
Abstract
Objective
This study used electroencephalography to explore the behavioral and electrophysiological effects of task interruption on performance.
Background
Task interruption is known to harm work performance, especially on working memory-related tasks. However, most studies pay little attention to cognitive processes by exploring brain activity and ignore the cumulative effect of sequential interruptions.
Method
Thirty-four healthy participants performed a spatial 2-back in three conditions: (1) interruptions with simple math questions, (2) suspensions with prolonged fixation cross, and (3) a pure 2-back. The measured outcomes comprise performance data, ERP amplitudes, EEG power, and subjective workload.
Results
Work performance decreased in the resumption trials, and cumulative interruptions had a more destructive effect on performance. EEG results showed that the P2 and P3 amplitudes induced by the 2-back task significantly increased after interruptions; theta and alpha power increased after interruptions. The P3 amplitude and alpha power induced by interruptions were significantly higher than that induced by suspensions.
Conclusion
Behavioral data revealed the disruptive effect of interruptions on postinterruption performance and the cumulative effect of interruptions on accuracy. Changes in ERP amplitudes and EEG power indicate the mechanisms of attention reallocation and working memory during interruptions. Larger P3 amplitudes and alpha power after interruptions than after suspensions suggested the inhibition of irrelevant information. These results may support the memory for goals model and improve the understanding of the effects of interruption on working memory.
Application
Focusing upon the mechanisms at play during the interruption process can support interruption management to ensure work safety and efficiency.
Introduction
An interruption is defined as the suspension of a primary task before its completion with the intent of returning to and completing the original work. Considerable evidence indicates that task interruptions are disruptive (Trafton & Monk, 2007) and are generally considered to exert negative effects on performance and increase the completion time and error rate of primary tasks (Bailey & Konstan, 2006), which results in a higher cognitive workload (Lin et al., 2013). Interruptions may lead to safety accidents (Couffe & Michael, 2017). For example, the accident reports of Eastern Air Lines Flight 401 indicated that interruption exerts considerable effects on pilots and increases the likelihood of accidents (Aircraft accident report, Eastern Air Lines, Inc. Lockheed L-1011, N310EA, Miami, Florida, 1973).
Most of the current research on interruptions has focused on the effects of interruptions on behavioral performance, and several characteristics modulate interruptions (Borghouts et al., 2020; Lee & Duffy, 2015; Monk et al., 2008; Pankok et al., 2017). Few researchers have examined relevant cognitive processes (Werner et al., 2015), and few studies have investigated the neural mechanisms in the brain. This study aims to identify the reasons for the observed decline in working memory (WM) performance caused by interruptions. We hope to understand more details of the interruption process and explore the underlying mechanisms at the neurophysiological level. Understanding the cognitive process during an interruption and how the harmful effects of interruption on performance are generated is necessary to improve performance after task interruption.
Trafton et al. (2003) proposed several stages of the task interruption process, as shown in Figure 1. Interruption lag refers to the time between the presentation of an interrupting stimulus and the beginning of the interruption task. Resumption lag is the time between the end of the secondary task and the recommencement of the primary task. It is generally believed that attention and WM play essential roles in an interruption process. Altmann and Trafton (2002) proposed the memory for goals (MFG) model based on the WM mechanism, which describes WM encoding and resumption of the interrupted task pertaining to the goal. Memory theory is the core of the MFG model, and each subtask is characterized by a goal. A suspended goal must be primed by a retrieval cue to overcome the interference from residual memories concerning old goals. The goal retrieval time affects the length of the resumption lag, which alters performance after an interruption task (Lin et al., 2013). Therefore, it is necessary to understand the time course of recovery after task interruption (Altmann & Trafton, 2007). Timeline of the task interruption process proposed by Trafton et al. (2003).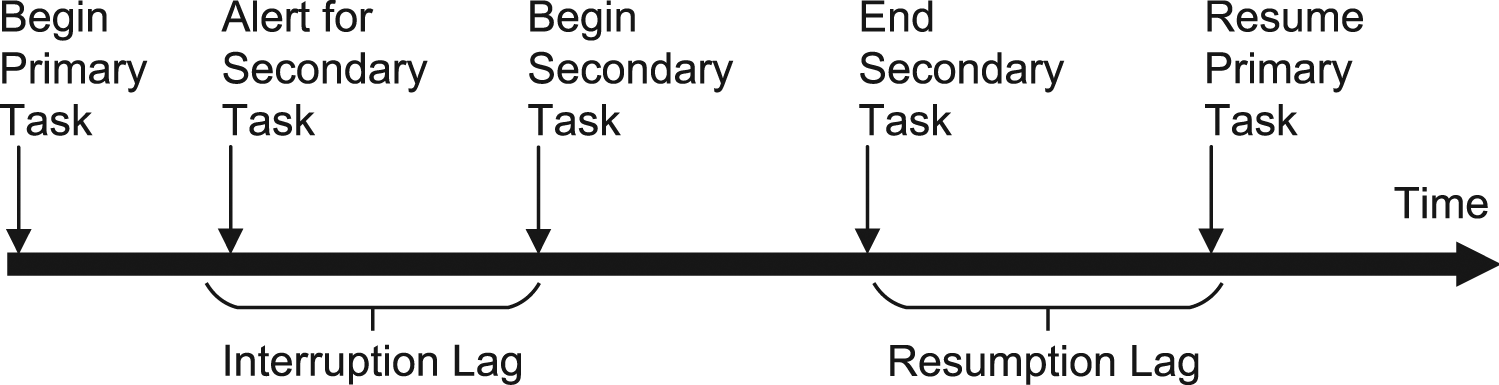
The decline of performance after an interruption is due to decreased memory activation of the primary task (Altmann and Trafton, 2002).
Notably, task interruption is not an isolated phenomenon. Sequential occurrence of interruptions is common in daily work (Baethge et al., 2015). However, there is little research to explore whether sequential interruptions have a cumulative effect. Although some researchers performed sequential interrupted tasks in their experiments, such as Altmann and Trafton (2007) interrupted participants up to 12 times in a computer game, Zickerick et al. (2020) had 25% interference trials in all trials, the additional effect of multiple interruptions on performance has not been noticed. Baethge et al. (2015) provided insight into the interruption accumulation process and explained how cumulative interruptions could lead to qualitatively different effects. Unfortunately, this study has no quantitative data to support this effect more effectively. In addition, what happens to attentional resources and WM during the interruption, and how does it affect WM performance after the interruption? To date, the underlying neural mechanisms of task interruption are not fully understood. Clapp et al. (2010) studied the mechanisms of WM disruption by external interference, including distraction and interruption. The results suggested that, unlike the encoded items that are maintained during distraction, the memory is not maintained when the participant is interrupted but is reactivated during the period after the interruption. The interfering stimuli of interrupters exerted a more detrimental effect on WM performance than distractors. Zanto and Gazzaley (2009) used a visual scene delayed-recognition paradigm and indicated that optimal WM performance depended on the ability to effectively filter irrelevant information via neural suppression to prevent the overloading of a limited memory capacity. Zickerick, Rösner, et al. (2020) examined the effect of interruptions and distractions on attentional and WM processes and indicated that a person’s focus on primary task information was impaired following an interruption, which manifested as reduced alpha lateralization. Another study by Zickerick et al. (2020) indicated that participants could successfully reallocate attention when back to the primary task after interference, which is reflected in the increased P3 amplitude. Berry et al. (2009) suggested that practice-related changes in processing interference positively affected WM performance. These studies focused on the differences in interference types (distraction vs. interruption). The terms “distraction” and “interruption” are often used interchangeably in everyday language, but they are different concepts (Boehm-Davis & Remington, 2009; Parasuraman et al., 1996). However, the changes in the neural activity of the brain during the interruption process are not clear. Rejer and Jankowski (2016, 2017) analyzed brain activity patterns associated with interruptions of the cognitive process by showing internet advertisements during a text-reading task. However, average signal power differences in three stages of task interruption (i.e., before, during, and after) were only observed in individual subjects.
The present study focused on the interruption process. To address these gaps mentioned above, we explored the differences before and after an interruption using the indexes of behavioral performance and specific ERPs and EEG bands. The cumulative effect of sequential interruptions is studied through behavioral data analysis by behavioral performance. Task interruption relates to the inhibition of irrelevant information, task switching, and WM retrieval, which are core aspects of executive control (Van der Linden et al., 2003). The 2-back task is a classic WM paradigm that involves online monitoring, updating, and manipulation of remembered information. This paradigm has been used to investigate the neural basis of WM processes (Owen et al., 2005). We used a 2-back task as the primary task and added math task interruptions. According to the goal activation theory of the MFG model (Altmann & Trafton, 2002), a long delay will cause memory decline. The evidence presented in previous literature suggests that interference from irrelevant information discourages WM reactivation. Is forgetting from WM better explained by decay or interference? The answer to this question is also the topic of ongoing debate in WM research (Portrat et al., 2008). To clarify that the reason for memory decline after the interruption is time-related decay or interference-based forgetting, we design task suspension without additional interference and action using the prolonged fixation cross. Healthy volunteers were recruited to complete the n-back task, and their behavioral performance, ERP amplitudes, and EEG power were collected.
We focused on the P2 and posterior P3 components of ERP because these components were previously shown to be evoked by the n-back task (Han et al., 2013) and modulated by attentional allocation and WM (Lin et al., 2020; Polich, 2007). The P2 component is a core aspect of many cognitive tasks, including the n-back task, and it reflects the early stage of information processing (Ciecko-Michalska et al., 2012; Huang & Luo, 2007). It is generally believed that P2 is related to stimulus detection, stimulus switching, and attentional resources (Kieffaber & Hetrick, 2005; Luck, 2014; Potts, 2004). P2 responses are modulated by cognitive load (Vilà-Balló et al., 2018), and an increase in WM load demand leads to an increase in P2 amplitude (Yuan et al., 2016). Many studies have confirmed that P3 is related to resource allocation, memory storage, and inhibition processing and serves as a link between stimulus features and attention (Elward et al., 2013; Patel & Azzam, 2005; Wiswede et al., 2007). With the increase in memory load, subsequent attention resources increased P3 amplitudes due to the high resource demand for task processing (Kok, 2001). An inhibition hypothesis of P3 has been confirmed by many studies (Chikara et al., 2018; Polich, 2007; Roberts et al., 1994). Smith et al. (2013) have shown that the NoGo effect of P3 is more pronounced under the condition of press response because the participants need to inhibit their external response; this inhibition is one of the reasons for the production of P3. The P3 amplitude is related to the inhibitory ability. The stronger the inhibitory ability, the greater the P3 amplitude (Smith et al., 2006). Randall and Smith (2011) found that the P3 amplitude of the correct response was greater than the wrong response under NoGo conditions, indicating that the P3 amplitude would increase with the enhancement of the inhibitory control.
Theta and alpha oscillations have been shown to track WM-related processes (Popov et al., 2018; Riddle et al., 2020). Some evidence suggests that theta oscillation is proposed as a neural mechanism supporting WM (Sauseng et al., 2010), and alpha oscillation supports attentional orientation and inhibitory function (Klimesch, 2012). The n-back task has found that increased theta power at frontal electrodes accompanies increased memory load (Zhao et al., 2017). Theta oscillations are sensitive to the amount of information (Jensen & Tesche, 2002). The increase in theta power correlates with episodic long-term and working memory (Klimesch, 1999). Theta power was observed to increase when information encoding was sustained during information maintenance (Raghavachari et al., 2001). Much evidence has demonstrated that alpha power increases with WM load (Jensen et al., 2002), but some researchers have obtained opposite modulations of alpha activity (Tsoneva et al., 2011). This difference may be related to the different nature of memory tasks (Popov et al., 2018). More studies on alpha oscillations in the control of WM suggested an association between alpha activities and local cortical inhibition driven by cognitive control systems (Bonnefond & Jensen, 2012; Clayton et al., 2015; Wöstmann et al., 2019).
In summary, this study attempts to determine the cumulative effect of sequential interruptions on behavioral performance and explores the electrophysiological mechanism correlates of attentional control and WM processes underlying the impact of interruptions on subsequent performance. The working memory of the suspended goal for the primary task is retrieved and restored following interruptions according to MFG (Altmann & Trafton, 2002). People shift their attention from the interruption task to the primary task and inhibit the information interference raised from residual memory of the interruption task. Based on these findings, we assume that (1) sequential interruption exerts a negative effect on WM performance; (2) P2 and P3 amplitudes in trials after an interruption would be significantly increased compared with trials before an interruption, and P3 amplitudes would be larger in trials after an interruption than in trials after a suspension; and (3) interruptions lead to higher WM loads than suspensions and noninterrupted tasks, accompanied by an increase in theta power, and increased demand for information inhibition after interruption results in larger alpha power.
Methods
Participants
This research complied with the American Psychological Association Code of Ethics and was approved by the Human Factors and Ergonomics Laboratory, Beijing Jiaotong University. The experiment was performed with 34 healthy adults (19 males and 15 females) aged 22–29 years. All participants were right-handed and had no behavioral or cognitive impairment. Before the experiment, each subject volunteered, provided consent, and was fully informed about the experiment. One subject withdrew from the experiment due to physical discomfort. Therefore, the final sample size was 33 (Mage = 23.94, SDage = 1. 63).
Stimuli and Experimental Design
We designed three tasks (interruption, suspension, and baseline), which are shown in Figure 2. All three tasks were designed based on the 2-back spatial task in which a black square was shown for 500 ms at one of eight predefined locations except the center. A fixed cross was presented in the center of the screen for 2000 ms between two trials. Participants were asked to decide whether the location of black square in each item matched the one that was presented 2-items ago. A secondary math task requiring addition within 100 appeared on the screen as the interruption. Each task interruption contained 5 math questions. A math question was presented for 500 ms, followed by a fixed cross for 2000 ms. Participants were asked to judge whether the math answer was greater than or equal to 50. After completing the interruption of 5 math questions, the 2-back task was resumed. The participants were required to determine whether the current stimulus matched the stimulus from two steps earlier in the sequence and ignore the interruption trials (math tasks). The only difference in the suspension task from the interruption task was that the 2-back task was suspended by a prolonged presentation of the fixation cross. The prolonged fixation cross had the same duration as the five math questions. A pure 2-back task was also performed as the baseline condition. Baseline data were then used to compare task interruption and suspension task outcomes. Experimental design based on the spatial 2-back task. Three WM tasks were included in the experiment. The primary task (2-back) in the task interruption condition was interrupted by five math tasks (addition in which the sum was less than 100; participants decided whether the sum of the math equation was greater than or equal to 50), and the 2-back task resumed. The primary task (2-back) in the suspension condition was suspended by the prolonged presentation of the fixation cross (same duration as the math tasks). A delay occurred before participants were allowed to resume the 2-back task. Only a 2-back task was performed in the baseline condition.
Two resumption trials and twelve pre-interruption trials were interspersed between two interruptions or suspensions. Since the primary task was the 2-back task, the two trials after the interruptions or suspensions were regarded as the trial type for resumption. The following 12 2-back trials before the next interruption or suspension were pre-interruption trials. Interruption and suspension conditions consisted of two blocks with 156 pre-interruption trials and 24 resumption trials each, in which a block contained 12 interruptions or suspensions. The baseline condition consisted of two blocks with 160 normal trials each. A 5 minute break was included between the two blocks to prevent mental fatigue. Participants were instructed to press the “f” key with their left hand for a match (correct answer for math tasks) and the “j” key with their right hand for mismatch (incorrect answer for math tasks) as quickly and accurately as possible. The ratio of match trials and mismatch trials was 1:1, which was also the ratio of correct and incorrect trials for math tasks.
Procedures
This study used a within-subject experimental design. Participants were first trained for 30 minutes and were provided feedback on their performance (Herzog & Fahle, 1997) to ensure that their performance did not increase significantly during the formal experiment. The order of task execution was random. Before each condition in the formal experiment, participants were informed of the task type to be performed. After each condition, the NASA Task Load Index (NASA-TLX) scale was completed to evaluate the subjective workload (Cao et al., 2009).
Apparatus
All participants performed the task on a Core i7-4770K, 16 GB memory computer using a 22-inch monitor (60 Hz refresh rate; 1920 × 1200 pixels) placed at 70 cm distance from participants. Participants responded with button presses on a standard keyboard that was placed in front of them.
EEG Recording and Analysis
The experiment was performed in a normally illuminated room with electrical shielding. EEG signals were continuously recorded from 64 scalp locations distributed according to the international 10–20 system using Ag/AgCl electrodes. Vertical electrooculogram (VEOG) data were recorded from electrodes above and below the left eye. Horizontal electrooculogram (HEOG) signals were monitored by placing electrodes at the outer canthi of both eyes. Scalp recordings were referenced online to the electrode between Cz and CPz and were rereferenced to the average of the left and right mastoids via offline algebraic computations. The impedance of all of the electrodes was maintained at less than 5 kΩ. All EEG signals were amplified by a SynAmps2 amplifier (Neuroscan Inc., USA) and digitized at a sampling rate of 1000 Hz.
Descriptive statistics for the number of trials in each condition (mean ± standard deviation)

We analyzed the ERP components of P2, P3, and EEG oscillations of theta (4–7 Hz) and alpha (8–12 Hz). We determined time windows centered on the peak by visually inspecting individual data and measured local peak amplitude as defined by Luck (2014). The mean amplitudes of the P2 and P3 components were measured separately during the time windows of 180–280 ms and 300–450 ms time-locked to the onset of all 2-back stimuli. Statistical analyses were performed on multiple electrodes: frontal electrodes at F3, Fz, and F4; central electrodes at C3, Cz, and C4; and parietal electrodes at P3, Pz, and P4 (Forester et al., 2020; Vilà-Balló et al., 2018). The theta and alpha bands in the time window of 0–1000 ms poststimulus onset were analyzed separately at Fz, Cz, and Pz with a baseline window of −500 ms to −200 ms (Boksem et al., 2005). The relative wavelet energies of the theta and alpha bands were analyzed using wavelet packet time-frequency analysis (Ting et al., 2008). Daubechies db4 was selected as the mother wavelet because of the high signal-to-noise ratio value (Balasubramanian et al., 2018). One-way repeated-measures analysis of variance (rm-ANOVA) was used to analyze the effect of the task type (interruption vs. suspension vs. baseline) in pre-interruption trials. The 2 (task type: interruption vs. suspension) × 2 (trial type: pre-interruption vs. resumption) two-way rm-ANOVA was performed to analyze the main effects and interaction effects on ERP amplitudes and EEG power. Analyses were limited to correct trials.
Behavioral data included the response time (RT) and accuracy of 2-back tasks. Using the NASA-TLX, we calculated the subjective mental workload. We performed three separate one-way rm-ANOVAs to analyze the task type effect on overall performance, including RT and accuracy, subjective workload, and two separate two-way 2 (task type: interruption vs. suspension) × 2 (trial type: pre-interruption vs. resumption) rm-ANOVAs on RT and accuracy.
The reported p values for the significance level of all rm-ANOVAs results were set at 0.05, and effect sizes were calculated using partial eta squared (
Results
Behavioral Data
Descriptive statistics for behavioral data (means ± standard deviations). The behavioral data show performance on 2-back trials, including the RT and accuracy of pre-interruption and resumption trials in task interruption and suspension conditions and weighted average performance
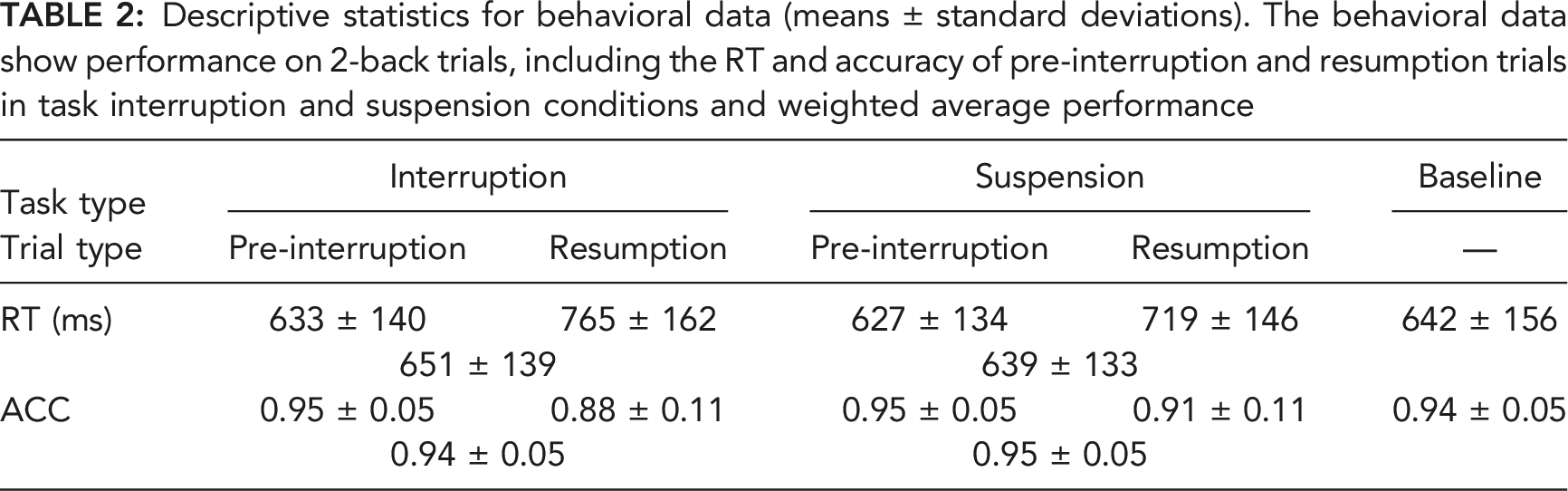
A 2 × 2 rm-ANOVA was performed with the task type and trial type, as shown in Figure 3. The main effects of task type, F (1, 32) = 5.362, p = .027, Behavioral data. Simple two-way interaction effect of the task type and trial type on RT and accuracy. Error bars indicate 95% confidence intervals (CIs).
Significant main effects of the task type, F (1, 32) = 9.153, p = .005,
Figure 4 shows the RT and accuracy of serial trials after interruptions or suspensions, including two resumption and twelve pre-interruption trials under interruption and suspension conditions. A 2 task type × 14 trial position rm-ANOVA revealed main effects of position on RT, F (1, 32) = 54.650, p < .001, RT and accuracy of each resumption trial and pre-interruption trial under the task interruption and suspension conditions. Positions 1 and 2 are resumption trials, and positions 3 to 14 are pre-interruption trials before the next interruption or suspension. Error bars represent 95% confidence intervals (CIs).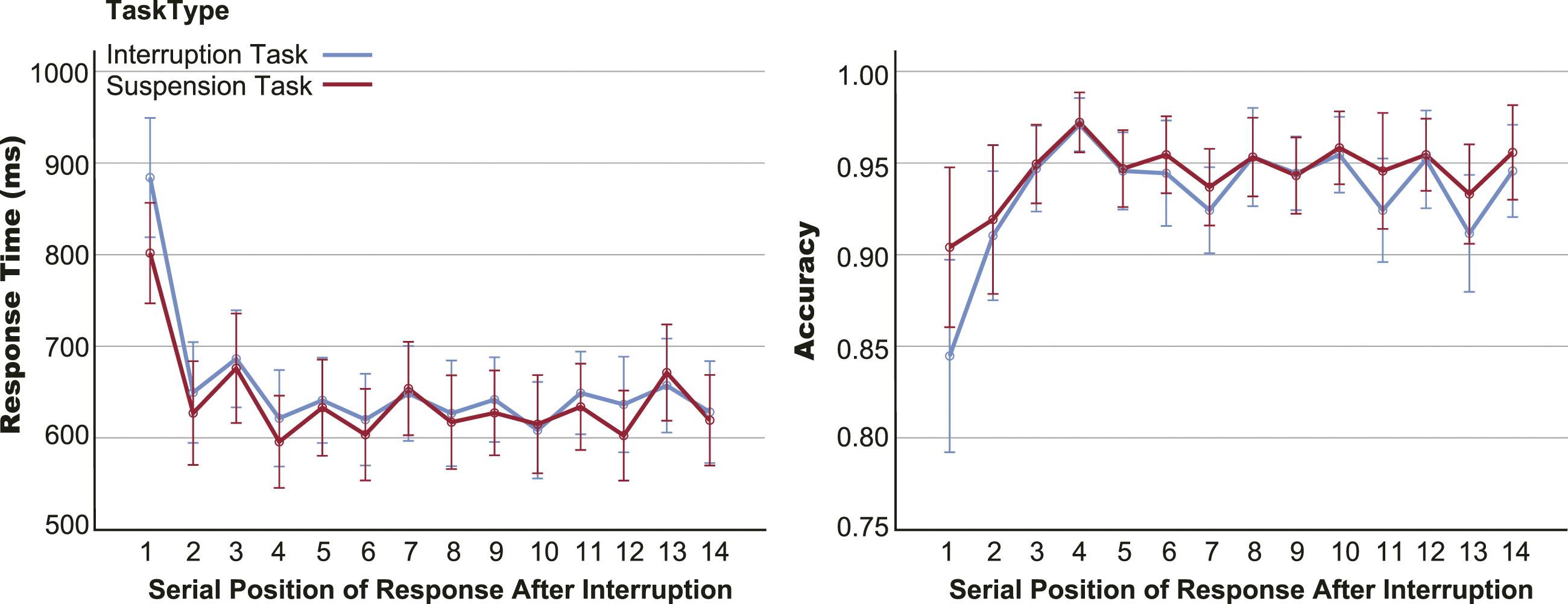
To explore the impact of cumulative interruptions on performance, we assessed whether a difference in performance was present as interruptions or suspensions increased, as shown in Figure 5. The 2 task type × 12 times rm-ANOVA results did not reveal significant differences in RT after multiple interruptions or suspensions, F (11, 352) = 1.635, p = 0.136, RT and accuracy for all trials (resumption and pre-interruption trials) with multiple interruptions or suspensions. Error bars are 95% confidence intervals (CIs).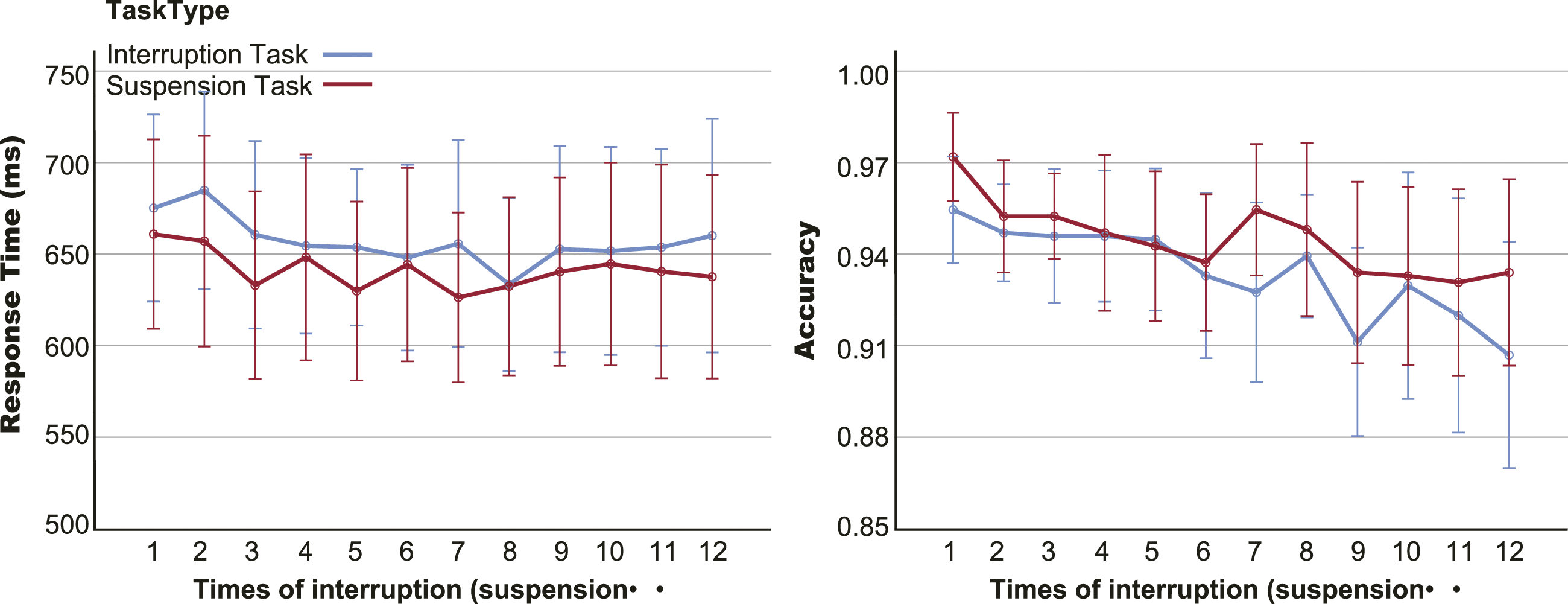
ERP Results
Mean P2 Amplitude
We first investigated differences in P2 amplitude in pre-interruption trials among three task types. The results indicated that the mean P2 amplitude did not vary with the task type in pre-interruption trials at the frontal, F (2, 64) = 1.063, p = .351,
Two-way rm-ANOVA results for P2 and P3 amplitudes across electrode sites. ***p < .001, **p < .01, and *p < .05
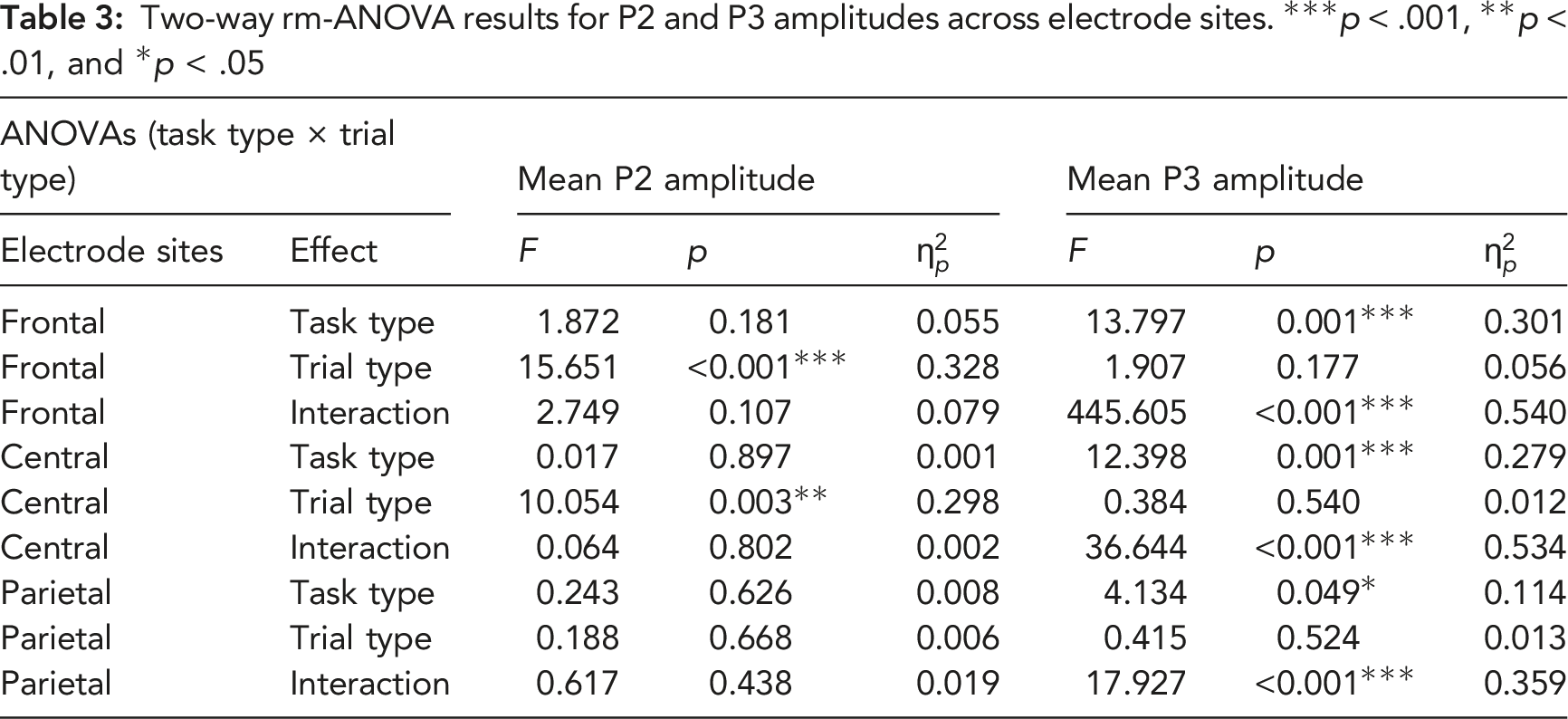
Grand averages of P2 and P3 ERP components at frontal, central, and parietal electrodes. The black line illustrates all trials of the baseline task. The blue and red dashed lines depict the pre-interruption trials for interruptions and suspensions. The blue and red solid lines illustrate the resumption trials for interruptions and suspension. (B) The scalp topographies display the mean amplitude recorded under each condition between 180–280 ms for P2 and between 300–450 ms for P3.
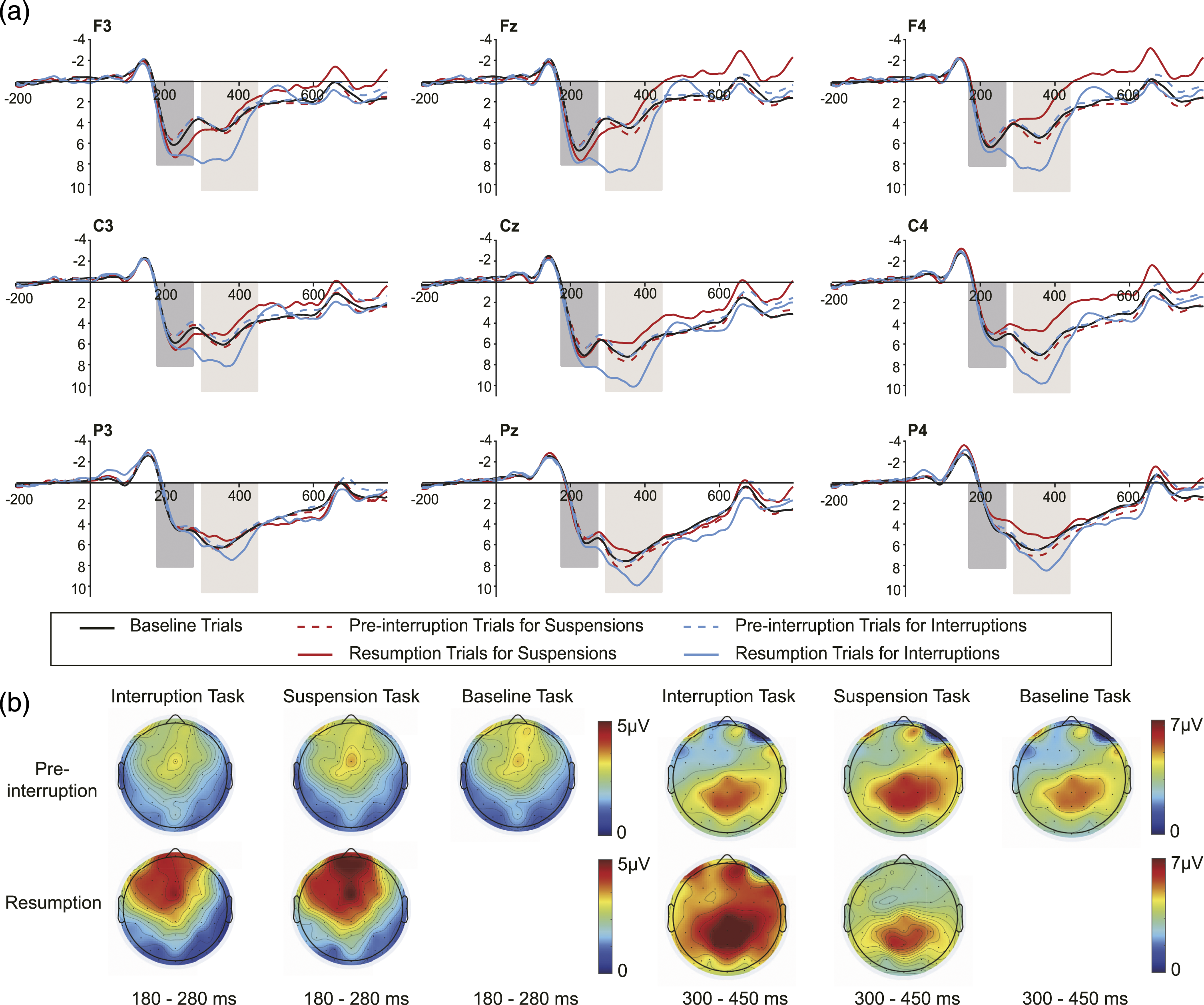
Mean P2 and P3 amplitudes of each condition at (A) frontal, (B) central, and (C) parietal electrodes. Error bars represent 95% confidence intervals (CIs).
Mean P3 Amplitude
One-way rm-ANOVA results did not reveal a significant main effect of task type at the frontal, F (2, 64) = 1.644, p = .206,
At frontal sites, the main effects of trial type and the interaction effect were significant. The simple effect analyses were performed. Resumption trials elicited larger P3 mean amplitudes in interruption conditions than pre-interruption trials, F (1, 32) = 32.308, p < .001,
At central sites, we identified a significant main effect of trial type and interaction effect. Subsequent analyses showed that task interruption elicited larger P3 amplitudes in resumption trials than in pre-interruption trials, F (1, 32) = 27.074, p < .001,
At parietal sites, we detected a significant main effect of the trial type and interaction effect. No significant decreasing trend in P3 amplitudes was observed for suspensions in resumption trials compared with pre-interruption trials, F (1, 32) = 3.042, p = .091,
EEG Power
Theta
Figure 8 illustrates the topographic distribution of relative theta power for each condition, and Figure 9 shows the statistical results. One-way rm-ANOVA results showed that there was no significant difference in pre-interruption trials among the three tasks (interruption, suspension and baseline), F (2, 64) = 0.983, p = .380, Topographic representation of the grand average of relative power in theta and alpha bands. The baseline task illustrates all trials of the 2-back task. Interruption and suspension tasks illustrate pre-interruption, during, and resumption trials. The during trials were math tasks for interruptions and prolonged fixation crosses for suspensions. Statistical results of relative theta and alpha energy. The during trials were math tasks for interruptions and prolonged fixation crosses for suspensions. Error bars represent 95% confidence intervals (CIs).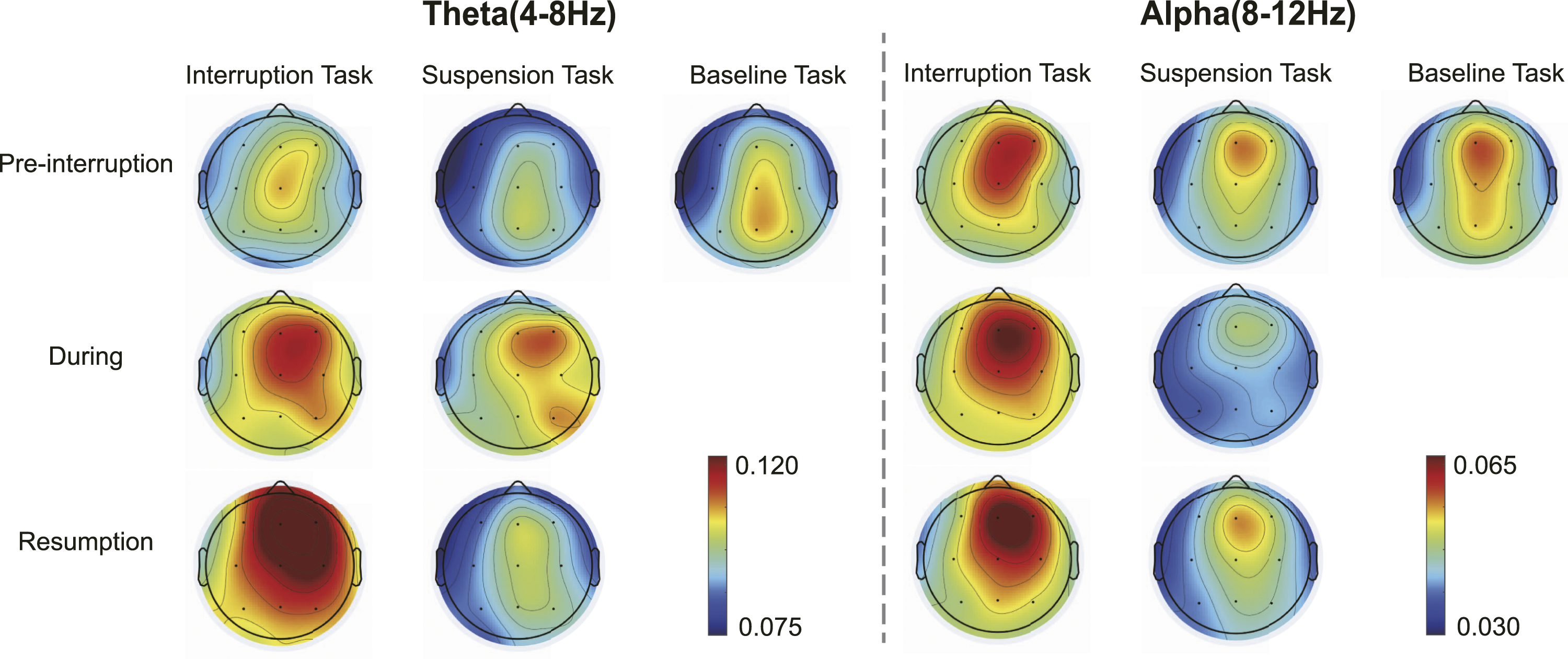
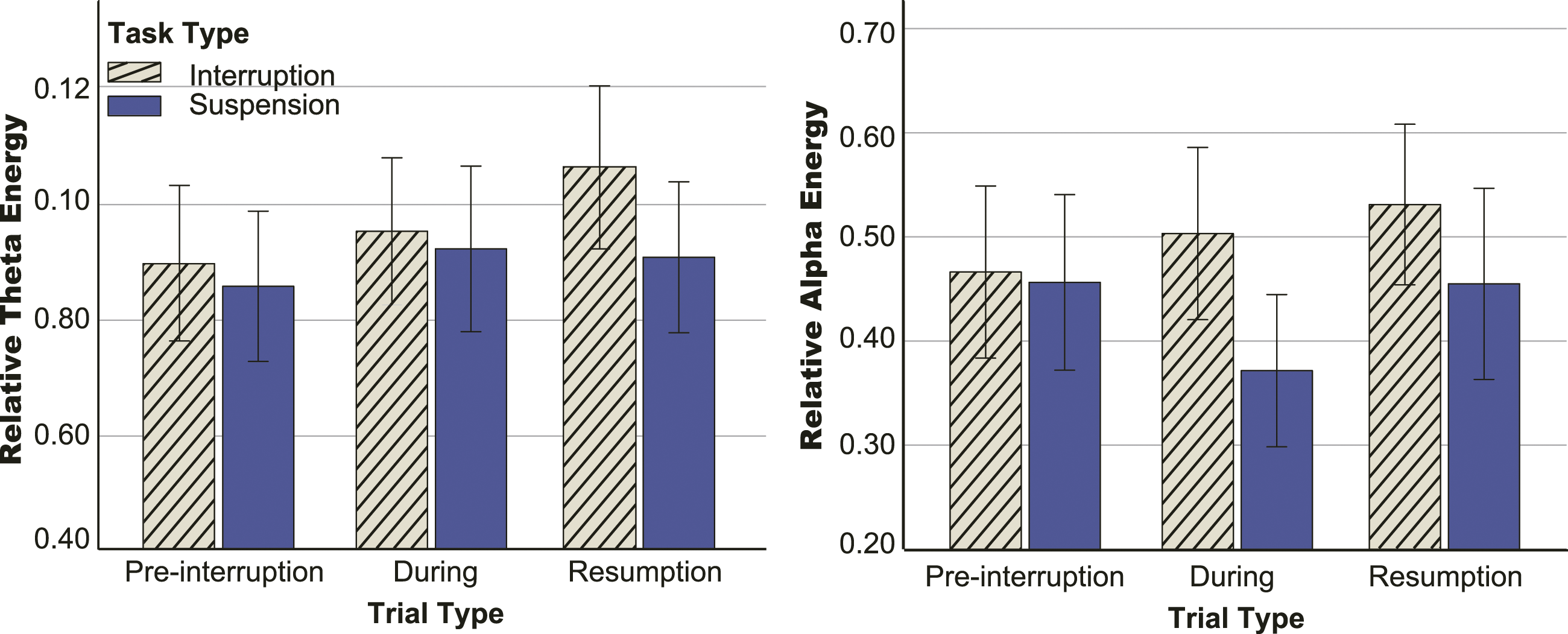
The 2 (task type: interruption vs. suspension) × 2 (trial type: pre-interruption vs. resumption) two-way rm-ANOVA results showed the main effects of task type, F (1, 32) = 9.549, p = .004,
To further test the changes in EEG oscillation during interruptions (math tasks) and suspensions (prolonged fixation cross), a 2 (task type: interruption vs. suspension) × 3 (trial type: pre-interruption vs. during vs. resumption) rm-ANOVA was performed. The main effects of task type, F(1, 32) = 5.462, p = .024,
Alpha
One-way rm-ANOVA results showed that there was no significant difference in pre-interruption trials among the three tasks, F (2, 64) = 0.149, p = .862,
Subjective Workload
The subjective workload evaluation was obtained from NASA-TLX after completing each condition. One-way rm-ANOVA was performed and revealed the main effect of the task type on subjective workload, F (2, 64) = 30.660, p < .001,
Discussion
This study examined the effects of interruption on WM performance using spatial n-back tasks, and neural activity was recorded during the interruption. The statistical results did not reveal a disruptive effect of interruptions of overall WM performance, including RT and accuracy. However, we found that interruptions damage the subsequent performance, and the damage is mainly focused on the first trial of resumption. Cumulative interruptions reduced the accuracy of overall WM performance as the number of interruptions increased. The differences between trial types and task types by analyzing the amplitudes of P2 and P3 components support our hypotheses, interruption evoked greater P2 and P3 amplitudes in resumption trials, and interruptions and suspensions produced different P3 amplitudes in resumption trials. EEG power analysis showed that theta was greater for interruptions than suspensions and for resumption trials compared to pre-interruption and during trials. Alpha power increased in resumption trials under interruptions compared to suspensions, further confirming the mechanism of information interference in the interruption process. These results clearly reflect the effect of interruption on attention processing and WM.
At a behavioral level, in contrast to most previous interruption studies (Puranik et al., 2020), we did not find that interruptions decreased overall primary task performance. Based on the 2×2 rm-ANOVA results, task performance decreased in the resumption trials compared to pre-interruption trials under the interruption and suspension conditions. An interruption in the process of memory coding is generally predicted to distract attention, weaken memory coding processing, and lead to poor memory behavior (Cowan, 1998; Mulligan, 1998). Detailed analyses of each trial revealed that the decreased performance on resumption trials mainly occurred in the first resumption trial. The increased RT recorded in the first resumption trial was likely due to the existence of a resumption lag after the interruption, which may reflect a process of memory activation (Altmann & Trafton, 2002). This finding is consistent with Altmann and Trafton (2007), who measured the time course of responses following an interruption.
Interestingly, performance on pre-interruption trials under interruption and suspension conditions was slightly higher than that on the baseline task (Table 2). This finding indicated that an interruption might lead to a phenomenon of speedup after interruptions (Trafton & Monk, 2007). Ratwani et al. (2006) argued that speedup after interruptions was attributable to perceptual speedup rather than a response of the cognitive system. However, we found that the speedup persisted in most pre-interruption trials, and this result was more likely due to a compensatory mechanism (Zijlstra et al., 1999). According to the compensatory control model (Hockey, 1997), the “effort monitor” is sensitive to increasing the control demands. After an interruption, active control improved performance, including increased working memory or executive control. This finding also supports studies that proposed that a simple interruption task positively affected performance (Speier et al., 2003). Notably, speedup requires greater effort to maintain performance, which increases subjective mental workload (Fonseca et al., 2018). The subjective mental load of the interruption condition was the highest, followed by the suspension condition. Even if there are no secondary tasks, the speedup effect caused by interrupted tasks makes its workload higher than the baseline.
Next, let us focus on the effect of cumulative interruptions on performance. Our experimental design included 12 interruptions or suspensions in one block; this is a cumulative interruption because multiple sequential interruptions occur over a period of time. Baethge et al. (2015) think that cumulative interruptions exerted stronger effects on WM performance than a single interruption, reflected in many aspects, such as time pressure, error, reaction time, and emotional states. Our results showed this cumulative effect of multiple sequential interruptions on accuracy. As described previously, the additional compensatory responses after each interruption result in great resource depletion (Hockey, 1997). According to conservation of resources theory (Hobfoll, 1989), mobilizing additional effort cannot continue indefinitely. Resource recovery takes time (Sonnentag & Zijlstra, 2006), which is not allowed in sequential interruption tasks. Consequently, cumulative interruptions gradually reduced the accuracy. However, we did not observe a cumulative effect on RT. One possible explanation is that the primary and secondary tasks are simple and repeated. Participants knew that interruptions or suspensions would occur in the subsequent tasks, and therefore, they had psychological preparation to respond as soon as possible.
The ERP and EEG results indicated a significant change in brain activity after an interruption compared to the activity observed during the primary task. The P2 component is often distributed around the central frontal areas and likely reflects the initial stage of context updating in the n-back task (Lenartowicz et al., 2010). Our results revealed that interruptions led to increased P2 amplitudes at frontal-central sites in resumption trials, which reflected context updating of task switching. The interruption is closely analogous to the exogenously instructed task switch. The difference is that participants must return to the primary task after interruptions, but the task switch is not necessarily (DARMOUL et al., 2015). Some task switch researchers reported that the P2 amplitude was diminished or absent in repeated trials (Rubin & Meiran, 2005). Therefore, resumption trials with longer ISIs elicited larger P2 amplitudes than pre-interruption trials in suspension conditions. The P2 amplitude is related to relevant stimuli, which show enhanced prefrontal positivity to the instructed targets (Potts et al., 2004). This finding explained the lack of change in the stimulus interval of the task interruption condition (the five math interruption trials were not relevant to the primary task) and the increase in the P2 amplitude of resumption trials after interruption. The P2 amplitude also reflects the early allocation of attention (Crowley & Colrain, 2004) and memory activation (Lenartowicz et al., 2010; Sakai, 2003). WM might have been jeopardized due to interruption, causing more attentional resources in processing and updating the resumption stimulus for WM reactivation and greater P2 amplitudes.
For the P3 component, our result is consistent with previous studies, such as Vilà-Balló et al. (2018) and Zickerick et al. (2020). Both studies suggested a pronounced increase in the amplitude of the P3 component after interference. As mentioned in the literature review, most studies have confirmed that P3 elicited by WM tasks (e.g., n-back) was brain activity that reflected attentional resource and cognitive control processes. The P3 amplitude reflects the mental resources invested by the individual to a certain extent (Polich, 2007; Scharinger et al., 2017; Sumich et al., 2008). Notably, we found that interruption and suspension conditions showed different changes in trial types, especially in resumption trials. One hypothesis about P3 as an inhibitory component suggested that P3 and its underlying subprocesses reflected rapid neural inhibition of ongoing activity to promote the transmission of stimulus information from frontal (P3a) to temporal-parietal (P3b) locations (Polich, 2007). Many studies have confirmed that the P3 amplitude increases with increasing inhibitory functions of the brain (Randall & Smith, 2011). The suspended WM in this study had to be recalled to continue the primary 2-back task under the suspension condition. The WM needed to overcome additional interference from math tasks in the task interruption condition. Therefore, the brain devotes more resources to inhibit the processing of irrelevant information while focusing attention on the resumption stimulus (Soltani & Knight, 2000). This finding explains the increased P3 amplitude of resumption trials under interruption but not suspension conditions. This result is consistent with the theory that P300 changes with the workload. The primary task is simple, and a 2 s stimulus interval is sufficient. Additional attention resources may be used to prepare for subsequent task-related stimuli. With the increase in memory load, due to the increase in task processing demands, the subsequent attention resource engagement contributed to the increase in P300 amplitude (Kok, 2001; Vilà-Balló et al., 2018).
Katayama and Polich (1998) reported the P300 amplitude elicited by the distracter and target in the parietal electrodes for easy tasks, and a larger P300 amplitude distributed over the frontal/central electrodes was observed for difficult tasks, which is consistent with our results for P3 amplitudes over different electrode regions. The simple primary task primarily elicited central and parietal P300 in pre-interruption trials under three conditions and in resumption trials under the suspension condition. Participants reported that they performed mental rehearsal during the suspension, making the recall relatively simple after the suspensions. The task becomes difficult in resumption trials after the interruption, which elicited P300 in the prefrontal lobe. PFC retrieval and reactivation activities are also enhanced with the high demand for interference suppression in the task interruption condition (Sakai, 2003; Sakai & Passingham, 2004) (see Figure 6). Researchers found that the influence of interference from an irrelevant domain was larger in posterior areas than frontal areas (Pelosi et al., 1992), which supported the finding that the difference in P3 amplitude was greater than the difference in P2 amplitude. According to previous studies, the prefrontal-parietal network is closely related to attention and WM processes (Cabeza & Nyberg, 2000; Duncan, 2006). With the shift in attention and increased amount of information in WM, the activity of the prefrontal-parietal network also increases, which is consistent with the experimental results from the current study. The results indicated a critical role for the prefrontal-parietal network in the interruption process.
Theta and alpha bands play important roles in WM control (Schmiedt et al., 2005). Theta power increases as memory load increases (Sauseng et al., 2010). The increasing theta power gradually increased workload in pre-interruption, during, and resumption trials of interruption conditions. Only during trials showed higher power in the suspension conditions because the participants performed mental rehearsal for the upcoming two resumption trials. Alpha oscillations have always been associated with the inhibition of task-irrelevant information (Manza et al., 2014). The present results are consistent with the claim that greater alpha activity indicates an increase in cortical inhibition, reflected in the large difference in during trials between suspension and interruption conditions. It was necessary to inhibit interference information from the math tasks during interruptions but not during suspensions. Therefore, alpha power was reduced in suspension conditions.
Overall, these findings were consistent with the MFG model (Altmann & Trafton, 2002). The emergence of the interruption required attention resources to be allocated to address the secondary tasks, activated new goals, and depreciated the WM of the primary task. According to our comparative analysis of suspensions and interruptions, a long time interruption causes WM decay (Monk et al., 2004), and irrelevant information from secondary tasks exerts greater interference effects on WM. The results from alpha power and P3 amplitudes are consistent with this conclusion. The interference of irrelevant information must be suppressed, and attention must be refocused toward the primary task to recall WM after the interruption. This process led to an increase in RT and a decrease in ACC after the interruption. The interruption task also increased the subjective load.
Limitations
The current study has a few limitations. The primary task in our experiment was a 2-back spatial WM paradigm. As mentioned in the discussion, the task is simple, and interruption did not negatively affect overall performance. The applicability of the conclusion to interruptions in other paradigms and complex tasks must be verified. Task interruption is a problem that often occurs in a human-computer interaction environment. The effect of interruption must be verified in a more complicated environment in the future. The current study only analyzed physiological and cognitive mechanisms of interruption. Several characteristics modulate interruptions, such as task similarity (Oulasvirta & Saariluoma, 2004), complexity (Eyrolle & Cellier, 2000), interruption duration (Monk et al., 2008), and frequency (Basoglu et al., 2009), and the modulation mechanism requires further study. The fixed number of trials between two interruptions or suspensions may allow a certain expectation for the interruption, although we did not tell the participants about this. Even if the interruption occurs but cannot be accurately predicted (it is possible in theory, but participants have no time to address it during the actual experiment), the participant can roughly judge whether the interruption is about to occur. In upcoming experiments, we randomize the number of trials between interruptions. A problem worthy of further study is whether the predictability of the interruption affects the performance after the interruption.
Conclusions and Practical Implications
The current study revealed a harmful effect of interruption on WM in the resumption trials, and cumulative interruptions had a more destructive effect on WM performance. Mean amplitudes of P2 and P3 components changed significantly after the interruption, which was implicated in attention-related and WM-related neuroactivity. Increased P3 amplitudes and alpha power in interruption tasks illustrate that in addition to the decay of WM over time, the interference of irrelevant information exerted a stronger effect on resumption lag and WM performance. Interruptions increased WM load, manifested by an increase in theta. This research improves our understanding of the physiological and cognitive mechanisms during interruptions. Our findings provide better theoretical support for the MFG model and interruption management research.
Footnotes
Acknowledgments
Yue-Yuan Chen: Conceptualization, Methodology, Formal analysis, and Writing - Original draft preparation. Wei-Ning Fang: Supervision, Data curation, Validation, and Funding acquisition. Bei-Yuan Guo: Software, and Reviewing and Editing. Hai-Feng Bao: Software and Visualization. This work was supported by the National Natural Science Foundation of China (Grant Number: 72271015).
Key points
We investigated the effect of interruption on primary performance, ERP components and EEG bands. We explored the time course of resumption after interruptions and the cumulative effect of interruptions at the behavior level. The mechanism of interruption’s impact on performance relies on attentional reallocation and working memory. We verify the information interference mechanism of interruptions and support the MFG model.