
Abstract
The emergence and institutionalization of cooperation in sizable groups without reciprocity receives considerable attention in game-theoretical modeling. Agents in our study play the Prisoner’s Dilemma game cooperating with tolerably similar neighbors. They may imitate cultural markers (tags) and tolerance from more successful neighbors. Alternatively, they break ties to out-group neighbors. New partners are selected either from neighbors’ neighbors (clustering) or randomly. Variations in network plasticity (the likelihood of changing partners rather than being influenced) and clustering are explored. With high plasticity and high clustering, networks tend to fragment. With low plasticity and low clustering, networks tend toward global cooperation, but with severe losses of cultural diversity and tolerance. Cooperation in such regimes also proves to be vulnerable to defection. Between, there is a space displaying relatively stable and widespread cooperation with diversity and tolerance. We note some important structural characteristics of the networks evolving in this space.
Keywords
A regime that is not able to maintain relatively high levels of cooperation among most of its members may confront considerable disintegrative selection pressure. Cooperation, however, faces “social dilemma” (Dawes 1980; Glance and Huberman 1994; Kollock 1998) where the rational behavior of individuals aggregates to Pareto-inefficient outcomes. Under the circumstances, social norms render stable patterns of cooperative interactions possible. Direct reciprocity is one of the simplest explanations of where norms of cooperation come from (Axelrod 1984). However, it is not suitable for the evolution of cooperation in sizable populations consisting of those who rarely interact with the same partners.
It is a theoretical and empirical puzzle to explain the emergence of cooperation and its sustainment in the setting of one-shot interaction without reciprocity. If helping someone does not influence the likelihood of receiving help either from the recipient (i.e., “direct reciprocity”) or from third parties (i.e., “indirect reciprocity”), we now need to consider how cooperating with strangers is possible in spite of incurred costs. Rather than “altruistic” motivation or “general” trust, a common base for such cooperation may be in-group favoritism or “parochialism” (Bowles and Gintis 2004).
People may read observable markers (cues) to categorize others into “us” and “them” on the basis of perceived dissimilarities, that is, social distances. How can global cooperation emerge from local interactions of actors who may have limited tolerance for cooperating with others who they see as different from themselves? Parochial solidarity stops at group boundaries. Within-group cooperation and between-group noncooperation do not result in society-wide cooperation. We can expect that a society can achieve a high level of cooperation either when its members look very alike, lacking in cultural diversity (cf. “mechanical solidarity” in Durkheim [1893] 1933), or when more tolerant agents constitute a majority of the population who accept a broader range of members as in-group.
In-group favoritism along categorical boundaries enables discriminators to share disproportionate benefits. But, similarity-based parochial cooperation may be no longer stable against defectors when their markers are sufficiently alike within the tolerance range of the typical members of the dominant group. The failure of “secret hand-shaking” (Robson 1990) by which cooperators parochially help each other may lead to a serious downward spiral of cooperation. It is also expected that a more tolerant society is more likely to degenerate into a betrayal one because a wider pool of defectors are recognized as in-group.
People not only consider how to interact with partners but usually have the option of whether to interact with others. Such partner selection can maintain cooperation via “spatial reciprocity” if cooperators benefit from interactions with their own kind while avoiding contacts with defectors. But, again, this occurs at a cost in societies where “birds of a ‘cue’ flock together.” As with Schelling’s model (1971), if people switch neighbors based on homophily (McPherson, Smith-Lovin, and Cook 2001), a moderate degree of such preference at the micro level can produce a high level of segregation with very homogeneous factions at the macro level.
The current study seeks to investigate sociopsychological and structural mechanisms that may underlie the evolution of nonreciprocal cooperation in networked societies. We simulate the Prisoner’s Dilemma game dynamics on fixed networks and then on evolving networks by varying the likelihood of choosing tolerably similar partners relative to imitating higher scoring neighbors. Under the circumstances, we explore how parochial exchange in structured populations can lead to emergent regimes that remain cohesive and display high levels of cooperation, while being relatively stable against in-group deviants. And, we identify conditions that allow for such societies that, at the same time, maintain moderate degrees of cultural diversity and tolerance for diversity.
Two Mechanisms of Cooperation
The present research, initially informed by evolutionary game theory and evolutionary psychology, considers two complementary mechanisms through which cooperators preferentially interact with each other. First, “spatiality” is a critical factor. The argument for natural selection of defection is predicated on a well-mixed population, where everybody interacts equally likely and also randomly with everybody else. This approximation is used by a standard approach to evolutionary game dynamics, but real populations are not well mixed (Nowak 2006). Instead, some individuals interact more often than others; people are embedded in social networks interacting with a subset of population. In a canonical study by Nowak and May (1992), if agents locally interacting with a fixed set of partners leave offspring in the neighborhood in proportion to fitness, the likelihood of the survival of altruists (those who always help others) in the presence of defectors increases. In other words, cooperation can evolve even in the setting of one-shot interaction with the help of “network reciprocity.” If preferential partner selection (e.g., breaking ties with defectors) were available, cooperative societies would be more robust. Emergent clusters of altruists on either static or dynamic networks can benefit from interactions with their own kind while ostracizing defectors.
Second, partner identification matters. The kin selection theory (Hamilton 1964) asserts that the propensity to cooperate between two individuals is determined by how much genes they share. However, it is inappropriate for cooperation toward nonrelatives especially prominent in human societies. Krebs (1987) instead suggests that evolutionary pressure had induced humans to use various heuristic cues for “phenotype matching” as they no longer lived in small and homogenous groups such as clans, tribes, and bands. Evolutionary psychologists contend that one thrust of the evolution of human intelligence involves the detection of cheaters (Cosmides and Tooby 1992). People tend to decide whether to cooperate depending on the trustworthiness of interaction partners. Not a few studies (e.g., Nettle and Dunbar 1997; Nowak and Sigmund 1998) emphasize that generalized exchange does not necessarily rest on those who indiscriminately help one another; discriminators who conditionally help others can enhance cooperation without “spatial reciprocity,” which is otherwise impossible with undiscriminating cooperators alone. As long as a system is evolved that reliably signals trustworthiness, cooperation can be maintained even in unstructured populations where random interaction occurs.
Tag-based Action and Selection
One way in which cooperation can evolve among nonkin depends on the ability of actors to identify partners through the use of sociocultural markers, and differentially interact with others who are likely to cooperate. Human agents have “tags” (Holland 1993) as observable markers to form cultural groups; in other words, reading tags enables humans to make distinctions between in-group members and “others.” This tag-based partner identification (tagging) and its consequences are widely confirmed in social psychological experiments guided by social identity theory (Hogg 2006): people tend to distinguish “us” from “them” through categorization as a cognitive process of classifying stimuli on the basis of similarities between them (Tajfel 1974; Turner 1982); categorization based on a trivial label promotes the allocation of more rewards to in-group members in a commons dilemma (Kramer and Brewer 1984); and cooperation occurs in a simultaneous one-shot Prisoner’s Dilemma game when players expect benefits from coplayers staying the same categories as their own (Karp et al. 1993; Yamagishi and Kiyonari 2000; Kiyonari and Yamagishi 2004).
Recent research reports that cooperation in multiagent Prisoner’s Dilemma games can be significantly enhanced in tag-based systems (Riolo 1997; Riolo, Cohen, and Axelord 2001; Axelrod, Hammond, and Grafen 2004; Hales 2000, 2004; Edmonds and Hales 2005; Traulsen and Schuster 2003; Jansen and van Baalen 2006; Kim 2010a, 2010b). Tagging may play two different roles in the evolution of cooperation: affecting what actions to choose and affecting how to select partners. First, in the action-centered approach, players use tags to decide whether to cooperate toward coplayers. For example, agents cooperate only with perceived in-group members, those who share the same tags as their own (Axelrod, Hammond, and Grafen 2004; Traulsen and Schuster 2003; Jansen and van Baalen 2006) or those who have tolerable social distances (Riolo et al. 2001; Kim 2010a, 2010b). Interaction partners are randomly drawn from the population or adjacent neighbors. Second, in the selection-centered approach, it is the chance of interaction that is affected by tagging. For instance, agents move around to search the population for similar partners (Riolo 1997) or identical partners (Hales 2000, 2004; Edmonds and Hales 2005). Strategies are not based on tag-based perception in these models.
Coevolution of Cultural Traits, Cooperation, and Networks
Riolo et al. (2001) is the first model of nonreciprocal cooperation from the action-oriented approach. In their model, agents cooperate only with similar others, and tags and tolerance for diversity are inherited by the offspring. The major finding is that emergent societies reach high levels of global cooperation, but tolerance drastically decreases during the first few generations. Although more discriminating individuals are selected over generations, cooperative exchange increases as societies continue to become homogenized. The replication by Edmonds and Hales (2003) reveals that emergent societies tend to attain universal cooperation in completely homogenous populations consisting of the strongest discriminators. These discriminators with the minimum tolerance cooperate only with partners displaying exactly the same tags as their own and otherwise refuse to cooperate. Riolo et al. do not consider the invasion by unconditional defectors who look identical to parochial cooperators, resulting in the disintegration of society-wide cooperation. Noticeably, existing studies commonly report the failure of sustainable cooperation led by agents who conditionally help reliable partners on the basis of linguistic markers (Nettle and Dunbar 1997), image scores (Nowak and Sigmund 1998), or communication tokens (Miller, Butts, and Rode 2002).
The findings of the loss of diversity, the growth of intolerance, and the instability of cooperation in Riolo et al.’s (2001) model are disturbing. How is it possible that a society establishes more robust parochial cooperation without a great amount of diversity and tolerance erosion? There are several mechanisms that might lead to such outcomes (Kim 2010a, 2010b). First, very high rates of mutation in tags and tolerance (e.g., ineffective assimilation in a cultural model) could supply sufficient heterogeneity. Consequently, agents keep drawing distinctions while more tolerant individuals are continuously introduced. Second, larger societies would be more likely to maintain cooperation with diversity; selection favors defection, but it cannot overrun cooperative niches. Third, if some elements of tags are immutable, existing parochial cooperators would be less likely to accept mutant defectors as in-group members. These possible directions are not further examined here. Our focus is instead on some well-known stylized facts: individuals in many real situations are not forced to interact with the same partners and to be influenced by them, individuals often terminate the current relationships in search for others, and the average degree to which individuals change their partners is the characteristic of a society.
Models of the coevolution of action strategy and network selection in social dilemma games have indeed become a major research focus (Eguíliz et al. 2005; Santos, Pachecco, and Lenaerts 2006; Hanaki et al. 2007; Buskens, Corten, and Weesie 2008; Fu et al. 2008). In such artificial societies, adaptively learning or rational agents interact and are influenced by their neighbors, as in social dilemma games on fixed networks. In addition, agents may break ties to existing neighbors and form new ones. The interplay of influence of the network embedding on agents and rewiring of their network positions gives rise to the coevolution of agent traits including strategies in the population and the network topology.
In such coevolutionary models, the theoretical essence lies in the rules by which agents interact with, are influenced by, and break and make ties with members in a society. In evolutionary game-theoretical approaches (Weibull 1995; Schlag 1998), agents simply imitate the strategy either of the most successful neighbor or of more successful one at the previous round (influence). Lab experiments report that such imitation is commonly used by human subjects (Traulsen et al. 2010). However, there are inconsistencies between how to interact with partners (action) and how to choose them (selection); conventionally, agents use a single pure strategy, “always-cooperate” or “always-defect,” toward all neighbors, whereas they choose partners by reading image scores (Fu et al. 2008), randomly (Eguíliz et al. 2005), or based on a myopic calculation of expected benefits and costs (Hanaki et al. 2007). Further, both tie dissolution and tie construction are unilateral (e.g., Fu et al. 2008), or the former is consensual while the latter is unilateral (e.g., Eguíliz et al. 2005). Neither is realistic.
In classical game-theoretical approaches, a consistency between the action rule and the selection rule is maintained (e.g., Buskens et al. 2008); rational agents attempt to maximize their utilities while considering their partner’s strategy. When the rational approach is not available, imitation is very plausible for agents with minimal cognitive ability. Agents in the current model imitate cultural markers from more successful neighbors and modify tolerance toward their levels; since agents do not have complete knowledge about the status of network, all possible choices of strategy including tie formation and deletion cannot be considered. Tolerance is a proxy strategy, but agents do not change it for utility maximization. Also, agents do not strategically manage ties in a forward-looking manner.
Cultural tags play a role in interaction in our model; agents cooperate with tolerably similar others, but refuse to cooperate with the rest. Agents determine who is sufficiently similar or dissimilar by comparing the amount of difference with the other to a “threshold’ of tolerance. This threshold, like cultural tags, is influenced by more successful neighbors at the previous round. Tags also play a role in selection. Tolerance can be a plausible heuristic for partner change. However, it leads to an inconsistency between the action rule (i.e., tag-based similarity) and the selection rule (i.e., tolerance). Another effect is a serious bias toward cooperation because agents break ties with neighbors who are more likely to defect (and furthermore create new ones with those who are predisposed to cooperate). Our agents terminate ties unilaterally with dissimilar others and build new ones to similar others. For a new tie to be created, however, the difference between the two agents must fall within their tolerance levels; that is, new ties are formed by mutual consent. Notice that more similar (dissimilar) partners are not necessarily those who have higher (lower) propensities to cooperate because tags are independent with tolerance.
Our agents either are influenced by more successful neighbors or break a tie and form a new one, rather than doing both at the same time. In the current model, the likelihood of these alternatives is exogenous (e.g., Santos et al. 2006; Fu et al. 2008), which we call network plasticity (i.e., the relative likelihood of changing partners vs. imitating higher scoring neighbors). This network plasticity may differ from one society to another. Given a certain amount of plasticity, social networks change with varying degrees of a closure bias (Granovetter 1973; Watts 1999). Another characteristic of a society is plausibly “network closure” (i.e., the average extent to which interpersonal networks are transitive) reflecting the overall preference for homophilic interaction. It is exogenously treated in the current model, too (e.g., Hanaki et al. 2007; Fu et al. 2008). Agents may form new ties, by mutual consent, with neighbors-of-neighbors (network closure) or agents randomly recruited from the whole population.
In sum, the artificial societies we explore in the following are ones in which agents imitate cultural traits, both markers and tolerance, from more successful neighbors. Alternatively, they may unilaterally break ties with dissimilar neighbors and consensually form new ties to similar new partners selected either by referral or from the entire society. We seek to understand the effects of network plasticity and transitive closure bias in the selection of new partners on the coevolutionary dynamics of parochial cooperation and network topology. A particular interest is whether variation in network plasticity and clustering enables stable cooperative societies even in the presence of emerging defectors displaying tolerably similar markers that reduce a tendency toward decreased cultural diversity and high intolerance.
The Model
We construct a multiagent model (Wilensky 1999). After initialization, the stages of simulation per round are similarity perception, interactions, payoff calculation, and cultural evolution or network evolution, as explained in more detail in the following. Agents are selected in a random order without any particular schedules, which avoid the execution order effect. We use synchronous updating 1 ; the environment is updated for the next stage only after all agents have finished their actions at the current stage. We compare, if necessary, differences between our model and Riolo et al.’s (2001) with reference to verification issues raised by Roberts and Sherratt (2002) and Edmonds and Hales (2003).
Initialization
N(= 100) agents with randomly assigned tags and tolerance are located on an Erdös-Rényi random network where ties are binary and symmetric. Tags and tolerance are uniformly distributed. The network follows a Poisson degree distribution in which every possible edge is created independently with a fixed probability = .06. We define the neighborhood set Ni
as agents directly connected with agent i. Each agent i has a tag list of the length L in which cultural markers are binary: ti
Similarity perception
If L ≥ 1, agent i has perception of dissimilarity to neighbor j based on Hamming distance defined by
Interaction
Agents simultaneously play a one-shot Prisoner’s Dilemma game with all their neighbors. Agent i cooperates with agent j (strategy sij = 1) if the neighbor j is perceived as in-group; otherwise, agent i defects (sij = 0). Unlike Riolo et al. (2001) without unconditional defectors, agent i in our model always defects in spite of no difference between two tags as long as Ti = 0. On the other hand, agent i always cooperate if Ti = L + 1, given that the maximum tag difference is L.
If L = 0, there are no perceived social distances because all agents are identical. Since min(T) = 0 and max(T) = 1 at L = 0, one group of agents with T = 1 always cooperate, but the other group with T = 0 always defect. In this way, our model at L = 0 represents a population purely composed of unconditional cooperators and defectors (i.e., no discriminators), and generally at L ≥ 1, agents with the minimum tolerance (T = 0) are indiscriminate defectors, while agents with the maximum tolerance (T = L + 1) are indiscriminate cooperators. Agents of in-between values (T
Payoff calculation
If an agent cooperates, she pays cost c and produces benefit b. If she defects but her partner cooperates, she takes benefit b from the partner, without paying cost c. If b > c > 0, the payoff matrix in Table S1 in the replication material satisfies the Prisoner’s Dilemma game where mutual cooperation is Pareto optimal, but both rational players are worse off by choosing defection. The total payoff of agent i can be defined by
Cultural evolution or network evolution
There are a number of related and contingent processes at the last stage of our simulation, as summarized in Table S2. Each agent either updates its cultural traits toward those of a randomly chosen neighbor, if that neighbor is more successful (see the next section, “Payoff-based Imitation”). Alternatively, it breaks a tie and makes a new one if a new partner is available (see the following section, “Homophily-based Partner Selection”). Each of these processes is subject to stochastic errors.
A parameter called network plasticity p reflects the ratio of a time scale of agent’s switching interaction partners (τ
a
) to another time scale of agent’s imitating more successful neighbors (τ
e
).
2
Payoff-based imitation
To update cultural traits of agent i, agent j
Updating cultural traits is payoff-based and local in the current model, which builds upon a classical rule of strategy updating in evolutionary games on networks. Agent j
Homophily-based partner selection
Given a randomly selected agent j
Experimental Design
The analysis that follows proceeds along two steps. For both experiments, the number of agents is 100 and the average degree of initial Erdös-Rényi random networks <k> = 6. The first baseline set of experiments have agents locally interacting on fixed random networks, being influenced to change their tags and tolerance without mistakes. The focus is on the likelihood of the survival of cooperation under varying conditions of the complexity of culture (i.e., tag length L) and the payoffs. We also examine the characteristic levels of cultural diversity and tolerance at cooperative equilibrium. Additionally, we introduce 1 percent of error in payoff-based imitation to show the instability of parochial cooperation on static random networks, given the same conditions of the benefit-to-cost ratio and the tag length used in the following main experiments.
The second main experiments select an intermediate case from the baseline experiments (b/c = 4 and L = 4) for two reasons. First, cooperation can easily survive if the tag length is sufficiently high, but if it is too short, the evolution of cooperative societies is significantly disturbed (See Table S6). Next, as the benefit-to-cost ratio and the tag length increase, there is a greater probability that more tolerant agents and cultural diversity are preserved (See Table S7 and S8). b/c = 4 and L = 4 does not significantly affect this tendency. We should address that as the tag length becomes longer when holding the rate of imitation errors constant, there is less chance that in-group deviants are accidentally born (i.e., mutant defectors with similar tags that are acceptable to a dominant group of cooperators). This is why we intentionally introduce a random attack by 16 defectors at every 200th round. It is the minimum number to cover 24 possible tags, given that L = 4. All conditions of the main experiments include that a single agent commits mistakes in imitation per round while another agent does in tie-rewiring at the same round.
We are concerned with the consequences of variation in network plasticity and homophily-driven triadic closure on the demography of agents: how much cultural differentiation is there in the population? and how tolerant are agents of cultural differences? We also examine the topology of evolving networks in which agents are embedded: Under what conditions do “small-world” networks emerge? and What are the shape of evolved networks in terms of clustering, path length, and the degree distribution? Finally, we focus on the stability of parochial cooperation besides the level of cooperation: How frequently do emerging defectors lead to the collapse of cooperation? How often does defection become the dominant culture? and How is it possible that parochial cooperation is significantly more robust on evolving networks?
The Baseline Experiments
A two-factor factorial design is used. One factor is the benefit-to-cost ratio (b/c), which has the levels 2, 4, 6, and 8. The other factor is the length of agent’s tag strings (L) which takes on the values of 0, 2, 4, or 6. The level zero is particularly critical because it means that agents are not able to distinguish in-group from out-group. One hundred independent replications are performed in each condition, yielding 1,600 cases in total. Each experiment is run for 1,000 rounds or until universal cooperation or universal defection emerges, whichever comes first. According to Scheffé test (α = .05), there are no initial mean differences in the average clustering coefficients, the average path lengths, the densities, or the average degrees between any two experiment groups. Initial average levels of tolerance and of cultural diversity are dependent on L by design, but there are no significant mean differences between b/c conditions within L. The parameter setting of the baseline experiments is presented in Table S3.
The Main Experiments
The main experiments are also a two-factor factorial design. The first factor is network plasticity (p), which varies from ten to fifty in increments of ten. For each level of plasticity, the degree of closure (q) varies from zero to fifty in the same increments. We first notice that emergent societies tend to dissolve into separate communities within 2,000 rounds under experiment as network plasticity increases when controlling for closure and vice versa. Errors in partner selection (1 percent of mistakes) weaken this tendency, but to a limited extent. First, models in which changing partners is more common than changing cultural traits influenced by more successful neighbors (i.e., p > 50) commonly result in disconnected structures even at the minimal level of clustering in our experimental setting (q = 10). Even at p ≤ 50, when agents are quite likely to break ties with dissimilar others and form new ones from among the neighbors of remaining neighbors, there is a strong tendency for society to dissolve into separate communities (—s in Table S4). Within each community, local cooperation is high. The network density and the average clustering significantly decrease. Agents become more connected to neighbors displaying similar markers as their own, while the average number of emergent cultural groups (i.e., tag clusters) is relatively high. In other words, the local convergence (i.e., very high degrees of local homophily of tags) and global divergence of culture is prominent. Emergent societies can retain cultural diversity only because they are fragmented into isolated villages. Also, there become less chances of global cascades of “betrayal” triggered by emerging defectors in disconnected networks. We thus restrict our attention to lower levels of network plasticity and clustering, where the population remains connected at 1 percent of random tie-rewiring.
The final design for the main experiment does not consist of the 6 by 5 (q by p) factorial, but rather the truncated version shown in Table S4. Group(i,j) hereafter denotes the experimental group at network plasticity p = i and degree of closure q = j. In each of the 20 remaining conditions, 50 replications are performed, resulting in a total case of 1,000. Each trial is run for 2,000 rounds. All outcome measures are taken only between 1,000 rounds and 2,000 rounds, to allow the model to reach its characteristic state from the varying initial conditions. The parameter setting of the main experiments is presented in Table S5. For statistical analysis hereafter, we include four dummy variables of network plasticity (p = 10 is the reference category) and five for network closure (q = 0 is the reference).
Across 1,000 random networks in the design, the mean initial degree of agents is 5.96 (standard deviation [SD] = .323; i.e., the network density is .06, and the average clustering .06 with SD = .012). The average path length is 2.76 (SD = .074). The mean initial tolerance of agents is 2.50 (SD = .171), and their average dissimilarity to their neighbors is 2.00 (SD = .064). According to Scheffé test (α = .05), there are no initial mean differences between any two experimental groups in the endogenous variables such as the average clustering coefficient, the average path length, the density, the average degree, the average tolerance, the average dissimilarity, the proportions of each tolerance (from T = 0 through T = 5), and the average degrees of each tolerance.
Results
The Baseline Experiments
We first examine the effects of tag length and the benefit-to-cost ratio on the likelihood that cooperation survives across the replications in each experimental condition (See Table S6). We discover that unconditional cooperators—agents who cannot distinguish in-group from out-group members given L = 0—can never defeat unconditional defectors in structured populations, except under the condition that the benefit-to-cost-ratio exceeds the average degree. 5 The ability to draw even crude distinctions between “us” and “them,” however, makes cooperation evolve (L ≥ 2) under any condition in which the benefit-to-cost ratio is less than the average number of neighbors. The more fine-grained the distinction (i.e., the longer the tag length), the greater is the tendency toward cooperation.
Cooperation among parochial agents is not achieved without consequences. Table S7 shows that there is a very strong tendency for cultural homogenization, which is measured by the average Hamming distance in the population at cooperative equilibrium. Under all but the most favorable conditions for cooperation, cultural diversity is rapidly eroded by strong local assimilative influence. Emergent cooperative societies are less likely to retain cultural diversity as cooperation becomes more costly and the tag length becomes shorter.
Table S8 presents the average tolerance of agents in emergent cooperative societies at cooperative equilibrium. As societies become rapidly homogenized, agents learn intolerance very quickly from more successful others; agents adapt themselves to a continuous loss of cultural heterogeneity which otherwise serves as a guide for their discriminating actions toward others. There finally remain agents who are willing to cooperate only with very similar others and defect otherwise. The most discriminating cooperative strategy (T = 1) dominates because it is able to beat unconditional defection and is at least as good as any other strategies based on higher levels of tolerance. More than minimal tolerance can be sustained, but only when the tag length becomes longer and the benefit-to-cost ratio becomes higher.
The baseline experiments show that parochial cooperation in static random networks can emerge and prosper in a broad range of the benefit-to-cost ratios. As is illustrated in Figure 1, tolerance is uniformly distributed in the population at the initial stage (upper left). Once agents start to cooperate with tolerably similar neighbors, multiple cultural groups emerge that earn disproportionate benefits of parochial cooperation although arbitrary tags are meaningless at the beginning. During the first few generations, within-group favoritism temporarily coexists with between-group discrimination (upper right). However, as societies become more homogenized, agents displaying stronger parochialism become increasingly dominant. In other words, more tolerant agents are merged into a larger component of less tolerant agents, which leads to growing parochialism across the entire population. We notice that the overall level of global cooperation does not seriously decrease, in spite of a significant loss of the average tolerance, due to increased cultural homogeneity. A single tag cluster finally evolves, where a vast majority of discriminators are those who cooperate only with partners displaying exactly the same markers as their own (lower left).
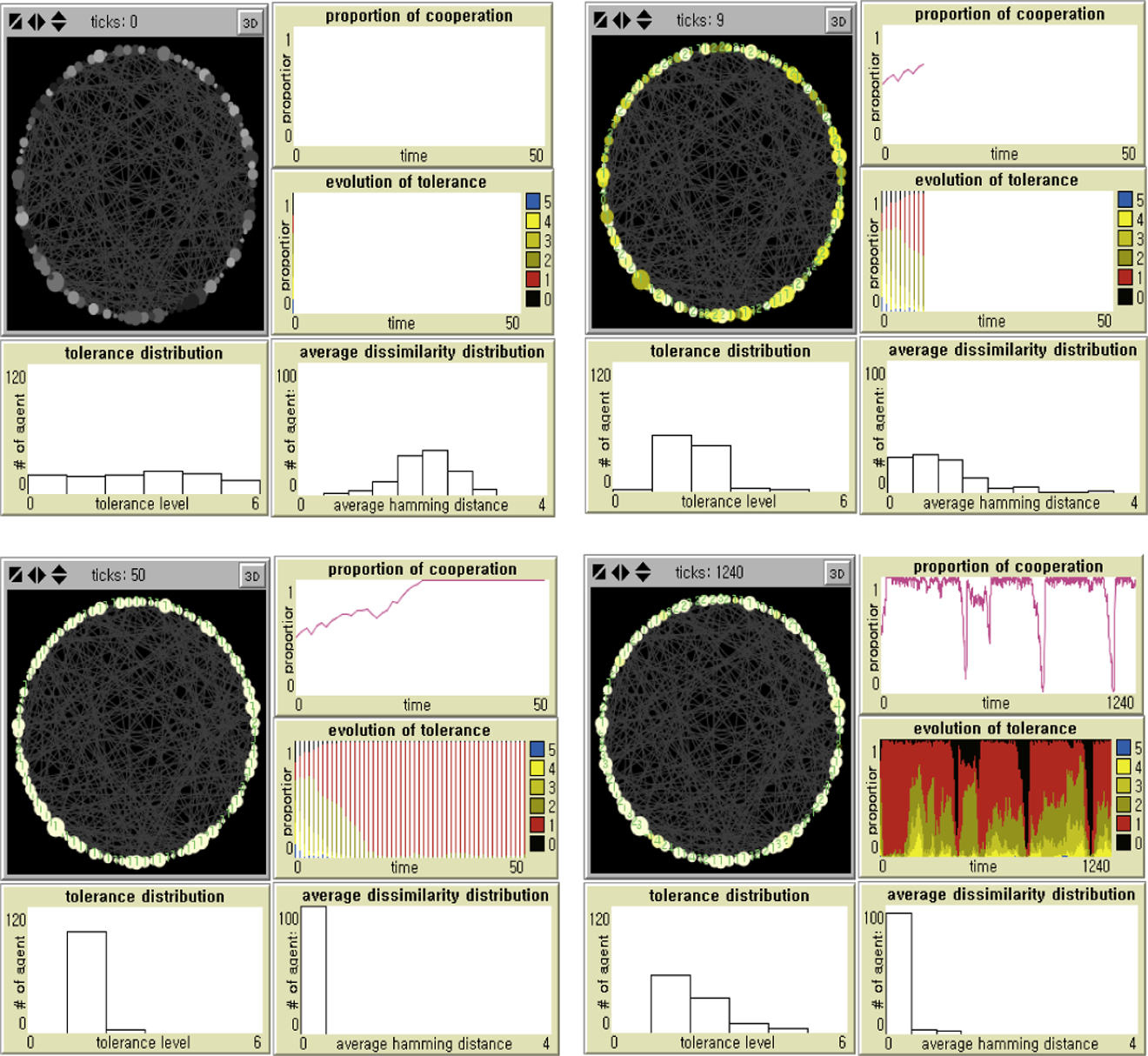
Dynamics of parochial cooperation on static random networks.
In order to examine the long-run dynamics of an emergent cooperative society, we implement 1 percent of stochastic errors in imitation after fifty rounds (lower right in Figure 1). There are several remarkable results. First, there can be a random drift toward more tolerance, indicated by an increase in the yellow areas (plot of “Evolution of Tolerance” in lower right, where “Yellows” for 2 ≤ T ≤ 4 and “Blue” for T = 5). As long as a majority of less parochial agents have the same tag strings as those of the strongest discriminators (T = 1), societies reach high levels of global cooperation, but without a considerable increase in tag diversity.
Next, even the minimum number of deviants (1 percent of the population) has big impacts on the dynamics of parochial cooperation. First, as cooperators, either parochial (Yellow) or not (Blue), are exploited by intolerant mutants (T = 0), local and global cascades of noncooperation proceed (Black). Notice that the chance of a transition to a “betrayal society” increases whenever the demography gradually changes toward the coexistence of cooperators with higher levels of tolerance. However, even the most discriminating cooperators are inevitably gullible to emerging defectors with the exactly same tags as their own. In other words, they can be dominant, but are not evolutionarily stable in the presence of defectors.
Second, a “betrayal society” is not an absorbing state in the presence of imitation errors. It is rather quickly repaired in small-scale societies when agents commit mistakes. Since defectors do not benefit from each other, once such errors create discriminators, new cultural groups evolve as they preferentially interact with one another. As the population becomes more and more homogenized, agents increasingly lose tolerance. Cultural diversity barely increases in a newly emerging cooperative society (plot of Average Dissimilarity Distribution).
The Main Experiments
We investigate the effects of network plasticity and clustering on the demography of emerging cooperative societies, network topologies, and their stability, by introducing the minimum number of defectors on a regular basis given the tag length = 4. Figure 2 shows a typical example of evolving networks with a relatively high plasticity and a moderate tendency to transitive tie-making, where sixteen emerging defectors randomly attack cooperators at every 200th round. The overall patterns contrast notably from those of the static random network in Figure 1. Across multiple replications, we observe that the diversity of culture and the average levels of agent tolerance in such networks are much greater than those in static societies. Also, we find that there is a tendency for the degree distribution to become unequal, particularly when more tolerant agents successfully accumulate social capital, as indicated by the plot of “Degree Distribution” and the average size of more tolerant nodes in Figure 2. Another valuable finding is that although the amount of mistakes in imitation is the same, plastic networks with transitive closure characteristically display much less severe cycles toward “betrayal societies.” Hereafter, we present the major results in three aspects: diversity and tolerance, network topologies, and robustness.

Dynamics of parochial cooperation on an evolving network.
Diversity and Tolerance
Homogeneity of culture
On fixed random networks cooperation can be sustained, but only with a loss of cultural diversity. Results from the main experiments show that societies with greater plasticity are more likely to retain global diversity than those in which payoff-based imitation outweighs partner selection. To the extent that agents move about, global diversity is protected against the emergence of a single dominant culture. Besides, when new partners are chosen from among neighbors of neighbors, there is an additional modest tendency toward sustaining global diversity. In Figure 3, we offer the average perceived dissimilarity in the population over the last 1,000 rounds across experimental conditions. All effects of network plasticity and the effects of clustering at q = 30 are statistically significant (α = .01), according to our linear regression analysis (adjusted R 2 = .283).

Average cultural diversity.
Tolerance
In societies where tolerance is learned from successful neighbors on random static networks, there is a strong tendency for tolerance of cultural differences to erode. In the main experiments, we find that the trend toward lower levels of tolerance is not significantly affected either by network plasticity or by clustering, according to our regression analysis. In addition, we notice that the most discriminating cooperators (T = 1) still dominate emergent societies. But, we also observe that as network plasticity increases, the proportion of unconditional cooperators (T = 5) becomes slightly high, while defectors (T = 0) are less likely to remain in the population. We present the average proportions of these three strategies on Figure S1 in the replication material.
For further analysis of the spatial configuration of parochialism and cultural groups as tag clusters, we introduce two measures of homophily at the local level 6 : the degree of link homophily in tags and the degree of link homophily in tolerance. Additionally, we calculate the number of cultural groups which share the same tags in the population. Figure 4 shows that network plasticity does not affect “birds of a ‘tolerance’ flock together.” However, the effects of network clustering are significantly positive (q = 20 and 30 at α = .01, and q = 40 and 50 at α = .05). This means that cooperators displaying similar degrees of parochialism tend to associate with each other as closure in the selection of new partners increases. Next, our regression analysis (adjusted R 2 = .31) indicates that network plasticity negatively affects the extent to which “birds of a ‘tag’ flock together.” In other words, local cultural diversity can hardly be preserved in societies where partner change is less common than imitating role models. But, we do not observe any significant effect of network closure, except q = 30 (α = .05). Finally, emergent societies tend to retain more cultural groups when networks become plastic and new partners are selected locally more than globally. Our regression analysis (adjusted R 2 = .776) reveals that both effects are significant at α = .01 except q = 50 (α = .05).
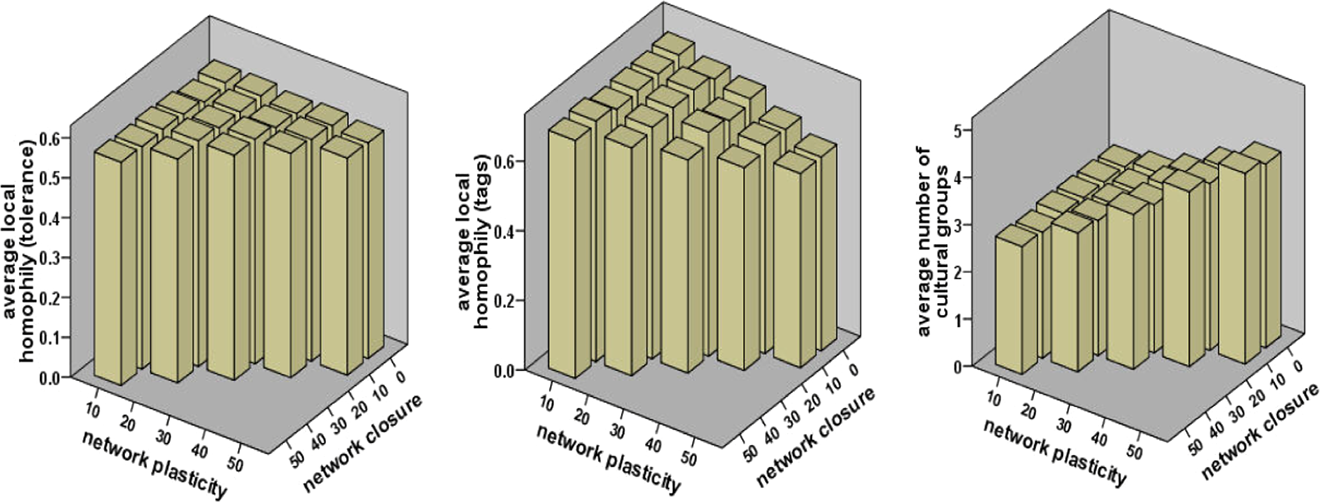
Average local homophily and average number of cultural groups.
Network Topologies
Clustering, path length, and small-world-ness
For any given level of network plasticity, the greater the tendency to select new interaction partners from among neighbors-of-neighbors, the greater the clustering of the overall network. Also unsurprisingly, as the tendency toward clustering increases, average path lengths in the whole population increase. As the joint effects of network plasticity and a bias of triadic closure continue, the density of local clusters increases at a decreasing rate, while the density of ties between clusters declines. At the extreme, high biases toward transitive partner selection result in the graph becoming disconnected with a significant diminution in network density. The realized average clustering coefficients and the average path lengths are provided in Table 1.
Average Clustering Coefficient and Average Path Length
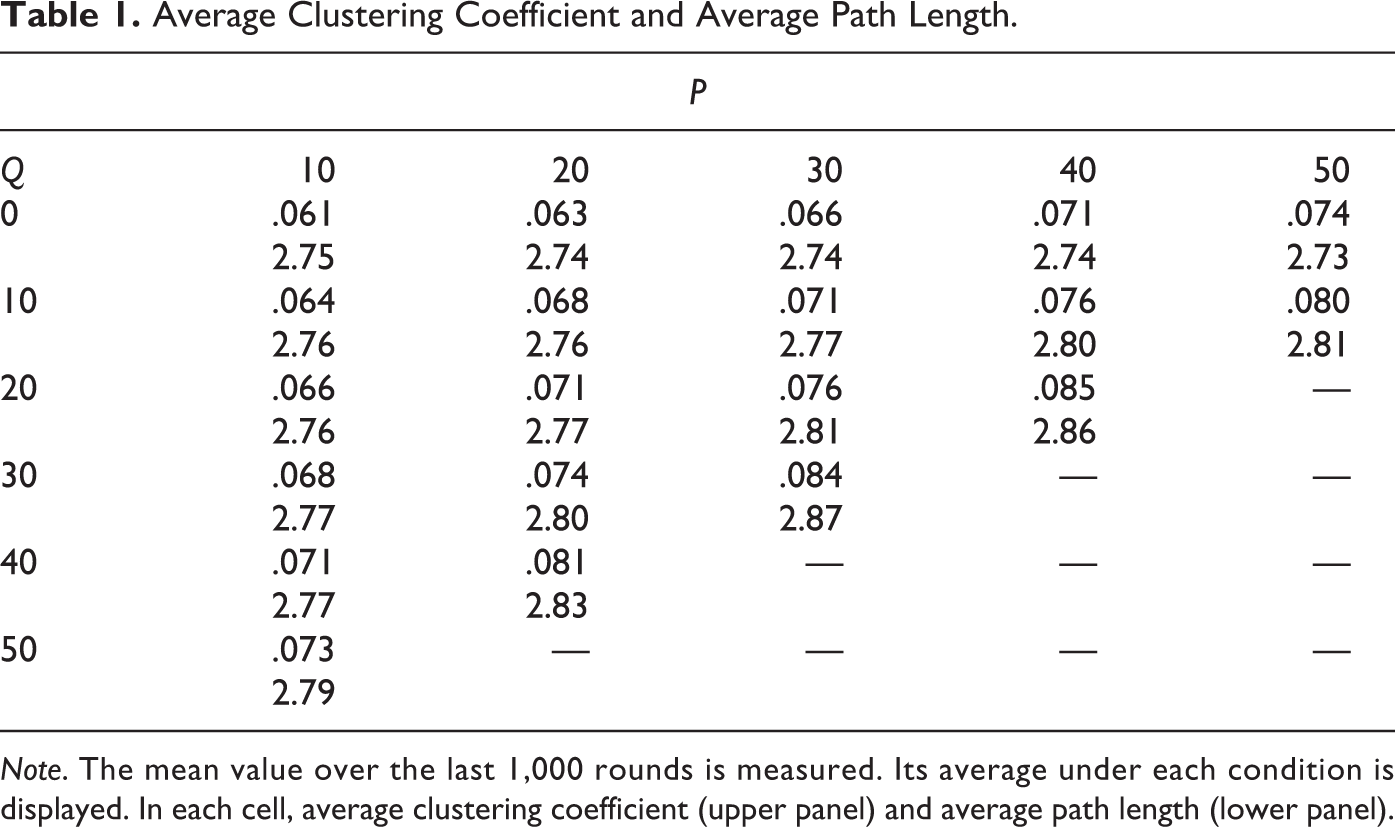
Note. The mean value over the last 1,000 rounds is measured. Its average under each condition is displayed. In each cell, average clustering coefficient (upper panel) and average path length (lower panel).
A small-world network is a graph with n vertices and average degree <k> that exhibits the average path length APL ≈ APL random (n, <k>), but the average clustering coefficient ACC >> ACC random ≈ <k> / n (Watts 1999). For each of the conditions of the main experiment, strict tests (α = .01) were performed if the mean average clustering coefficients (and the mean average path lengths) of fifty evolved networks are greater than (and differ from) the mean average clustering coefficients and (the mean average path lengths) of their original random networks at the initial stage. The former is one-tailed, whereas the latter is two-tailed. All emergent networks at network closure q ≥ 10 demonstrate similar degrees of clustering as those of random networks, whereas only some of them satisfy “small-world-ness”: G (10, 10), G (20, 10), G (30, 10), G (10, 20), G (20, 20), G (10, 30), G (10, 40), and G (10, 50). Evolved networks in all other groups cannot be classified as “small-world” because they display longer average path lengths than those of small-world networks.
Degree distribution
When network plasticity is present, there is a modest trend for evolved networks to move toward more unequal distributions of agent degree. Our regression analysis (adjusted R 2 = .160) shows that all effects are statistically significant at α = .01, except p = 20 (α = .05). The tendency toward transitive closure in partner selection does not significantly affect the maximum degree distribution. We exhibit the average maximum network degree under each experimental condition in Figure S2. Recall that agents in our model can unilaterally break old links to perceived out-group neighbors only when they can initiate new relationships with in-group partners by mutual consent. More tolerant agents are able to find new partners, but they are less likely to terminate existing ties. In contrast, less tolerant agents tend to be unsuccessful in discontinuing on-going relationships because they are less likely to meet new in-group partners. Consequently, there is a tendency for moderately tolerant agents to gain greater social capital. It is not statistically significant even at α = .05, but the most highly connected actors tend to be those who have relatively high degrees of tolerance (M = 1.824; SD = .312). Besides, there is a statistically significant tendency (α = .01) for the average degree of unconditional defectors to decline with increasing network plasticity and closure.
Robustness
Analytically, cooperative strategies (1 ≤ T ≤ L + 1) are not evolutionarily stable (see the supplementary text in the replication material). In other words, cooperative societies, once slightly disturbed, do not return immediately by selection to the original equilibrium. In our main experiments, cooperators have growing difficulties in restoring the system as random attacks are more frequently undertaken by more defectors. Nonetheless, we discover that cooperation is considerably stabilized as a network evolves over time with a relatively high degree of plasticity and a moderate degree of clustering, as illustrated in Figure 2.
We further examine how often local and global cascades of noncooperation happen in dynamic networks (Figure 5). The first one is measured by the frequency that emerging defectors reach a majority of the population after their random attacks. The occurrence of global cascades of noncooperation is measured by how frequently there is the transition from universal cooperation to universal defection (i.e., the number of the half downward cycles from 1 to 0 in the proportion of cooperation). Global cascades of noncooperation are not completely averted even in highly mobile and clustered societies, but both results in Figure 5 show that as network plasticity or/and clustering in forming new ties increases, cooperation generally becomes more stable. There are two reasons for increased stability. Breaks in the cooperative culture of a given local cluster take longer to be transmitted to other groups on evolved networks with weaker degrees of global penetration. Additionally, high clustering makes it easier for mutant discriminating cooperators to reinforce one another, which leads to a more rapid “repair” of local breaks in cooperation.
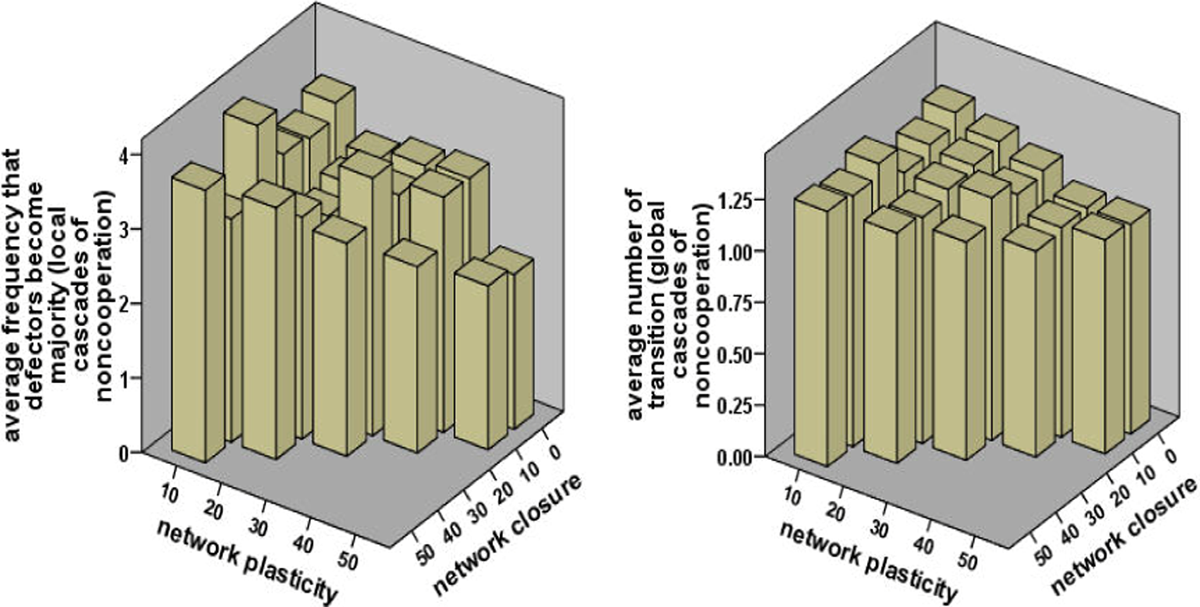
Average frequency of local and global cascades of noncooperation.
Discussion
Relatively stable and high levels of cooperation in sizable groups are commonly observed empirically, but have been a challenge to research on a multiagent one-shot Prisoner’s Dilemma game. We were informed by existing studies on two mechanisms, cultural and structural, that highlight the importance of preferential interaction among cooperators: partner identification and spatiality. First, we discovered that altruists alone can beat defectors when agents play such a game with local neighbors if the benefit-to-cost ratio of cooperation is higher relative to the number of interaction partners, and cooperation cannot survive otherwise.
Second, we manipulated the population in such a way that it consists of not only indiscriminate cooperators and defectors but also discriminators who have varying degrees of the in-group–out-group bias by reading observable tags. These discriminating egoists believe that in-group members help each other, and out-group members are predisposed to refuse to help. They may have a fear of being exploited by intolerably dissimilar partners, not necessarily out-group hostility. Similarity-based cooperation toward trustworthy partners can yield disproportionate benefits among those discriminators. This implies that parochialism may be a major requisite for the emergence of nonreciprocal cooperation and its prolongation when helping others is relatively costly in the setting of one-shot interaction.
Third, we found that cultural groups endogenously evolve with some categories, albeit arbitrary at the beginning, increasingly salient. Tolerance for diversity is, however, rapidly eroded in emerging societies due to an increasing loss of cultural heterogeneity under strong assimilation. Agents become more parochial against perceived out-group, but high levels of global cooperation are still possible among increasingly homogeneous agents. Most of time the most discriminating egoists, who are willing to cooperate only toward neighbors with exactly the same markers, form a vast majority of the population.
When agents err in imitating tags and tolerance of more successful neighbors, it leads to two consequences in the long-run dynamics of cooperation in static random networks. First, populations experience an inflow of more tolerant agents and unconditional cooperators. However, the tolerant agents do not maintain cultural diversity in static societies as they quickly copy tag strings from the strongest discriminators. Second, they are very susceptible to defectors since they are more likely to be tolerant of culturally heterogeneous others. Even the strongest discriminators are inherently vulnerable to indistinguishable defectors. For these reasons, it is inevitable that cooperative societies degenerate into betrayal ones. However, as parochial mutants are clustered to help each other, they can establish a new cooperative society. There are endless oscillations between global cooperation and global betrayal in tag-based societies.
In the main experiments, we investigated the dynamics of cooperation in tag-based societies, where agents not only take discriminating actions toward neighbors, but they also change their interaction partners to socialize with the members of one’s group. They can drop ties to dissimilar neighbors and make new ties to similar others with a closure bias, instead of learning from more successful neighbors. We found that, if social mobility and clustering are too high, societies devolve into cohesive yet disconnected subpopulations. If social mobility and clustering are too low, there are no significant differences between static societies and partly mobile societies: parochial cooperation is propelled mostly by the strongest discriminators; a highly homogeneous population is needed for globalized cooperation; and cooperation is unstable against mutant defectors. There is, however, a part of the parameter space between these two outcomes. It is on highly plastic networks evolving with fairly low global penetration that the long-run dynamics of cooperation manifest significantly different results. Here, cooperation is more stable in the presence of random attacks by emerging defectors. Emergent societies also retain higher tolerance and more cultural diversity.
How is this possible despite the fact that parochial cooperators are inherently gullible to defectors displaying tolerably similar markers in societies with completely mimicable tags? First, structural conditions lessen the likelihood of a successful invasion by emerging defectors; network clustering and reduced global penetration minimize the disruptive effects of an apparent “in-group” member betraying others. Defectors can more easily attack existing cooperative societies with increased tolerance, but local clusters of parochial cooperators reemerge to inhibit cascades of noncooperation. Defectors are less likely to become a majority because it is more difficult for them to traverse networks with long path lengths. On the other hand, more cosmopolitan agents have the potential to construct more inclusive ties in the course of partner selection with random tie-rewiring. Evolving networks become characterized by a moderate inequality in social capital as less parochial agents build outward social networks more easily than intolerant agents. Consequently, it frequently happens that newly emerging tolerant agents of different tags contribute to different cultural groups, instead of being absorbed into highly homogenized cooperative clusters of the strongest discriminators. To the extent that all these processes reinforce each other immediately after invasion by emerging defectors, a new cooperative society can emerge with increased tolerance and more cultural diversity.
Conclusion
We have sought to determine whether it is possible to create an artificial society that optimizes a number of properties: society-wide cooperation; the dynamic stability of cooperation against defection; cultural diversity; tolerance; and connectedness. The answer is both encouraging and discouraging. On the positive side, we have shown that such an artificial society is plausible—simultaneously achieving those goals is not a logical impossibility. Relatively robust, widespread cooperation among diverse and tolerant agents is theoretically possible even without either direct or indirect reciprocity. Furthermore, this desirable outcome is buttressed by a particular network topology with moderate clustering and long path lengths (i.e., it is not a small world), where there is a positive correlation between agent degree and tolerance.
On the negative side, our results suggest that the range of key structural conditions that sustain such desirable cooperative regimes may be rather narrow. Too much or too little propensity to select new interaction partners parochially results in suboptimal outcomes. More importantly, too much, or too little freedom to make and break ties with interaction partners leads to suboptimal outcomes. Fragmentation of the population into locally cooperating but nonsynchronized clusters is a common outcome in our simulated societies. Widespread cooperation can be achieved with some ease if societies are willing to sacrifice multicultures and tolerance for diversity. More cosmopolitan agents can significantly remain in evolving clustered networks of overlapping cultural groups under the optimal regime, but they are not highly persistent even within the solution space of parameters.
Cooperation in dyadic situations itself is a social dilemma, but cooperation in heterogeneous subpopulations faces yet another dilemma. People live most of their lives in fairly dense local networks of similar others, with fewer and weaker ties to members of other groups. Stable cooperation in homogenous and isolated populations may be relatively easy to achieve, but at the cost of cultural diversity at the local level. Cultural differentiation at the global level can be maintained in spite of local influence and homophily, but society-wide cooperation is problematic in terms of scalability. On the other hand, the creation of such cooperation across local clusters may be possible if there is a strong tendency toward homogenization in connected populations, but multiculturalism is more hardly preservable in relatively small-scale societies.
Despite the apparent limitations on widespread cooperation in diverse societies, such patterns are empirically common even without either a central authority or reciprocation. Also, it is not perfectly straightforward to distinguish cooperators from defectors. People tend to project own behavior onto others, or they tend to read various forms of signals of trustworthiness, as in our study. Ostracizing defectors is commonly highlighted as a key mechanism for cooperation, but it is not always beneficial to the society as a whole because it tends to obstruct large-scale cooperation. We instead commonly observe that where widespread cooperation emerges, it is very often susceptible to cascades of betrayal, punishment, and mistrust in the mere presence of deviants. Finally, cultural homogenization is not necessarily imposed. It is rather that interpersonal networks at the local level are not completely homogeneous.
Our computational research has intended to demonstrate that a regime of interaction with desirable properties—widespread cooperation, robustness against defection, moderate tolerance, and cultural diversity—is, in principle, achievable in tag-based networked societies of in-group favoritism and homophily. Such regimes exist in spite of high peer pressure on learning intolerance in the local environment under conditions balancing the ease of tie breaking and making and social influence resulting in increasing homogenization of culture. The regimes that exhibit these properties display structural characteristics of a moderate inequality of centrality of actors in terms of social capital measured by the number of ties and also a positive association between social capital and tolerance for diversity.
The main thrust of our research is the computational investigation of structural mechanisms leading to various modes of cooperation in networked societies. Sociopsychological processes in the proposed model such as us-and-them distinction (Tajfel 1974), in-group favoritism (Kramer and Brewer 1984), and homophily (McPherson, Smith-Lovin, and Cook 2001) are nearly universal in human societies. Additionally, we plausibly assume that actors locally interact with a subset of the population (e.g., neighbors, friends, and colleagues) and that they imitate more successful interaction partners (e.g., role models) when rationality fails to work. The main structural parameters in the model are empirically relevant. Network plasticity represents the extent to which relationships can be dissolved and formed relative to social learning. Network clustering reflects the degree in which interpersonal relationships are transitive. They may vary from one society to another. For instance, such relational mobility (or network closure) tends to be lower (or higher) in a collectivistic society than in an individualistic society (Schug et al. 2009; Igarashi et al. 2008).
The current study suggests three modes of cooperation. When changing tolerance and social markers outweighed network plasticity with low clustering, a society would emerge characterized by highly strong parochialism with severe losses of cultural diversity. This type of society would tend to be significantly vulnerable to tolerably similar defectors. When network plasticity overrode social learning with high clustering, a society would fragment while losing cohesion, as in Schelling’s model. Between, there would evolve a more tolerant society with diversity where global cooperation is relatively stable against those in-group deviants. The present research implies that structural parameters as such may be fundamental for a better understanding of cross-societal variations in the scalability and stability of cooperation in regimes of tolerance and cultural diversity.
Footnotes
Acknowledgment
We are grateful to Vincent Buskens, Rick Riolo, Yoshimichi Sato, Hiroshi Hamada, Yutaka Nakai, and anonymous reviewers for valuable comments on earlier drafts of this article. They were presented at the 21st Group Processes Conference, August 7, 2009 (San Francisco) and at the 50th Japanese Association of Mathematical Sociology Conference, September 10, 2010 (Saitama).
Authors’ Note
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The author Jae-Woo Kim benefited from the 2009 fellowship provided by the Utrecht University in the Netherlands.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.