
Abstract
Geographic factors such as rugged terrain and distance from capital cities are widely believed to prolong civil wars by enabling rebel groups to resist total defeat. This article argues that prevalence of malaria can similarly serve to asymmetrically enhance rebels’ defensive capabilities and thus prolong civil war. Malaria prevalence does so in three complementary ways. First, while malaria can inflict costs on both government and rebel troops, these costs are magnified for larger and denser human groups; thereby ensuring that the costs of malaria will often be higher among government troop deployments. Second, because government soldiers are rotated in and out of conflict zones whereas insurgents typically are not, the former are likely to have a higher nonimmune exposure rate than the latter, which further ensures that government forces will be more susceptible to contracting and spreading malaria. Third, malaria can also indirectly prolong civil war by helping to maintain a socio-geographic environment that is conducive to insurgency. These three complementary factors advantage rebel forces’ abilities to resist defeat by government forces and prolong civil conflicts. I empirically test these arguments by examining the duration of civil wars and find strong support for a prolonging effect of malaria on civil conflict.
[F]or many centuries, the course of military campaigns was determined as much by the presence of malaria as by military strength.
As the previously mentioned quote suggests, malaria has long played a central role in conflict processes. Yet, we lack systematic theory for how malaria affects conflict and civil war, in particular. Indeed, aside from a single recent argument for the conflict-inducing effects of malaria (Letendre, Fincher, and Thornhill 2010)—which was called into question in a subsequent critique (Hendrix and Gleditsch 2012)—the comparative politics and international relations literatures on civil war have generally ignored malaria as a key independent variable. Alongside this scarcity in malaria-oriented conflict research, civil war scholars have nevertheless identified a wide variety of other geographic factors that affect civil war, including forest cover, mountainous terrain, physical distance, access to natural resources, and proximity to international borders (e.g., Fearon and Laitin 2003; Buhaug, Gates, and Lujala 2009). Hence, the relative absence of civil conflict research on malaria is striking, especially when one considers the strong correspondence between civil war persistence and malaria prevalence in locations such as sub-Saharan Africa, India, Myanmar, Central America, and Colombia, as well as the common observation that throughout our history, malaria has killed more soldiers than has actual warfare (Ashall 1994; Ayala 2007, 175). This article therefore re-asks the question, What are the effects of malaria on civil war?
To answer this question, I develop a theoretical argument to suggest that malaria prevalence can increase the duration of civil war. To this end, the theory discussed subsequently does not suggest that malaria has any direct effect on civil war onset, but rather, it suggests that malaria will often ensure that those civil wars that have begun will be more protracted and more difficult to end. In developing the logic to this argument, I first draw upon extant civil conflict research regarding how various geographic factors (e.g., rough terrain) enhance rebel groups’ defensive capabilities and therefore their abilities to resist total military defeat. As argued subsequently, these dynamics affect the duration of civil war but do little to enhance the abilities of rebel groups to win such conflicts. By pairing this logic with existing ecological theories of malaria’s interactive effects on human populations, I then identify a number of complementary pathways by which malaria is likely to enhance rebel defensive capabilities, and thus prolong civil war.
In particular, I argue that malaria prevalence—defined here as the pre-conflict level of malaria exposure within a given geographic area—prolongs civil war through three complementary channels. First, while malaria can inflict costs on both government and rebel forces, the literature reviewed subsequently suggests that these costs will often be magnified for larger and denser groups. Accordingly, the costs of malaria—and the likelihood of an epidemic—will be higher for (what are relatively larger in size and less diffuse) government troop deployments than for rebel groups—thereby converting any government advantage in offensive power to a disadvantage. Second, because government troops are rotated in and out of conflict zones whereas insurgents typically are not, the former are likely to have a higher nonimmune exposure rate than are the latter, which further ensures that government soldiers will be more susceptible to contracting and spreading malaria. Third, high malaria prevalence also prevents a government from successfully constructing infrastructure, providing social services, and establishing a physical presence within a conflict region. As such, malaria can indirectly prolong civil war by contributing to a low state capacity environment in conflict zones that is conducive to insurgency.
I focus these arguments on malaria, rather than on other infectious diseases, for several interrelated reasons. Of note, malaria’s unique global preeminence, qualities of persistence, and general transcendence of poverty as an overriding determinant make this disease unequalled in its relative impacts on, and exogeneity to, human activities such as development and conflict. As Gallup and Sachs (2001, 87) argue in this regard, “[u]nlike other important diseases in poor countries caused by deficient living conditions, such as diarrhea, tuberculosis, and schistosomiasis, malaria is not primarily a consequence of poverty; its extent and severity are largely determined by climate and ecology.” Hence, assessments of the effects of disease prevalence on civil war duration will be most precise, and salient, when assessed with respect to malaria, as opposed to other diseases. This is especially true here, given that a number of the causal mechanisms posited subsequently are contingent upon mosquitos as primary disease transmitters. 1 This article therefore empirically evaluates the aforementioned malaria-based claims by examining the duration of all civil wars, 1945 to 2004, with the aid of a series of Cox proportional hazards models. In support of my arguments, these tests reveal that an area’s pre-conflict levels of geographic malaria prevalence have a significant, and robust, prolonging effect on civil war duration, particularly among civil wars with rebel actors that are verifiably weaker than opposing government forces. What is more, these effects are also substantively sizable and are comparable in magnitude to the estimated effects of other well-known determinants of civil war duration, such as rebel groups’ degrees of territorial control, rebel group strength, and democracy.
Through these findings, this study makes several important contributions to the literatures on political conflict and political–economic development. Past research has failed to fully explain what effects malaria has had on civil conflict, if any. As mentioned previously, this deficiency is striking, given the strong correspondence between civil war persistence and malaria prevalence during the post–World War II era. The current project addresses this deficiency by providing a systematic theory for the prolonging effects of geographic malaria prevalence on contemporary rebellion and civil war. These theoretical linkages, in turn, may allow future scholars to better explain why in many instances superior offensive capabilities do not translate to (timely) military victory, such as was the case in the United States–Vietnam war, and during the more recent decades of the Fuerzas Armadas Revolucionarias de Colombia (FARC) insurgency in Colombia. Second, while past social science research (e.g., Sachs and Malaney 2002) has largely characterized the social and political–economic burdens of malaria as additive—and uniformly increasing in costs at the country level—the theory presented here offers a more nuanced relationship, in that, it portrays the burden of malaria to be interactive, and most severe for sparsely populated areas (and the social or political activities therein) that see large, but pre-urbanized, increases in human populations and population densities. As discussed subsequently, this insight has implications for our understandings of malaria’s effects on economic development more generally, and on state capacity, natural resource extraction, electoral turnout, crime, and social cohesion more specifically.
The remainder of this article proceeds in three parts. In the following section, I draw upon the literature pertaining to (1) civil war and (2) malaria ecology in order to identify the causal mechanisms that potentially lead malaria to enhance the defensive capabilities of rebel groups and to prolong civil war, leading me to derive two testable hypotheses. The analysis section then begins by describing the data, estimation model, variables, and research design that I use to test these hypotheses. Following this description, the core of my analysis section presents several empirical tests of my hypotheses, interprets the substantive significance of my findings, and subjects these findings to a battery of robustness tests. I conclude with a discussion of my results and their implications.
Theory
While civil wars frequently vary in their underlying strategic dynamics (de Rouen and Sobek 2004; Fearon 2004), a majority of the world’s contemporary rebellions have been fought as insurgencies. There is a clear logic as to why this is so. Strategies of insurgency—defined here as militarized civil conflicts involving small groups undertaking guerilla warfare in rural settings (Fearon and Laitin 2003, 75)—offer a number of distinct tactical advantages to rebel forces and are thus the preferred strategy of violence for groups seeking to rebel against more powerful states. In particular, insurgencies allow rebels to persist in the face of larger and better armed government forces, essentially by enhancing rebel groups’ abilities to resist or evade government repression within their areas of operation (Cunningham, Gleditsch, and Salehyan 2009, 575). As argued subsequently, these defensive advantages arise through a combination of (1) rebel groups’ typically sparse, clandestine, and sporadic tactics of guerilla warfare and (2) the socio-geographic features of conflict areas. Together, these features level the playing field; making it more difficult for government forces to successfully utilize their superior offensive strengths and thereby ensuring that relatively weaker, and smaller, rebel groups can persist even in the face of significant power asymmetries.
Several important factors have been identified as uniquely advantaging rebel forces along these lines. Rough terrains, most notably mountains and jungles, have long been characterized as offering asymmetrical defensive advantages to small or weak rebel groups (Fearon and Laitin 2003; Buhaug, Gates and Lujala 2009, 547). As Buhaug, Gates, and Lujala (2009, 551) succinctly note in this regard, “[r]ugged terrain…favors small, irregular bands by making them harder to detect and defeat by government forces. A group that can retreat to areas where it is protected from the enemy can more easily regroup, rearm, and train, and is therefore able to continue fighting for prolonged periods.” With this same logic, rebel groups’ control over territory, the distance of a conflict region from a government’s capital, and rebel access to porous international borders, have each similarly been shown to geographically advantage rebels’ defensive capabilities in the face of superior government forces (Buhaug, Gates, and Lujala 2009; Cunningham, Gleditsch, and Salehyan 2009). As a result, civil wars exhibiting these latter features, often described as peripheral insurgencies, have proved exceptionally difficult to end (Fearon 2004, 277). Finally, social attributes can also at times enhance the defensive capabilities of rebel groups in comparable manners. For instance, as Fearon and Laitin (2003, 76) note, larger populations, and better rebel knowledge of local populations, can each improve rebel groups’ abilities to evade or resist repression by adversarial government forces.
Each of the previously mentioned factors erodes a government’s superior offensive capabilities by asymmetrically lowering government troops’ abilities to inflict costs upon (typically weaker) rebel groups. Critically, however, the defensive advantages provided to insurgents by these factors will not necessarily translate into higher probabilities of rebel victory, as they do not directly enhance rebel groups’ abilities to increase their own offensive strengths or to defeat the still better equipped government forces on the government’s own territory. Rather, asymmetric defensive advantages—such as rugged terrain—merely help to ensure that rebel groups cannot themselves experience total defeat (Buhaug, Gates, and Lujala 2009; Cunningham, Gleditsch, and Salehyan 2009). This distinction is critical, as it suggests that the factors discussed previously will not alone determine a civil war’s ultimate outcome or winner, but rather, will predominately affect war duration. Better put, “unlike high offensive strength, the ability to resist is far less likely to translate into success at the negotiating table or concessions from a government. As such, we may see long conflicts where rebels are too weak to extract concessions or obtain negotiated settlements, yet too secure to be easily eradicated by governments” (Cunningham, Gleditsch, and Salehyan 2009, 575). Thus, defensive factors that favor small rebel groups’ pursuit of guerilla tactics will prolong civil wars but will do little to ensure one side’s victory over the other.
Drawing upon this logic, I argue subsequently that high malaria prevalence (within a conflict region) will similarly serve to asymmetrically enhance rebel groups’ defensive capabilities and thus prolong the duration of civil war. Stated briefly, malaria prevalence does so in three complementary ways. First, while malaria can inflict costs on both government and rebel groups, these costs are often multiplied for larger and denser groups. Hence, the costs of malaria—and the likelihood of an epidemic—will be higher for (what are typically larger in size and less diffuse) government troop deployments. Second, because government troops are rotated in and out of conflict zones whereas insurgents typically are not, the former are likely to have a higher nonimmune exposure rate than the latter, which further ensures that government soldiers will be more susceptible to contracting and spreading malaria. Third, high malaria prevalence also prevents a government from successfully constructing infrastructure, providing services, and establishing a political presence within a conflict region. As such, malaria can indirectly prolong civil war by helping to maintain a socio-geographic environment that is conducive to insurgency. Taken together, these three complementary factors thereby advantage rebel groups’ abilities to (1) resist defeat by government forces and (2) prolong civil conflict.
The first factor mentioned previously—which posits that larger and denser social groups will be more susceptible to malaria—is rooted in the literature on “frontier malaria.” This literature argues that the costs of malaria follow an inverted “U” shape with respect to human settlement in malarial regions: one first observes a relatively low malaria rate in areas with sparse, widely dispersed human populations (phase 1), then a spike in malaria rates during the initial stage of organized settlement (phase 2), and finally a decline in malaria rates as an area becomes increasingly urbanized (phase 3). 2 There are two reasons for the malaria spike in phase 2. First, relative to very sparse human populations, denser and larger populations of individuals—such as those accompanying organized mining, logging, or farming communities—directly increase the potential human transmission points for malaria by adding to the “tropical aggregation of labor” (Martens and Hall 2000, 105). Observing this process at work in the Congo, for example, Trape and Zoulani (1987, 23) note that “each human settlement initially favours the multiplication of breeding places and the presence of high density populations of A. gambiae” mosquitos, whereas Singer and de Castro (2001, 203) similarly observe among Amazonian frontier settlements that the “combination of high human density, high vector density, and lack of immunity was destined to produce a malaria outbreak.”
Second, and again relative to the sparse and diffuse human settlements found in phase 1, these denser (phase 2) human settlements are also necessarily accompanied by larger proximate sources of standing water (for water collection or irrigation), more deforestation, and higher rates of animal displacement (the mosquito’s alternate food source)—each of which further increases human exposure to mosquitos (Sawyer 1993; Martens and Hall 2000, 105). Hence, the initial disturbance of the natural environment by human settlement facilitates conditions that encourage more breeding of mosquitos, and more targeting of human hosts. These high malaria rates then typically persist within an area until a settlement becomes sufficiently urbanized (phase 3), wherein malaria declines alongside an urbanized settlement’s increased areas of cleared land, elimination of major sources of standing water, improvements in housing, better knowledge of the disease and access to health care, and higher rates of resistance (Barbieri, Sawyer, and Soares-Filho 2005, 851)—a process that can take 10 to 20 years to complete (Sawyer 1993, 1132).
Regarding government and rebel forces during civil conflicts, I argue that the dispersed, guerilla fighting styles of insurgent groups will help to situate these rebel actors within a phase 1 environment of low malaria exposure, whereas the typically larger, and more dense military structures of government forces instead align this group much more closely with the high malaria rates found within phase 2. Indeed, by virtue of being smaller in absolute number, and pursuing more diffuse styles of combat and habitation, insurgents are less likely to be deployed and stationed in semipermanent camps of medium to large groups, whereas government soldiers will often establish large-scale military bases and camps of these sorts when pursuing activities in conflict regions. 3 These latter practices accordingly help to ensure that government troops will be more prone to transmitting malaria between one another both directly, through their larger group size and density, and indirectly, through increased proximate standing water resources, semipermanent habitat construction, clear-cutting, and animal displacement—all of which foster conditions for higher vector contact and hence malaria transmission. Thus, taken together, traditional government military forces will asymmetrically bear the burden of conflict-zone malaria during times of war.
Historical accounts suggest that these factors do indeed severely harm and limit traditional military forces in malarial regions, especially as the size and density of a military force expands. Beadle and Hoffman (1993, 320) report, for example, that when the size of US naval forces in Vietnam during the Vietnam War was “increased by about 50% from fiscal year 1966 to fiscal year 1969, the incidence of malaria increased almost eightfold” (1993, 320), 4 whereas when the number of US naval forces then decreased dramatically during 1969–1970, the “malarial incidence rate among the personnel still in Vietnam decreased as well” (p. 321). Earlier US military occupations of Cuba and Haiti experienced similar, if not more severe, casualty rates as the result of malaria prevalence in conflict areas (Allen 1923; McCullough 1977, 406). Similarly, while observers at the time noted that Panama’s 1903 revolt against Colombia could have likely been quelled by a mere 600 Colombian troops, “a Colombian force of some two thousand men did attempt an overland march through the Darien wilderness, but ravaged by fever, they gave up and turned back” (McCullough 1977, 379), and with a US blockade in Panama’s Caribbean port, the Panamanian revolution was successful. Additional evidence also suggests that modern rebel groups recognize the costs imposed upon government forces by malaria. For example, as a Karen National Union spokesman noted with respect to his group’s rebellion against the Myanmar military junta (State Law and Order Restoration Council [SLORC]) in 1997, “[t]his will be a decisive engagement, either SLORC will triumph or they will be dragged down by the cost of the war, by heavy casualties and by growing disaffection within their army. And because they are operating in an endemic malaria area, they stand to lose as many men from sickness as from fighting.” 5 In full, these anecdotes accordingly suggest that state-based military forces have historically, and disproportionately, incurred high costs as a result of malaria prevalence.
Moreover, it is unlikely that these disadvantages will be offset by superior access to medicine among government forces. Most civil wars are fought in the developing world and in such contexts, government troops, like rebel groups, are often significantly underfunded in health and human services (Elbe 2002; Henk and Rupitya 2001, 18). For instance, in assessing the Filipino government’s counterinsurgent troops in 1986, Filipino president Corazon Aquino “noted that inadequate rations force government troops to steal from civilians and that heavy casualties from pneumonia and malaria, as well as combat deaths, could be traced to a lack of clothing and an almost complete absence of field doctors and medicine.” 6 Even among those government troops that are well funded, successful use of malaria abatement technologies has proved to be near-impossible for soldiers in the field. Joy (1999) observes for US troops in the Pacific during World War II, for example, that “environmental control could not be performed in forward combat areas (including the use of bed nets). Even with clothing discipline, hands and faces were exposed, and repellent, even if used, was rapidly removed in the sweat” (p. 203), whereas in more recent US conflicts, “Gambel et al. (1998) reviewed the use of personal preventive measures, to include application of topical insect repellents and permethrin treatment of clothing and netting, among deployed personnel. Their study demonstrated that deployed soldiers do not understand the use of these measures and as a result fail to use them properly, if at all” (Kotwal et al. 2005, 215). Furthermore, as argued subsequently, any natural immunity to malaria gained by government troops operating in malarial regions will be rapidly undone (especially relative to rebel groups) as a result of governments’ practices of frequent troop rotation in to and out of conflict zones. Hence, it is improbable that government troops will hold a superior medical advantage with respect to malaria—which has no known vaccine—in civil war settings.
As alluded to previously, a second socially disruptive feature of malaria that may also asymmetrically burden government troops during times of civil war concerns the relationship between military troop rotations and malaria immunity. Troop movements and rotations have been argued to exacerbate disease epidemics throughout history (Smallman-Raynor and Cliff 2004) as well as in more recent civil war torn regions such as Sierra Leone (Bazergan 2002). Regarding malaria immunity specifically, extant findings suggest that whereas individuals living in malarial regions build up immunity over time, newcomers to a malarial region typically have low-level immunity or are nonimmune, which in turn increases newcomers’ likelihoods of both catching malaria and spreading it (Martens and Hall 2000). Accordingly, research has found that “outsiders” face higher malaria burdens when visiting a malaria region than do local inhabitants, which is a reality that has historically limited intergroup contact, inhibited the transmissions of ideas, and prevented movements of labor (Sachs and Malaney 2002; Letendre, Fincher, and Thornhill 2010). Furthermore, because local acquired immunity often dissipates over the course of a year or so, local inhabitants that have acquired immunity but leave a malarial region for schooling or work will face the same burdens as migrants when they return home (Sachs and Malaney 2002, 684). In sum then, the presence of malaria in a given region imposes asymmetric costs upon individuals and groups, which counterintuitively are highest for those individuals or groups that have had the least amount of constant exposure to that region.
The newcomer characteristics described previoulsy closely resemble the troop deployment practices that governments undertake, to a disproportionate degree, while fighting civil wars. Civil conflicts are typically fought far from a government’s command center and resource base, within the regions of a country or territory that insurgents actively reside in (Buhaug, Gates, and Lujala 2009). As the civil conflict literature reviewed earlier suggests, these features help rebels to evade capture, and thus, enhance insurgents’ defensive capabilities. Accordingly, leaving conflict regions can be costly for rebel actors, as doing so often entails travel to areas that (1) are under more direct government control and (2) exhibit fewer opportunities for rebels to blend in or receive aid from sympathizers. Thus, the dynamics of civil war often necessitate that rebels occupy (malarial) conflict zones full-time, which in turn ensures that rebel actors will accordingly build a degree of immunity to malaria over time. By contrast, government soldiers generally live and train far from a civil war’s active conflict zones and are only deployed to conflict zones for military activity, thereby ensuring a higher malarial nonimmune rate among government soldiers. In addition, government troops, unlike rebels, are commonly provided with paid leave periods, or are otherwise rotated in and out of conflict regions over time (e.g., for medical treatment). This latter feature further guarantees that even if some government troops do acquire malaria resistance, the playing field will not remain level for long. Taken together, government soldiers will frequently experience higher malaria burdens due to nonimmunity than will rebel forces, even as the latter exhibit more persistent exposure to a malarial region. These phenomena were observed succinctly in one recent study of an Indian Air Force outpost within a highly malarial region, which noted that consequent heavy transmission of falciparum malaria among the tribals perennially contributes to the local tribal developing some immunity, leading to a stable state of malaria in them (Manson-Bahr 1982). This makes the nomadic service population who come in contact with such tribal areas within flight range of vector mosquitos highly vulnerable because of their non-immune status. These can raise additional problems in that the very presence of such non-immunes may convert an endemic stable malaria into an epidemic. (Banerjee 2001, 290)
Hence, malaria has the potential to asymmetrically burden government troop deployments within a conflict region. To the extent that this increases rebel groups’ abilities to resist a total military defeat, malaria will prolong civil war.
Third, malaria also enhances rebels’ defensive capabilities by preventing governments from successfully constructing infrastructure, providing services, and establishing a broader physical presence within a conflict region. Insurgents gain defensive advantages in civil wars when they possess, among other factors, deeper knowledge of local populations; favorable recruitment pools arising from poor socio-geographic conditions; and rugged terrains that preclude government transportation, occupation, or communication (Fearon and Laitin 2003; Buhaug, Gates, and Lujala 2009; Cunningham, Gleditsch, and Salehyan 2009). By inhibiting governments’ abilities to (1) construct transportation and communication infrastructure, (2) provide public services, and (3) maintain a political and bureaucratic presence in an area, malaria helps to ensure that those socio-geographic features that are favorable to insurgency, when already in place, will remain so. Herein, research suggests that frontier malarial regions, even when devoid of insurgency, will be dominated by political marginalization, unplanned development, agricultural failures, weak institutions, and low community cohesion (Singer and de Castro 2001; de Castro et al. 2006, 2452). At the same time, major public works projects and government attempts to build transportation infrastructure in malarial regions will often also be severely impeded or will fail altogether, which can further exacerbate existing malaria exposure levels (McCullough 1977; Martens and Hall 2000). In effect, these dynamics thus help to erode, or to maintain low existing levels of, state capacity within conflict regions, which itself has been shown to be related to civil war (Hendrix 2011). Consequently, malaria prevalence will maintain a socio-geographic setting that is conducive to insurgency, to rebel recruitment, and to rebel defensive capabilities more generally. As such, malaria prevalence can indirectly foster long, drawn-out civil wars, essentially by ensuring that rebels have access to the very resources that past scholarship suggests will allow them to resist total defeat.
In sum, there is good reason to suspect that, when malaria is prevalent in a conflict region, the factors discussed previously will combine to erode the offensive advantages traditionally held by governments during civil wars. By this same token, many of these malarial factors will also facilitate increased defensive capabilities among rebel groups. Specifically, the combination of high human density, low immunity, and high malaria prevalence can create a “perfect storm” for malaria epidemics among government forces—a phenomenon that is much less likely to arise within smaller, more diffuse, and relatively more immune rebel groups. Because past scholarship has consistently established a positive link between high defensive advantages among rebel groups and the persistence of civil wars, the preceding discussion accordingly leads to the following testable hypothesis:
Hypothesis 1 provides a simple and powerful prediction of the relationship between civil war duration and malaria prevalence. However, a key assumption within the theoretical argument put forth previously is that rebel groups will typically be smaller, weaker, and more diffuse than government forces. This is often, but not always, the case (Cunningham, Gleditsch, and Salehyan 2009). Under the circumstances where rebel groups are not substantially smaller and weaker than government troops, the aforementioned malaria induced advantages for rebel groups’ abilities to prolong civil war, especially with respect to the first causal mechanism discussed previously, are likely to be lessened (to nonexistent). Specifically, any disproportionate phase 1 advantages held by rebel groups will subside as rebel forces grow in power (or size) and accordingly shift their fighting strategies away from diffuse, embedded guerilla tactics toward more traditional military deployments (and thus a phase 2 environment). At the extreme, some exceedingly powerful rebel groups may even exhibit comparable troop rotation practices to those of rival government forces. In these respects, “weak rebel group strength” represents a necessary condition for several of my identified causal mechanisms, and thus a second implication of my theoretical argument is that Hypothesis 1 should be strongest when rebel groups are weaker than government forces. Hence, the subsequent analysis will also seek to evaluate the following implication:
Empirical Analysis
Sample and Dependent Variable
The previously mentioned hypotheses suggest that high malaria prevalence will prolong civil wars, especially when rebel groups are weaker than government forces. To test these hypotheses on a collection of all relevant cases, I begin with Cunningham, Gleditsch, and Salehyan’s (2009) 1945–2003 sample of civil war durations—which corresponds to an expanded coding of the civil wars identified in the Uppsala/PRIO Armed Conflict Data (ACD)—and I then extend these data forward by 1 year, to 2004, with the aid of the most recent release of the ACD (Gleditsch et al. 2002; Strånd 2006). Accordingly, my sample encompasses the most comprehensive set of civil wars currently available and includes any violent conflict involving state and rebel actors that generated at least twenty-five casualties in a given calender year. Akin to similar analyses, the resultant 404 unique civil conflicts (1945–2004) are recorded at the government-insurgent (i.e., conflict dyad) level of observation. 7
Next, and using the binary measure of coup d’états provided by Cunningham, Gleditsch, and Salehyan (2009), I remove all conflicts corresponding to coups because these conflicts could be reasonably argued to not correspond to civil wars per se. My dropping of these cases is consistent with Buhaug, Gates, and Lujala (2009), and I further demonstrate in the Supplemental Appendix that my results remain unchanged when coups are instead included in the sample and controlled for. In sum, my dependent variable is therefore best characterized as civil war duration and is measured as the number of months since the onset of a given civil conflict. 8 For civil war duration, each conflict dyad is then observed for each year in which it is ongoing or terminated, which allows me to include time-varying covariates specific to a given conflict dyad, or its component parts (i.e., state or rebel group). Following Cunningham, Gleditsch, and Salehyan (2009), conflicts experiencing a lull of two calender years or more in fighting are treated as separate conflict dyads. I provide detailed summary statistics for this dependent variable in the Supplemental Appendix.
Estimation Model
I use semiparametric Cox proportional hazard models with standard errors clustered on conflict dyad id to estimate the effects of my covariates on a dyad’s civil war duration. As such, the Cox models used subsequently specifically estimate the hazard rate of civil war termination at month t for a given conflict id (i) as a function of a baseline hazard of civil war termination, ho
(t), and my covariates (
Independent Variables
The first independent variable of interest for the study at hand is a country’s (or region’s) malaria prevalence prior to a civil conflict’s onset. Adding malaria prevalence to my sample involved three steps. First, data on the percentage of a country’s (or territory’s) land area with malaria in the years 1946, 1966, 1982, and 1994 were obtained from the Center for International Development (CID) Malaria data set (Gallup, Mellinger, and Sachs 2001). Second, geographic malaria prevalence was then matched to the regions (typically a country or territory) of conflict specified by Cunningham, Gleditsch, and Salehyan (2009) within the conflict dyad cases mentioned previously. Third, because civil wars themselves may increase a region’s malaria prevalence (Hendrix and Gleditsch 2012), the malaria prevalence at the time prior to a conflict’s onset was assigned as my malaria measure for each conflict country/region in my sample such that, for example, a conflict beginning anytime between January 1983 and December 1994 was assigned that conflict area’s recorded malaria prevalence in 1982 (for that conflict dyad’s entire duration), whereas a civil war beginning in December 1982 was instead assigned that area’s 1966 recorded malaria prevalence for each observed year–month of that conflict’s duration. Because the earliest available CID measure of malaria prevalence was for the year 1946, the twenty-nine civil wars that started in 1945 or 1946 were then dropped from my sample. The exclusion of these cases did not influence my primary results. As a whole, these coding decisions thereby establish malaria prevalence’s temporal precedence (over civil war duration), while also providing for a hard test of my theory, as in many cases a conflict dyad’s malaria prevalence was recorded five to ten years before the outbreak of a given civil war. 9
I use the proportion of a country’s land area with malaria to measure malaria prevalence, rather than the (oft used) proportion of a country’s population living in malarial regions (Gallup, Mellinger, and Sachs 2001), for two key reasons. First, because civil wars are often fought far from a country’s major urban centers (which tend to have lower malaria prevalence than rural regions), using a population-based malaria measure would systematically bias my malaria prevalence measure downward—relative to the actual malaria prevalence observed in conflict zones. For instance, while malaria was endemic across roughly 80 percent Colombia during the 1966–1994 period (Gallup, Mellinger, and Sachs 2001), only 40 percent of Colombia’s population are recorded as living in malaria regions by the CID during these intervals, largely due to the atypically low malaria rates of Bogotá and Medillín. This latter measure would thereby underestimate the malaria rates found within the (predominately rural) areas of government-FARC conflict for my sample, which primarily lie in the highly malarial low lying provinces of eastern Colombia. Second, and most important, any use of the CID’s population-based malaria measure would directly undermine my ability to establish malaria prevalence’s temporal precedence vis-à-vis civil war duration—an absolute necessity in studies of disease and civil war (Hendrix and Gleditsch 2012). Indeed, while the CID records malarial population-exposure rates for the same time intervals mentioned previously (i.e., 1946, 1966, 1982, and 1994), in each case, it uses geographic population projections for the year 1995 (along with 1946–1994 geographic malaria proportions) to derive its population percentages. Thus, in using the CID’s population-based malaria measure as an independent variable, I would frequently be assessing the malarial effects of mid-1990s population distributions on the durations of civil wars occurring (and concluding) during the 1950s through the 1980s. Given that civil wars themselves can affect countries’ subsequent population distributions (Toole 1995), such an approach would be severely damaging to inference.
The second hypothesis discussed previously suggests that weak rebel groups are a necessary condition for malaria prevalence to have its conflict-prolonging effects. To test this theory, I construct a measure of aggregate rebel group strength (rebel strength) and interact this measure with malaria prevalence. To operationalize rebel strength, I use Cunningham, Gleditsch, and Salehyan’s (2009, 581) ordinal indicator of “the relative strength of rebels as either ‘strong,’ ‘at parity,’ or ‘weak’ in terms of their ability to fight conventional wars,” which I set to range from 0 (weak) to 2 (strong). The four component parts to this index are drawn from the nonstate actors involved in intrastate conflicts (NSA) data set (Cunningham, Gleditsch, and Salehyan 2008) and include (1) whether a rebel organization has a clear central command (2) the mobilization capacity (in terms of number of available fighters) of a rebel group, (3) a rebel group’s ability to procure arms, and (4) rebel fighting capacity. I provide a more detailed discussion of each of these component parts in relation to my theory, along with summary statistics for rebel strength and malaria prevalence, in the Supplemental Appendix. Finally, as mentioned previously, I then interact rebel strength with malaria prevalence (malariaXrebel-strength) and include this interaction term and its two component parts to test Hypothesis 2 in my “full” Cox proportional hazard model specification.
Control Variables
I also add a number of control variables to my Cox proportional hazard models. Because the type of civil war may influence the relationship between rebel strength and civil war duration (Fearon 2004; Cunningham, Gleditsch, and Salehyan 2009), I include binary indicators for whether (or not) a given civil war can be characterized as a war on core territory, an ethnic conflict, or as a war involving two or more dyads. 10 For similar reasons, I also control for a conflict dyad’s index of national ethnic and linguistic fractionalization (ELF), taken from Fearon and Laitin (2003), as it could be argued that ethnic diversity is both (1) correlated with the presence of genetic barriers to malaria transmission (De Silvestri and Guglielmino 2000) and (2) positively associated with civil war duration (Collier, Hoeffler, and Söderbom 2004). The presence (or absence) of democracy has been found to be related to civil war duration (Cunningham, Gleditsch, and Salehyan 2009; Buhaug, Gates, and Lujala 2009), and through development, may itself be inhibited by high rates of malaria. Hence I also add a binary democracy indicator—coded 1 for country-years with a Polity IV (Marshall, Jaggers, and Gurr 2010) value higher than six—as a control in the models reported subsequently. I choose to include a binary democracy indicator so as to ensure that my specifications and results are as comparable as possible to past research on civil war duration (e.g., Cunningham, Gleditsch, and Salehyan 2009), though as shown in the Supplemental Appendix, the findings discussed subsequently are robust to the use of an ordinal Polity IV indicator in its place. The degree of a rebel group’s control over territory is a theoretically distinct construct from that of rebel strength (Cunningham, Gleditsch, and Salehyan 2009). Nevertheless, territorial control is likely to be correlated with rebel strength, in addition to exhibiting a positive effect on overall civil war duration (Cunningham, Gleditsch, and Salehyan 2009). I therefore include the NSA’s dichotomous indicator of whether (or not) a rebel group controls any territory as an additional control.
The remaining control variables are included to better account for the potential biases of omitted variables in my evaluation of the effects of malaria prevalence on civil war duration. According to extant research, malaria is highly persistent across many of the more populous regions of the world (Hay et al. 2004), whereas low levels of economic development and state capacity can often undermine a country’s ability to successfully reduce malaria prevalence (Sachs and Malaney 2002). At the same time, economic development, population size, and state capacity are several of the most commonly identified correlates of civil war duration (Collier, Hoeffler, and Söderbom 2004; Fearon 2004; Cunningham, Gleditsch, and Salehyan 2009). I thus include controls measuring the natural log of a country’s population and of gross domestic product (GDP) per capita, taken from Gleditsch (2002). However, because the use of GDP per capita as a proxy for state capacity has been called into question (Hendrix 2010), I also explore several more direct (albeit less temporally comprehensive) controls for state capacity in the Supplemental Appendix. Adverse terrain, such as forest cover, is frequently theorized to prolong civil war, and yet is often found to be insignificant predictor thereof (e.g., Buhaug, Gates, and Lujala 2009; Thyne 2012). Nevertheless, because malaria is inherently linked to geography, I follow the past studies of malaria’s effects on political–economic outcomes (e.g., Sachs and Malaney 2002) by controlling for a country’s tropical location—via either the percentage of a country’s landmass that lies in the geographical tropics (main models) or the (ln) latitude of a country’s geographic centroid (robustness models)—both taken from the CID (Gallup, Mellinger, and Sachs 2001).
In recent decades, the burdens of Malaria have proven to be particularly acute for Africa (Sachs and Malaney 2002), a region that is often characterized as exceptional with regard to civil war persistence (de Rouen and Sobek 2004). To ensure that no unobserved factors inherent to this region are influencing my findings, I complete my primary model specifications with an Africa region dummy. Finally, note that I evaluate and discuss a wide variety of additional controls—including many related to geographic factors, state capacity, additional diseases, and temporal adjustments—within the robustness section and Supplemental Appendix to this article. I refrain from adding many of these additional controls to the primary models reported here given the many problems associated with models using excessive control variables (Achen 2005), and because several of these controls lead to more limited sample sizes due to missingness or comparability issues.
Results
I present the main results for civil war duration sequentially in Table 1. This table begins with a baseline specification for malaria prevalence (Model 1), to which I then add the remaining controls described previously to arrive at a primary test of Hypothesis 1: malaria prevalence’s effects on civil war duration (Model 2). The final model reported in Table 1 (model 3) appends model 2 with the interaction term described previously (malariaXrebel-strength) so as to fully test Hypothesis 2. Importantly, because the coefficient estimates in Table 1 report the effects of my covariates on the baseline hazard of a conflict’s termination, positive coefficient estimates are associated with increases in the hazard of conflict termination (implying shorter civil war durations) and negative estimates are interpreted as decreasing the baseline hazard of conflict termination (thereby prolonging the duration of civil war). To directly evaluate my primary results, I first assess Hypothesis 1 via models 1 and 2 and then evaluate Hypothesis 2 (and Model 3).
Cox Proportional Hazard Estimates of Civil War Duration, 1947–2004.
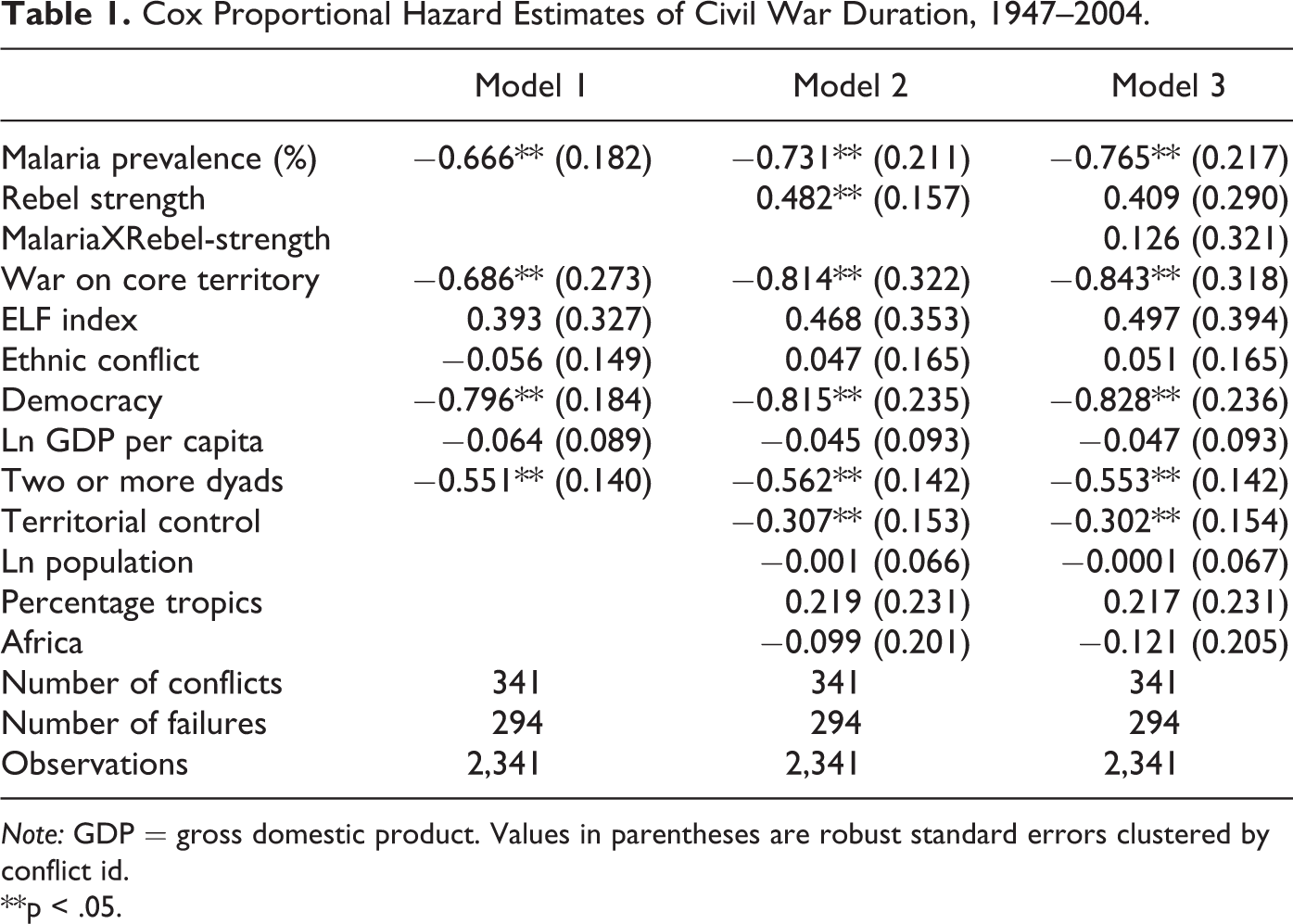
Note: GDP = gross domestic product. Values in parentheses are robust standard errors clustered by conflict id.
**p < .05.
Turning to Models 1–2, I find that the coefficient estimates for malaria prevalence are negative and statistically significant at the p < .01 level in both models. In support of Hypothesis 1, this implies that increases in the malaria prevalence of a country or region are associated with decreases in the baseline hazard of civil war failure and hence lead to longer civil war duration. As for the statistically significant control variables in models 1 and 2, one can note that the coefficient estimates for wars on core territory and (rebel) territorial control are negative and significant, confirming Cunningham, Gleditsch, and Salehyan’s (2009) findings that noncolonial civil wars, and civil wars with substantial rebel territorial control, typically last longer than do civil wars exhibiting less territorial salience. Likewise, democracy and two or more dyads are also negative and statistically significant (p < .01) in models 1 and 2. Hence, democratic countries, and civil wars involving multiple active dyads, each experience extended civil war durations. Finally, rebel strength is independently positive and significant in model 2, implying that increases in rebel strength decrease the duration of civil wars.
To assess whether the previously mentioned findings are substantively notable, I use the estimates from model 2 to plot a series of survival curves of civil war duration, given various values of malaria prevalence, as well as for different values of three additional independent variables that achieved statistical significance at least at the p < .05 level in model 2: rebel strength, democracy, and territorial control. The two most common (and policy-relevant) values for malaria prevalence in my sample are 0 (i.e., 0 percent) and 1 (i.e., 100 percent), which together encompass over half of all observations, and hence I examine the effects of malaria prevalence on civil war survival at these two intervals (i.e., for a 0-to-1 change). For my other plotted variables, I examine comparable changes, specifically a 0-to-2 (i.e., weak to strong) change in rebel strength, a 0-to-1 change in democracy, and a 0-to-1 change in rebels’ degree of territorial control.
Figure 1 presents the resultant collection of survival curves for civil war duration. For each subfigure, substantive effects were calculated while holding all other variables at their means or modes. In reporting survival on the y-axis, these graphs present the probability that a specific conflict dyad does not terminate at a particular point in time, given the passage of a certain number of years following a conflict’s onset (x-axis). Beginning first with malaria prevalence, one finds that—for any point in time—the predicted likelihood of civil war termination is higher for conflicts in low malarial regions than it is for conflicts within high malarial regions. Hence, the duration of civil war for high malaria prevalence regions is notably longer than that of civil wars in low malaria prevalence regions. Moreover, this effect is also notable in its substantive size: at the median level of civil war duration for my sample (five years), eliminating malaria from a high malaria prevalence country is estimated to increase the likelihood of a civil war’s immediate termination from roughly 50 percent to 80 percent—a 30 percent increase.
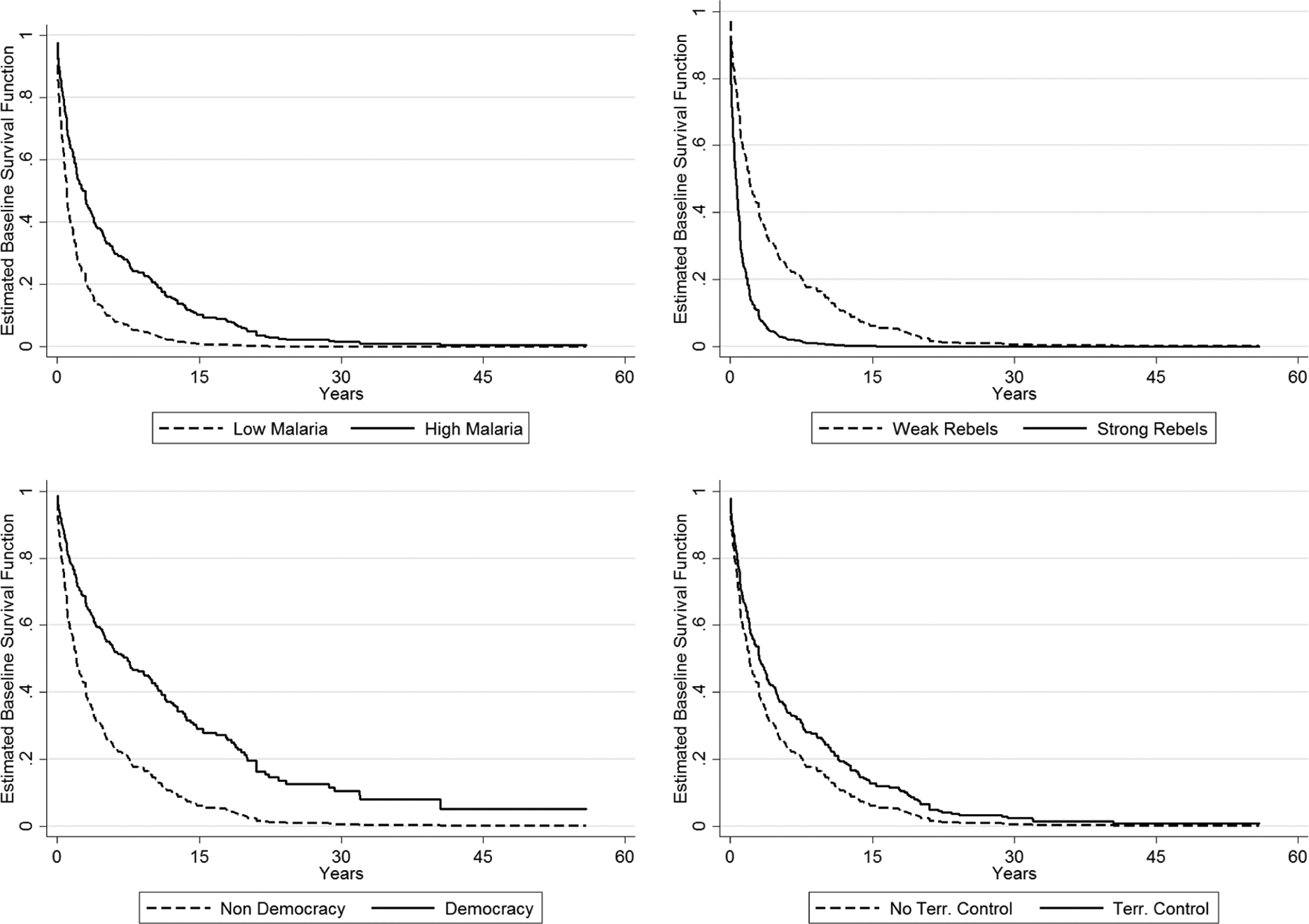
Comparisons of conditional survival rates for civil war.
The additional survival curves in Figure 1 situate the substantive findings for malaria prevalence (discussed previously) within our extant knowledge of civil war duration. Consistent with the findings and theory discussed in Cunningham, Gleditsch, and Salehyan (2009), increases in relative rebel strength (from weak to strong) are associated with faster civil war termination—with the resultant change in a conflict’s survival probability appearing to be comparable in size to the effects of malaria prevalence. Also consistent with past research (Cunningham, Gleditsch, and Salehyan 2009; Buhaug, Gates, and Lujala 2009), nondemocracies are estimated to have a higher probability of civil war termination (and hence shorter civil war duration) than are democracies, for all periods after a civil war onset—an effect that is significantly larger than that discussed for malaria prevalence previously. Finally, and similar to the effects of malaria prevalence, a rebel group’s achievement of territorial control is estimated to prolong the duration of a given civil war, at all periods since civil war inception, though in this case the effect of territorial control appears to be notably smaller than the comparably adverse effects of malaria prevalence. In sum, malaria appears to prolong civil war and does so with a magnitude that is comparable to several more frequently studied correlates of civil war duration.
To evaluate whether the conflict-prolonging effects of malaria are conditional on (weak) rebel strength (Hypothesis 2), I next turn to model 3 in Table 1. Before directly assessing this model’s support for Hypothesis 2, however, I first give mention to the control variable findings for this model. As found previously, democracy and territorial control are each negative and significant, suggesting that both of these covariates decreases the baseline hazard of civil war termination, thereby extending conflict duration. Civil wars fought on a government’s core territory as well as civil wars involving multiple dyads/actors are each estimated to last longer in model 3, perhaps owing to the higher degree of salience that is implied by conflicts involving these factors. On the other hand, the controls for ln GDP per capita, ln population, percentage tropics, ethnic fractionalization and Africa are insignificant in each model specification, suggesting perhaps that once the effects of malaria prevalence and my other controls are accounted for, the former controls have little significant direct effect on civil war duration.
I make use of model 3’s interaction term (malariaXrebel-strength) as well as each of its component parts (malaria prevalence and rebel strength), to fully evaluate the level of statistical support for Hypothesis 2. Recall that rebel strength is measured 0 to 2, with 0 values implying cases where rebels are recorded as being “weak.” Hence, in adding malariaXrebel-strength to my model specification, my individual malaria prevalence term is now interpreted as the direct effect of malaria prevalence when rebel strength equals 0 (i.e., for weak rebel groups), whereas the effect of malaria prevalence in parity and strong rebel group cases (i.e., when rebel strength >1) is now evaluated as the sum of malaria prevalence, malariaXrebel-strength, and rebel strength. In this regard, I do not expect a significant interaction term coefficient estimate, but rather, I expect only the individual malaria prevalence estimate to be (negative and) significant, as this composite term provides the estimate of malaria prevalence under weak rebel strength. Critically, my focus on—and interpretation of—the individual malaria prevalence coefficient term (rather than that of malariaXrebel-strength), is consistent with extant research on interaction terms, as it corresponds directly to the (unusual) hypothesis test case of “
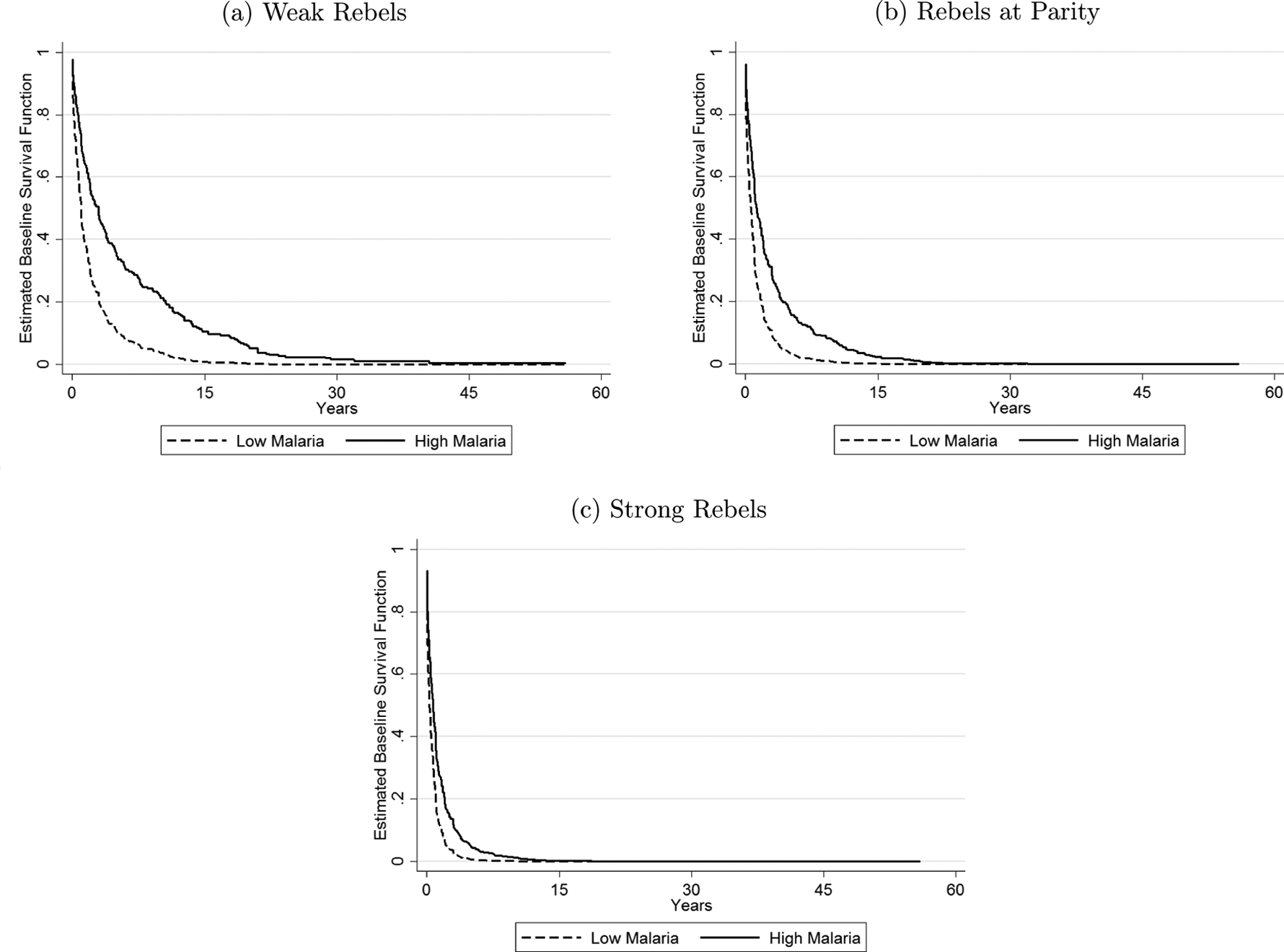
MalariaXRebel-strength survival rates for civil war.
Specifically, Figure 2 plots survival curves for civil war duration, given 0-to-1 changes in malaria prevalence, separately for rebel groups that are weaker than government forces (Figure 2a), at parity with government forces (Figure 2b), or stronger than government forces (Figure 2c). As before, all other controls are held to their means or modes. Figure 2a accordingly demonstrates the effect of malaria among weak rebel group conflicts, as derived from the individual malaria prevalence coefficient in model 3. Consistent with my previous discussion, this subfigure indicates that a 0-to-1 increase in malaria prevalence is estimated to prolong the duration of a given civil war, at all periods of time since a civil war’s inception—when a rebel group is significantly weaker than government forces. Similar to my earlier analysis, at the median level of civil war duration for the sample (five years), this effect implies that reducing the malaria prevalence or a country or region from its most common value in my sample (i.e., 1, or 100 percent) to its second most common value (0 percent) is estimated to increase a civil war’s likelihood of immediate termination from roughly 50 percent to 82 percent—a more than a 30 percent improvement. By contrast, a comparable change in malaria prevalence for rebel groups at parity with government forces (Figure 2b) is estimated to at best have a negligible effect on civil war duration, whereas among strong rebel groups, this effect disappears entirely—a 0-to-1 change in malaria prevalence has a near zero effect on a conflict dyad’s survival rate of civil war duration, for all periods ex post to civil war onset. Therefore, and in support of Hypothesis 2, malaria prevalence is associated with increases in civil war duration only among civil wars involving weak rebel groups.
The results discussed previously are robust to an extensive array of alternate model specifications. To save space, these robustness models are fully discussed and presented in the Supplemental Appendix. Stated briefly, my findings for malaria prevalence are robust to the inclusion of additional or alternate control variables, including multiple measures of state capacity, additional diseases such as yellow and dengue fever, geographic variables such as (1) country latitude, (2) a conflict zone’s distance to the capital city and proximity to border regions (and the interactions of these two variables), (3) the presence of various illicit goods in a conflict zone, and (4) the proportion forest or mountain cover within a conflict zone, and alternate measures of GDP and democracy. The models presented previously are also highly robust to alternate operationalizations of my dependent variable, including a civil war duration measure that also treats coups as civil wars, a measure that removes outlier cases, and a dependent duration variable that was aggregated at the conflict dyad, rather than rebel group conflict dyad level of aggregation. Likewise, I further establish that all key findings remain, and generally become stronger, when an interpolated malaria prevalence measure—that uses a region or country-year’s most temporally proximate recorded geographic malaria levels to assign its level of malaria prevalence 11 —is employed as my primary independent variable. In addition, the Supplemental Appendix also demonstrates that my core conclusions persist under various alternative frameworks for handling the temporal and spatial dependencies in my data, including year fixed effects, a number of frequently used parametric survival model estimators, and the inclusion of a multiplicative unit specific frailty term for conflict dyad id.
Conclusion
This article develops a theoretical argument to suggest that malaria prevalence will prolong civil wars. In particular, I theorize that malaria prevalence can inflict asymmetrically high costs upon government forces—relative to rebel groups—due to the former’s (1) larger size and less diffuse fighting style, (2) propensity for frequent troop rotations into and out of malarial conflict regions, and (3) dependence upon establishing a physical presence, building infrastructure, and providing public services in conflict regions in order to secure victory. As argued previously, each of these factors is adversely affected by a region’s pre-conflict levels of malaria in a manner that enhances the defensive capabilities of rebel groups. Consequently, when present, malaria is anticipated to enable rebel groups to avoid total defeat and to thus prolong the duration of civil war. In support of these expectations, I empirically establish that geographic malaria prevalence significantly increases the duration of civil wars—particularly for those civil wars in which rebel groups are decidedly weaker than government forces. Given the high social burdens of both malaria and civil war, these linkages have important policy relevance. Specifically, the findings discussed previously not only identify another avenue for malaria’s imposition of (indirect) costs upon developing countries but also suggest a possible mechanism by which the societal costs of current and future civil wars can be ameliorated through the eradication of malaria.
This article’s findings also have a number of important implications for the study of political violence. First and foremost, the previously mentioned results suggest that malaria prevalence can often turn superior offensive capabilities—which are traditionally seen as a major asset in intra- and interstate warfare—into a distinct disadvantage. As such, this article helps to explain why superior offensive capabilities have not always translated into (timely) military victory on the battlefield. In this regard, taking malaria more seriously may help to improve our understandings of historical cases such as the United States–Vietnam War, the FARC rebellion in Colombia, and the more general fragmentation patterns of countries across the globe through successful and unsuccessful independence movements. Likewise, and given that malaria abatement technologies and medicines are poorly abided by among troops in the field (Gambel et al. 1998; Joy 1999, 203), the previously mentioned theory and findings together suggest that governments faced with rebellions in malarial regions will be most successful in quickly ending these rebellions when they utilize more diffuse, and more embedded, guerilla fighting styles, as opposed to traditional offensive military tactics. Finally, and in line with extant research (Letendre, Fincher, and Thornhill 2010, 2012), the dynamics discussed previously, for malaria, may similarly hold for other major diseases—especially other mosquito-borne diseases. A number of initial findings presented in the Supplemental Appendix support this point, wherein the additional disease controls in Table A6 suggest that dengue fever, like malaria, may have significant civil conflict-prolonging effects. 12 Thus, further research into the effects of other preeminent infectious diseases on conflict and conflict duration is highly warranted.
The theory and findings discussed previously may also improve our understandings of the effects of malaria—and related diseases—on economic development, and on politics more generally. Extant arguments for the socioeconomic burden of malaria (e.g., Sachs and Malaney 2002) generally characterize this burden as a unit shift in the costs incurred by a society or country. By contrast, this article suggests a more nuanced relationship, wherein, above and beyond its baseline costs, malaria imposes compounding costs specifically upon those social and political activities that necessitate large, but un-urbanized, infusions of human populations (and population densities) into sparsely populated malarial regions. In addition to government troop deployments during civil wars, a number of additional political–economic activities will likely be burdened by these multiplicative effects. For example, voter turnout and election violence could be affected by malaria in a similar manner, not only due to the increased potential for election day dissuasion but also due to a comparative lack of transportation and polling station infrastructure in malarial regions. The construction of major public works projects, including dams, roads, power plants, and canals will also be particularly affected on these grounds—which implies that the burdens of malaria may inhibit not only modernization but also relations between neighboring states, and regional integration. Similar to the civil conflict dynamics laid forth previously, malaria prevalence may also inhibit governments’ abilities to police other diffuse human activities within their boarders, such as those pertaining to (organized) crime, terrorism, and the sale and transportation of illicit goods. With the socially interactive theory of malaria offered previously, these potential linkages are ripe for further examination.
Footnotes
Author’s Note
I would like to acknowledge the valuable suggestions that I received from the editor and reviewers of the Journal of Conflict Resolution and Joe Wright.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.