resource pools. However, we were unable to find the same patterns during the second study where, although workload was manipulated in a domain-specific manner, the overall task utilized multiple domains. Based on these results, measuring distinct physiological changes in response to workload demands within a resource domain is plausible, but that other measures, such as fNIRS, as well as inclusion of other brain regions, should be explored.
Key Points
Frontal alpha and beta activities may play a role in differentiating workload within the auditory domain, although may differ based on singular task manipulations or multitask manipulations.
Psychophysiological indices to identify workload within flight should continue to evaluate whether there are sensory domain-specific responses to workload changes through the inclusion of additional measurement techniques (e.g., fNIRS) and cortical regions (e.g., parietal).
EEG oscillations during domain-specific tasks support the domain-specificity of resources suggested by the MRT.
Footnotes
Author(s) Note
The author(s) of this article are U.S. government employees and created the article within the scope of their employment. As a work of the U.S. federal government, the content of the article is in the public domain.
Author Biographies
Kathryn A. Feltman is currently a research psychologist with the U.S. Army Aeromedical Research Laboratory. She obtained a PhD in experimental psychology from the University of North Dakota in 2016.
Kyle A. Bernhardt is affiliated with the U.S. Army Aeromedical Research Laboratory through a fellowship supported by the Oak Ridge Institute for Science and Education. He is currently a PhD student with an MA in experimental psychology from the University of North Dakota obtained in 2018.
Amanda M. Kelley is currently the chief of the Aeromedical Factors Team at the U.S. Army Aeromedical Research Laboratory. She obtained a PhD in experimental psychology from Bowling Green State University in 2007.