
Abstract
Tooth extraction is a fundamental dental intervention for managing oral infections; however, the resulting tooth loss is associated with systemic health risks, including cognitive decline and increased mortality. Alveolar bone resorption following tooth extraction significantly alters jaw morphology, complicating subsequent functional rehabilitation. Despite its clinical significance, the cellular and molecular mechanisms governing bone regeneration within extraction sockets remain poorly understood. Here, we identified group 2 innate lymphoid cells (ILC2s) as pivotal regulators of alveolar bone regeneration following tooth extraction. Using single-cell RNA sequencing and flow cytometry, we demonstrated that ILC2s accumulate and persist within extraction sockets beyond the acute inflammatory phase. Genetic lymphocyte ablation revealed that loss of all ILCs severely impairs bone formation, highlighting their essential role in socket regeneration. Following tooth extraction, ILC2s exhibited increased interleukin (IL)–13 production and promoted bone formation via IL-13–mediated activation of bone marrow–derived mesenchymal stromal cells. Single-cell RNA sequencing and histological analyses suggested that gingival fibroblasts are a major source of IL-33 in extraction sockets during mucosal healing. IL-33 activated ILC2s, increasing IL-13 production and promoting osteogenesis, whereas IL-33 neutralization suppressed ILC2 activation and impaired socket bone regeneration. Collectively, these findings suggest that the gingival fibroblast–IL-33–ILC2–IL-13 axis may contribute to the regulation of alveolar bone regeneration following tooth extraction. This study provides a biological basis for developing novel therapeutic strategies to optimize postextraction bone preservation.
Introduction
Tooth extraction is a common procedure that often leads to pronounced alveolar bone resorption (Kondo, Kanayama, et al 2023), thus complicating subsequent prosthodontic treatment (Ohtake et al 2025; Kondo et al 2026) and potentially impacting systemic health, including cognitive decline and increased mortality (Koka and Gupta 2018; Ma et al 2025). The healing of extraction sockets involves a coordinated immune response where various immune cell subsets, including neutrophils (Ando et al 2024), macrophages (Okawa et al 2022), and lymphocytes (Tsukasaki et al 2018; Kondo, Gleason, et al 2023; Shah et al 2025), regulate the transition from acute inflammation to bone formation (Kondo et al 2024a; Otake et al 2026). While the general stages of socket healing are well documented (Kamal et al 2025), the specific molecular axes bridging the initial inflammatory phase with effective alveolar bone regeneration remain to be fully elucidated.
Innate lymphoid cells (ILCs) are a rare population of haematopoietically derived immune cells that lack antigen-specific receptors and lineage-specific markers, functionally mirroring T helper cells (Moro et al 2010). ILCs are classified into the following 3 major groups based on their cytokine production profiles: natural killer (NK) cells, group 1 ILCs (ILC1s), ILC2s, and ILC3s. ILCs rapidly respond to local cytokine signals and are essential for maintaining barrier integrity and promoting tissue repair (Walker et al 2013). ILC2s exhibit tissue-specific functions depending on the local microenvironment. For example, adipose tissue–resident ILC2s regulate beige adipocyte formation through interleukin (IL)–13 production (Brestoff et al 2015), whereas lung-resident ILC2s promote epithelial repair following viral infection through amphiregulin secretion (Monticelli et al 2011). In the context of bone biology, ILC2s can regulate bone metabolism primarily through IL-13 production (Vivier et al 2018). Notably, IL-13 produced by ILC2s influences osteoclast activity and bone resorption (Omata et al 2018, 2022; Momiuchi et al 2021).
ILC2s are activated by IL-33 through its receptor ST2 (Amisaki et al 2025). IL-33 is predominantly produced by epithelial cells, endothelial cells, and fibroblasts and is released in response to tissue injury (Griesenauer and Paczesny 2017). Activation of the IL-33–ILC2 axis has been extensively documented in inflammatory diseases such as asthma (Barlow et al 2013), obesity (Ding et al 2016), and cancer (Alam et al 2022); accumulating evidence indicates that this signaling pathway also contributes to tissue repair. Fibroblasts are increasingly recognized as active regulators of tissue regeneration. Through cytokine production, fibroblasts shape the local immune microenvironment and influence immune cells and skeletal cells. Gingival fibroblasts modulate the activity of immune cells, osteoblasts, and osteoclasts, thereby contributing to the regulation of alveolar bone remodeling (Akashi et al 2023; Kondo, Gleason, et al 2023; Kondo, Kanayama et al 2023). Despite these findings, the roles of ILC2s in regulating osteoblast activity and alveolar bone regeneration remain poorly understood. In particular, cellular IL-33 sources and the mechanisms through which the IL-33–ILC2 pathway regulates bone regeneration within tooth extraction sockets have not yet been fully elucidated.
In this study, we aimed to elucidate the role of ILC2s in the complex process of extraction socket healing. Specifically, we sought to identify the cellular sources that trigger ILC2 activation and molecular pathways through which these cells regulate bone regeneration. Our results demonstrated that ILC2s are essential for postextraction bone formation, acting via an IL-13–mediated enhancement of osteoblastic activity. Furthermore, we revealed that this regenerative response is orchestrated by gingival fibroblast-derived IL-33, thereby establishing a novel fibroblast–IL-33–ILC2–IL-13 axis that governs oral osteoimmunology.
Materials and Methods
Details are provided in the Appendix.
Results
ILCs Contribute to Bone Formation in Tooth Extraction Sockets
After extraction of the maxillary first molars in wild-type (WT) mice, the oral mucosa gradually extended coronally, fully covering the sockets by day 16 (Fig. 1A). Micro–computed tomography (CT) analysis demonstrated that bone formation within the sockets initiated on day 8. By day 32, the extraction sockets were filled with newly formed bone (Fig. 1B, C), which was consistent with the histological findings. These results indicate that this tooth extraction model is well suited for analyzing the process of bone regeneration in extraction sockets.
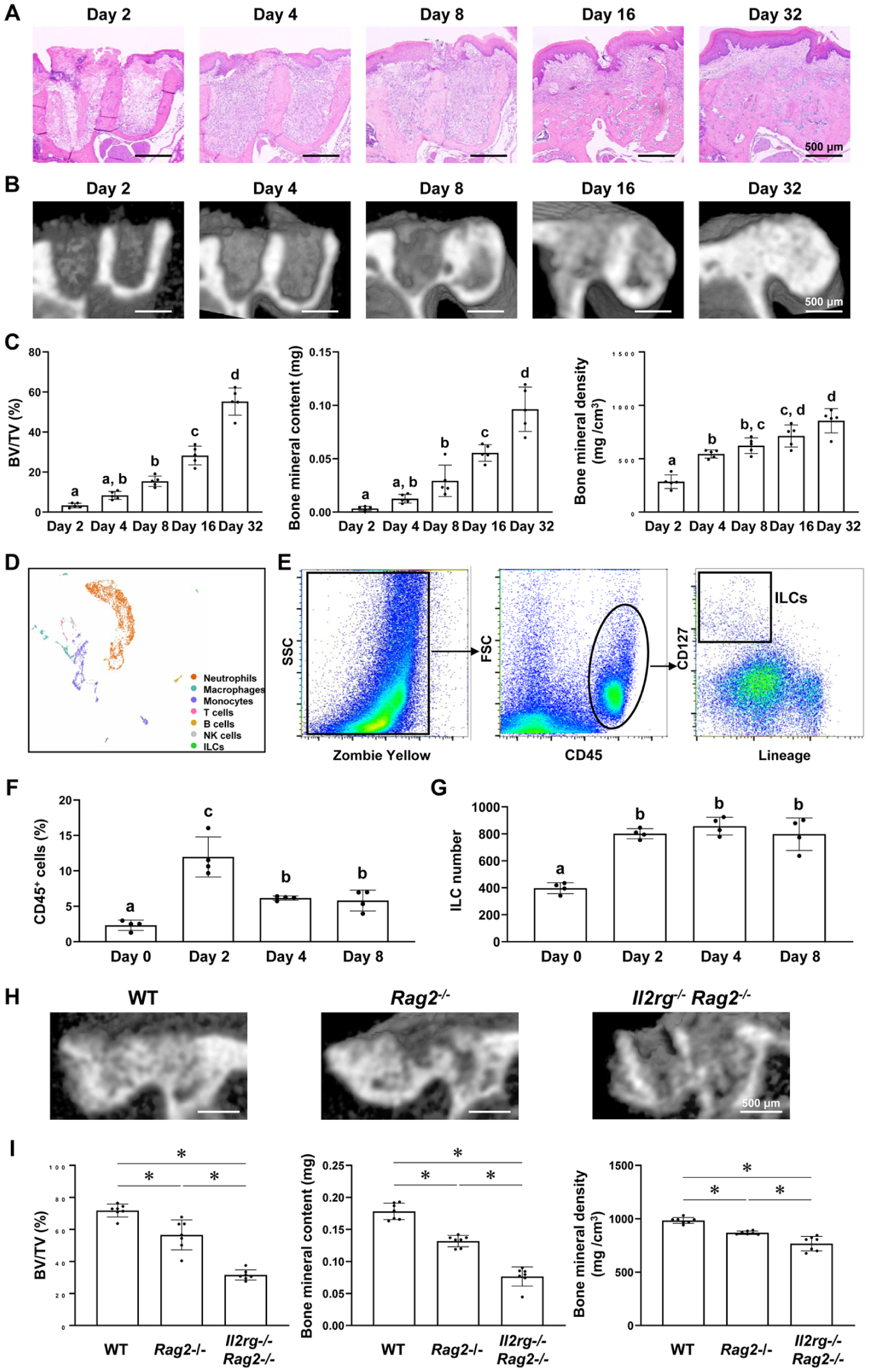
Innate lymphoid cells (ILCs) play important roles in bone regeneration in the tooth extraction sockets. (
Single-cell RNA sequencing (scRNA-seq) analysis identified a population of ILCs in the extraction sockets 8 d postextraction (Fig. 1D). To examine the temporal changes in total immune cells and ILCs, flow cytometry was performed on cells dissociated from the extraction sockets (Fig. 1E). The number of CD45+ immune cells, including neutrophils, macrophages, monocytes, T cells, B cells, and ILCs, increased rapidly by day 2 and declined after day 4 (Fig. 1F), reflecting the acute inflammatory phase. In contrast, the number of ILCs increased on day 2 and remained elevated beyond day 4 without a significant decline (Fig. 1G). These distinct kinetics suggest that ILCs may contribute to the regeneration of extraction sockets following acute inflammation.
To elucidate the specific roles of ILCs in socket regeneration, we used genetically modified mice lacking distinct lymphocyte subsets. Compared to WT mice, Rag2–/– mice, which lack T cells, B cells, and natural killer T (NKT) cells, exhibited significantly impaired bone formation. Notably, Il2rg–/–Rag2–/– mice, which lack all lymphocytes, including ILC1s, ILC2s, ILC3s, and NK cells, showed an even greater reduction in bone formation (Fig. 1H, I, Appendix Fig. 1). These findings underscore the essential role of ILCs in bone regeneration within extraction sockets, independent of—and complementary to—the functions of T cells, B cells, and NKT cells.
Temporal Dynamics of ILC Subsets in Extraction Sockets after Tooth Extraction
To identify the ILC subset critical for bone formation in extraction sockets, we analyzed the temporal changes of each population. ILC1s exhibited a modest increase and ILC3s a pronounced rise on day 2, following which both populations plateaued or declined. In contrast, the number of ILC2s continued to increase beyond day 2 and remained elevated (Fig. 2A–C). Based on these findings, we hypothesized that ILC2s play a specific role in socket regeneration. We then evaluated IL-13 production by ILC2s following tooth extraction and found that it increased significantly from day 4 onward (Fig. 2D, E). These results indicate that ILC2s may promote bone formation within tooth extraction sockets through the production of IL-13.
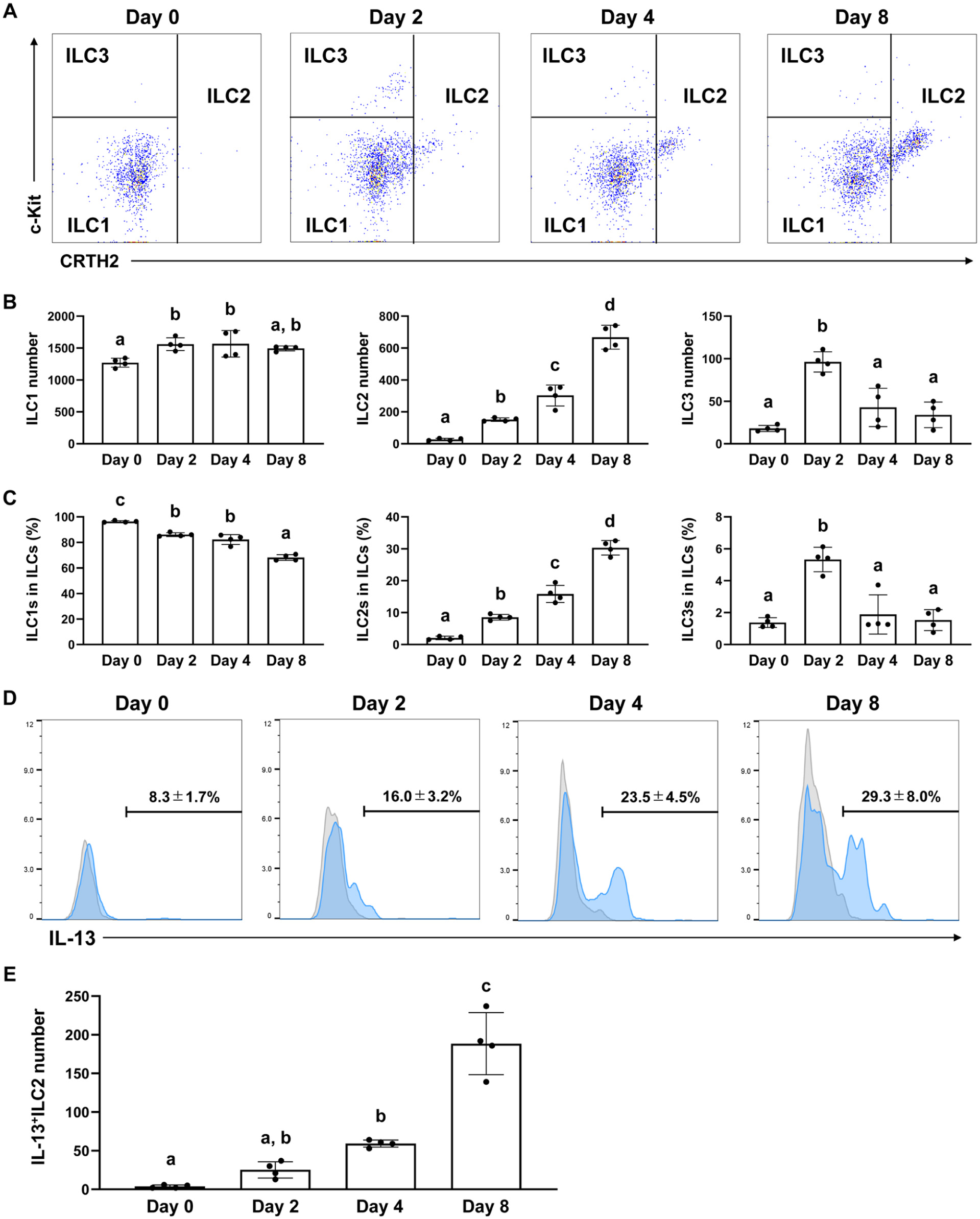
Changes in group 2 innate lymphoid cells (ILC2s) in the sockets after tooth extraction. (
ILC2s Promote Bone Formation in the Tooth Extraction Sockets through IL-13 Expression
Immunofluorescence staining of the extraction sockets revealed an increased number of IL-13–producing ILC2s following tooth extraction. These ILC2s were predominantly localized near the bone walls within the sockets rather than in the overlying oral mucosa (Fig. 3A). To investigate the potential role of ILC2-derived IL-13 in bone formation within extraction sockets, mouse bone marrow–derived mesenchymal stromal cells (BMSCs) were cultured in the presence of IL-13. IL-13 significantly enhanced matrix mineralization (Fig. 3B) and the expression of osteogenic marker genes (Runx2, Sp7, Alp, and Col1a1) (Fig. 3C) in BMSCs in a dose-dependent manner. In addition, IL-13 inhibited osteoclast differentiation in vitro. Tartaric acid and acid phosphatase (TRAP) staining of RAW264.7 cells cultured in osteoclast induction medium revealed that IL-13 reduced both the number and size of TRAP-positive multinucleated osteoclasts in a dose-dependent manner (Appendix Fig. 2). To examine its in vivo effects on socket bone regeneration, bone formation in mouse tooth extraction sockets was evaluated following the intravenous administration of IL-13. Compared with the phosphate-buffered saline (PBS)–treated control group, IL-13 administration significantly enhanced bone formation in the extraction sockets (Fig. 3D, E, Appendix Fig. 3). Collectively, these findings suggest that ILC2s promote bone formation within tooth extraction sockets by enhancing the osteogenesis of BMSCs via IL-13 production.
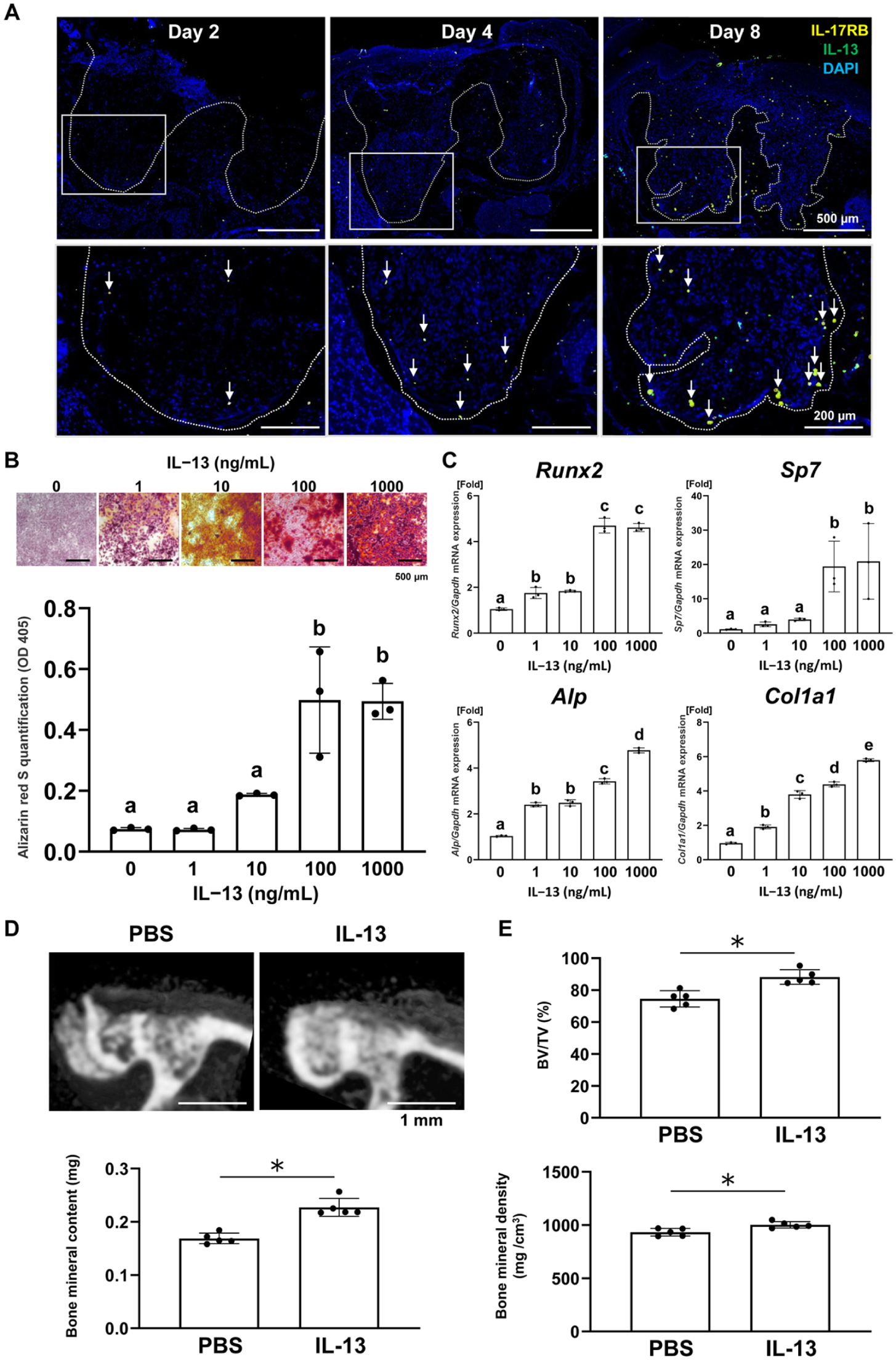
Group 2 innate lymphoid cells (ILC2s) promote bone formation in the tooth extraction sockets through interleukin (IL)–13 expression. (
Gingival Fibroblasts Are the Source of IL-33 after Tooth Extraction
scRNA-seq analysis demonstrated that ILC2s in the extraction sockets expressed Il1rl1, which encodes ST2, the receptor for IL-33 (Fig. 4A, B). To further validate the cellular source of IL-33 in oral tissues, we analyzed an independent scRNA-seq dataset derived from a mouse periodontitis model. Consistent with the extraction socket dataset, Il33 transcripts were predominantly detected in fibroblasts (Appendix Fig. 4). These findings suggest that IL-33 may contribute to ILC2 activation within the extraction sockets.
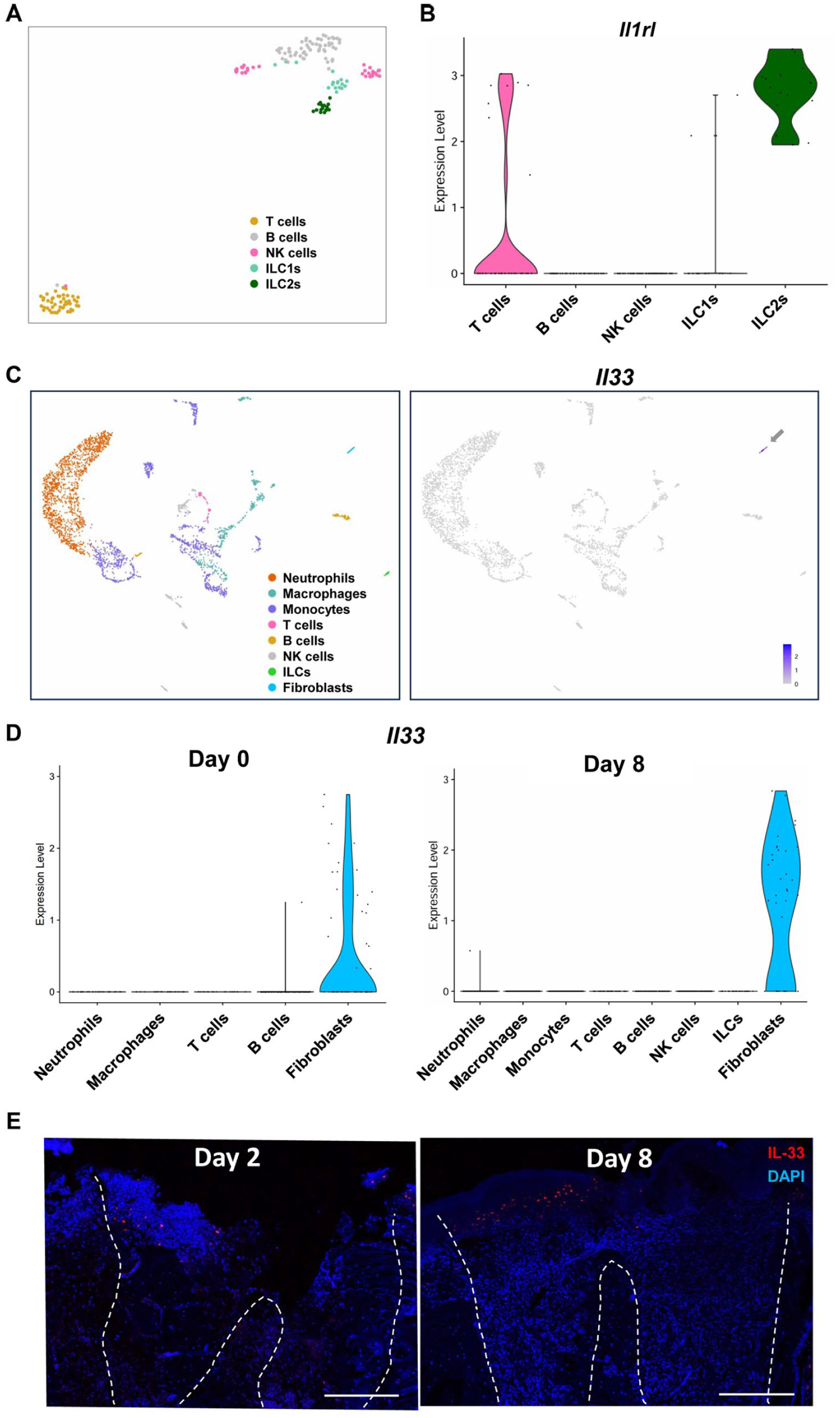
Gingival fibroblasts are the source of IL-33 after tooth extraction. (
We next identified the cellular sources of IL-33 in tissues surrounding the extraction sockets. High IL-33 expression was observed in the fibroblast cluster (Fig. 4C), indicating that gingival fibroblasts are the primary source of IL-33 in the extraction sockets. Moreover, Il33 expression in fibroblasts was upregulated on day 8 compared with that on day 0 (Fig. 4D). Histological analysis revealed that IL-33–positive oral mucosa gradually migrated into the coronal part of the extraction sockets (Fig. 4E). Collectively, these results indicate that gingival fibroblasts produce IL-33 as they infiltrate the upper region of the sockets during mucosal healing following tooth extraction. To further explore potential cellular interactions within the socket microenvironment, we performed CellChat analysis using our scRNA-seq dataset to predict intercellular communication toward ILCs. This analysis suggested that compared with other cell populations, fibroblasts provide strong predicted signaling inputs to ILCs (Appendix Fig. 5).
Gingival Fibroblasts Promote Bone Formation in the Tooth Extraction Sockets by Activating ILC2s via IL-33
To assess the effects of IL-33 on ILC2 activation in the extraction sockets, mice undergoing tooth extraction were intravenously administered either IL-33 or an anti–IL-33 monoclonal antibody. IL-33 administration significantly increased the number of ILC2s in the sockets (Fig. 5A, B). Furthermore, compared with the PBS-treated control, IL-33 enhanced IL-13 production by ILC2s (Fig. 5C, D) and significantly promoted bone formation in the sockets (Fig. 5E, F).
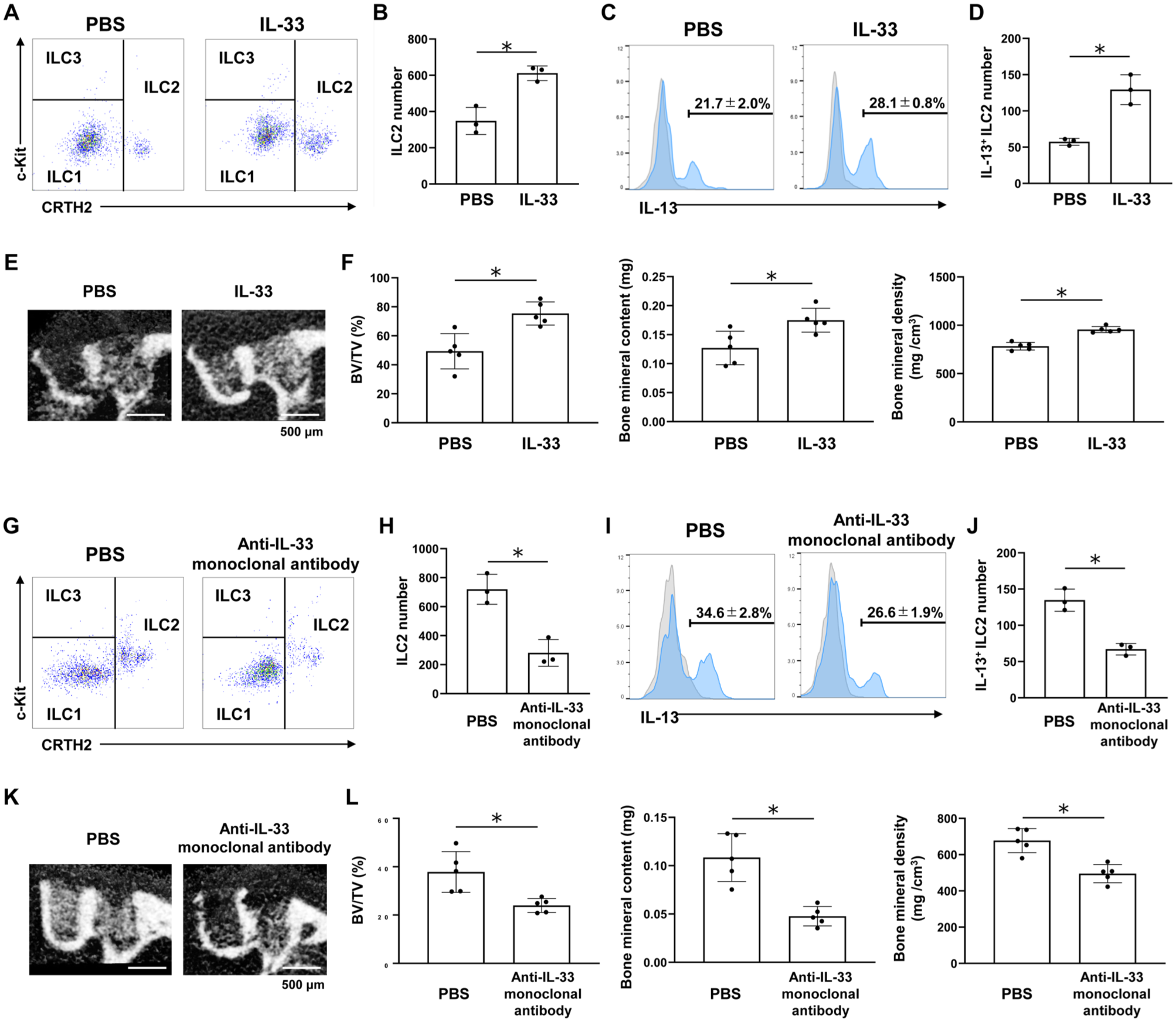
Interleukin (IL)–33 promotes bone formation in tooth extraction sockets through group 2 innate lymphoid cell (ILC2) activation. (
In contrast, administration of the anti–IL-33 monoclonal antibody significantly decreased the number of ILC2s in the sockets (Fig. 5G, H). Additionally, administration of anti–IL-33 monoclonal antibody suppressed IL-13 production by ILC2s (Fig. 5I, J) and resulted in significantly impaired bone formation relative to the control group (Fig. 5K, L).
Furthermore, IL-33 administration significantly promoted bone formation in Rag2-/- mice (Appendix Fig. 6). In contrast, IL-33 failed to enhance bone regeneration in Il2rg-/-Rag2-/- mice, which lack all ILC populations (Appendix Fig. 6).
To further examine whether IL-13 functions as a downstream mediator of IL-33 signaling, we performed rescue experiments under IL-33 blockade. Mice were treated with PBS, anti–IL-33 monoclonal antibody + PBS, or anti–IL-33 monoclonal antibody + IL-13 following tooth extraction. Coadministration of IL-13 significantly restored bone formation compared with the anti–IL-33 monoclonal antibody + PBS group (Appendix Fig. 7). Collectively, these findings suggest that IL-33 signaling contributes to ILC2 activation and promotes bone formation in tooth extraction sockets, at least in part through IL-13–mediated mechanisms.
Discussion
Although IL-33–mediated ILC2 activation has been described in several tissues, its role in oral wound healing and alveolar bone regeneration has not been previously defined. In this study, we demonstrated that ILC2s play a critical role in bone regeneration within tooth extraction sockets by promoting osteoblastic activity through IL-13 secretion. Our findings suggest a biological mechanism in which gingival fibroblasts may act as upstream regulators during socket healing. Gingival fibroblasts produce IL-33 during mucosal repair, which may contribute to the activation of ILC2s and promotion of osteogenesis. Thus, ILC2s represent important mediators of postextraction alveolar bone regeneration and highlight a potential functional role of the oral mucosa in osteoimmunology.
We identified IL-13 as a key effector molecule produced by ILC2s that promotes bone regeneration within tooth extraction sockets. In addition to directly enhancing the osteogenic differentiation of BMSCs, as demonstrated by increased mineralization in vitro, IL-13 likely supports bone formation through multiple complementary mechanisms. ILC2-derived IL-13 suppresses receptor activator of nuclear factor kappa B ligand (RANKL)–induced osteoclast differentiation and bone resorption (Omata et al 2020). Moreover, IL-13 modulates macrophage polarization toward an anti-inflammatory M2 phenotype (Yin et al 2022), thereby supporting tissue repair. IL-13 has also been implicated in promoting angiogenesis (Zhang et al 2023), a process essential for establishing the vascular network underlying bone regeneration. Collectively, these previous studies suggest that ILC2-derived IL-13 may contribute to alveolar bone regeneration through multiple mechanisms, including the inhibition of osteoclast activity, suppression of inflammatory responses, and promotion of angiogenesis. In the present study, we observed that IL-13 inhibited osteoclast differentiation in vitro, suggesting a potential role in suppressing osteoclastogenesis. However, the effects of IL-13 on inflammatory regulation and angiogenesis were not directly evaluated, and the in vivo contribution of osteoclast inhibition in socket healing remains to be fully elucidated. Therefore, these mechanisms warrant further investigation in the context of extraction socket healing.
Beyond the initial inflammatory phase, our findings suggest that the IL-33–ILC2–IL-13 axis plays a multifaceted role in the subsequent stages of bone regeneration, including matrix formation, mineralization, and remodeling. In our model, IL-13 significantly enhanced the expression of key osteogenic markers such as Runx2, Sp7, and Col1a1, as well as matrix mineralization in BMSCs. This indicates that ILC2-derived IL-13 directly supports the synthesis of the organic bone matrix and its subsequent mineralization. Additionally, our in vitro experiments demonstrated that IL-13 inhibited osteoclast differentiation, suggesting a potential role in regulating bone remodeling. Consistent with prior literature, ILC2-derived IL-13 can potentially modulate the remodeling process by limiting osteoclastogenesis and maintaining a proanabolic environment that supports the stabilization of newly formed bone. However, since osteoclast activity and the early exudative response were not directly evaluated in vivo in the present study, these mechanisms remain to be fully clarified in the context of extraction socket healing. Future studies are required to delineate how the IL-33–ILC2 axis coordinates the transition from the early inflammatory exudate to functional bone remodeling.
Although IL-13–mediated osteogenic differentiation represents a primary mechanism by which ILC2s facilitate bone regeneration, additional cytokines secreted by ILC2s may also contribute to this process. Activated ILC2s produce IL-4 (Neill et al 2010), which enhances osteoblast differentiation (Kondo et al 2024b), and amphiregulin (Monticelli et al 2011), which activates transforming growth factor–β signaling pathways involved in osteogenesis (Wang et al 2025). ILC2-derived IL-5 promotes eosinophil activation and subsequently suppresses osteoclastogenesis (Omata et al 2022). In addition, ILC2s can regulate osteoclast differentiation through the production of IL-4 and granulocyte–macrophage colony-stimulating factor. Thus, while IL-13 plays a central role, ILC2s potentially orchestrate bone regeneration through a coordinated network of cytokines that collectively create a microenvironment conducive to bone formation within extraction sockets. Further studies are required to clarify the roles of ILC2-derived cytokines other than IL-13 in regulating bone metabolism.
IL-33 is a well-characterized alarmin cytokine that plays a crucial role in mucosal immunity, including within the oral cavity. It is primarily expressed by epithelial cells, fibroblasts, and endothelial cells in oral tissues and is released upon tissue damage to alert and activate local immune responses (Wang et al 2023). Under steady-state conditions, IL-33 is predominantly expressed by epithelial cells, whereas during acute inflammation, Thy-1.2- fibroblasts/stromal cells become major sources of IL-33 in peri-root tissues. Beyond exacerbating inflammation, IL-33 regulates immune responses by promoting M2 macrophage polarization and suppressing excessive neutrophil infiltration, thereby protecting against alveolar bone loss (Liu et al 2024). However, the cellular sources and roles of IL-33 in the oral mucosa, particularly during wound healing, have not been fully elucidated. Our finding that gingival fibroblasts produce IL-33 following tooth extraction and may contribute to ILC2 activation provides additional insight into the cellular sources of IL-33 in oral tissues and suggests a potential role for the oral mucosa in immune-mediated bone regeneration.
Socket healing is regulated by multiple immune cell populations. A recent study has reported that NKT cells regulate alveolar bone regeneration during socket healing (Lin et al 2024). In this study, NKT cells rapidly infiltrate extraction sites during the early phase of healing and inhibit osteogenic differentiation through CXCL2 secretion, thereby attenuating bone regeneration. In contrast, our findings indicate that ILC2s play a distinct role during the later stages of socket healing by promoting osteoblastic activity through IL-13 production. Thus, different immune cell populations regulate alveolar bone regeneration in a temporally coordinated manner, with NKT cells acting during the early inflammatory phase and ILC2s contributing to the subsequent regenerative phase. In addition to NKT cells and ILC2s, other immune cells are also known to contribute to bone regeneration. Future studies will therefore be necessary to further elucidate the coordinated roles of diverse immune cell subsets and to clarify the interactions between ILC2s and other immune cells during the regulation of alveolar bone healing.
This study has several limitations. Although we demonstrated that ILC2-derived IL-13 promotes the osteogenic differentiation of BMSCs, its effects on other cellular processes involved in socket healing, such as the modulation of immune populations, angiogenesis, and fibroblast activation, were not fully explored. In addition, the present study did not directly assess osteogenic or osteoclastic activities in vivo using histological markers. Therefore, the relative contributions of bone formation and bone resorption to the observed bone volume changes remain to be fully clarified. Future studies incorporating histological or immunofluorescence analyses of osteogenic and osteoclast markers will be important to more precisely characterize the cellular mechanisms underlying ILC2-mediated bone regeneration in extraction sockets. Furthermore, while our data identified gingival fibroblasts as a major source of IL-33 postextraction, the functional role of fibroblast-derived IL-33 in vivo was not directly tested. Future investigations using conditional knockout models, such as fibroblast-specific IL-33–conditional knockout mice, will be necessary to directly determine the impact of fibroblast-derived IL-33 on ILC2 activation and downstream cytokine production during socket healing. Another limitation of the present study is that only female mice were used for the in vivo experiments. Bone metabolism and bone regeneration can be influenced by sex hormones (Lerner 2006). Although all experimental groups of the same sex were compared under identical conditions, future studies including both male and female mice will be necessary to determine whether the IL-33–ILC2 axis regulates extraction socket healing in a sex-dependent manner. A further limitation of this study is that IL-33, IL-13, and anti–IL-33 monoclonal antibody were administered systemically via intravenous injection. Although our analyses demonstrated localized immune responses and cytokine expression within the extraction socket, systemic immune modulation cannot be completely excluded. Therefore, the observed effects on bone regeneration may reflect both local and systemic immune regulation. Future studies using local cytokine delivery approaches will be necessary to further distinguish the local versus systemic contributions of IL-33–ILC2 signaling during socket healing.
In conclusion, this study suggests that the gingival fibroblast–IL-33–ILC2–IL-13 axis may play an important role in regulating bone regeneration after tooth extraction. These findings advance our understanding regarding the immunological regulation of bone healing and provide a foundation for innovative therapeutic strategies to improve clinical outcomes in dentistry.
Author Contributions
Y. Sato, T. Kondo, contributed to conception and design, data acquisition, drafted and critically revised manuscript; K. Otake, S. Ambo, A. Ashry, K. Engkatanachai, contributed to conception and design, data acquisition, analysis, and interpretation, drafted and critically revised manuscript; H. Egusa, contributed to conception and design, data interpretation, drafted and critically revised the manuscript. All authors gave their final approval and agreed to be accountable for all aspects of the work.
Supplemental Material
sj-docx-1-jdr-10.1177_00220345261449746 – Supplemental material for Gingival Fibroblast-Driven Osteoimmunology via the IL-33–ILC2–IL-13 Axis
Supplemental material, sj-docx-1-jdr-10.1177_00220345261449746 for Gingival Fibroblast-Driven Osteoimmunology via the IL-33–ILC2–IL-13 Axis by Y. Sato, T. Kondo, K. Otake, S. Ambo, A. Ashry, K. Engkatanachai and H. Egusa in Journal of Dental Research
Supplemental Material
sj-pdf-2-jdr-10.1177_00220345261449746 – Supplemental material for Gingival Fibroblast-Driven Osteoimmunology via the IL-33–ILC2–IL-13 Axis
Supplemental material, sj-pdf-2-jdr-10.1177_00220345261449746 for Gingival Fibroblast-Driven Osteoimmunology via the IL-33–ILC2–IL-13 Axis by Y. Sato, T. Kondo, K. Otake, S. Ambo, A. Ashry, K. Engkatanachai and H. Egusa in Journal of Dental Research
Footnotes
Acknowledgements
The authors thank Professor Fumiki Katsuoka and the technical staff of the Department of Integrative Genomics, Tohoku Medical Megabank Organization, for their assistance with scRNA-seq.
A supplemental appendix to this article is available online.
This study was supported by Grants-in-Aid for Early-Career Scientists (23K16081, 25K20340, T.K.) from the Japan Society for the Promotion of Science.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Data Availability
We state where data supporting the results reported in this study can be found, including, where applicable, hyperlinks to publicly archived datasets analyzed or generated during the study, within the manuscript. scRNA-seq data are available at GEO: GSE308321 and GSE308322.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.