
Abstract
The design of many shelters for laboratory mice, particularly plastic shelters, is mainly driven by economic, ergonomic and hygiene considerations, with little focus on providing the animals with opportunities to express species-specific behaviours and control over their environment. Even in the presence of artificial shelters (plastic houses, tunnels), laboratory mice will readily burrow if given the opportunity. The main aim of this study was to gain a better understanding of the importance of access to sheltered space for laboratory mice. We pair-housed mice (BALB/c, SWISS and C57BL/6, male and female) in conventional laboratory cages that contained either (i) resources to build underground shelter (burrowing substrate) or (ii) artificial structures providing shelter (e.g. red plastic house). We assessed how mice used these resources, as well as their preferences when given a choice between different shelters. In addition, we assessed the insulating properties of different shelters. Mice spent most of their time underground in deep bedding and the least time inside the standard red plastic house, where they nested outside the house. They also showed a strong preference for the deep burrow cage, and the most relevant features of preferred shelters seem to be darkness and being underground. Nesting material alone was also less insulating, possibly contributing to an increased need for non-shivering thermogenesis.
Introduction
The housing of laboratory mice has not changed much for decades. They are conventionally housed in relatively barren cages with nesting material and sometimes a shelter, with the focus on the convenience of researchers and caretakers and not necessarily accounting for the animals’ preferences. However, conventional housing with limited resource diversity is associated with increased morbidity in animal models of stress-sensitive diseases and other signs of poor welfare. 1
Refinement of housing and husbandry practices has been an ongoing effort for several decades. Over time, housing has shifted from barren shoe-box cages to more enriched environments that include nesting material, shelters and other structural enrichment.2–4 At the same time, standard ambient temperatures in research facilities remain well below the mouse thermoneutral zone,5–7 which can influence metabolism and immune function.5,8,9 Providing mice with nesting material, which also fulfils the animals’ motivation to build nests, 10 is now mandatory in many countries11,12 and is a simple and biologically relevant method to alleviate cold stress. Nevertheless, even with nesting material and shelters, evidence shows that welfare can be further improved through the addition of supplementary resources. 1
Shelter is one of the most important resources for mice as it allows the animals to control their environment in several ways; from perceived safety and darkness, to behavioural thermoregulation and burrowing. In the wild, mice dig burrows where they nest and motivation for burrowing and sheltering in deep bedding has also been observed in laboratory settings.13–16 Despite evidence for high burrowing propensity of laboratory mice, commonly used shelters at research facilities consist of polycarbonate or cardboard houses, which provide the animals with a dark space. While evidence suggests that mice prefer paper-based shelters to plastic ones for nesting, 17 plastic houses remain widely used as they ensure visibility for easy daily checks.18,19 Published and anecdotal reports suggest, however, that mice rarely use such houses as shelters and often make a nest outside of them.20,21 These findings indicate that plastic shelters may not be fit-for-purpose. Environments that prevent natural behaviours, such as burrowing, can cause frustration and stress,22,23 lack of perceived safety can increase anxiety24,25 and the standard plastic mouse house may prevent the animals from minimising thermal discomfort thus increasing the need for non-shivering thermogenesis. 26 Therefore, the lack of appropriate shelter could be a source of chronic housing stress, affecting laboratory mice throughout their lives.
We conducted a series of studies to assess how much mice use different types of shelters, ranging from minimal enrichment (nesting material) 27 to what may be biologically more relevant (burrowing substrate).13,28 Our aim was to identify the characteristics of shelters that determine the animals’ preferences and that can be implemented into existing laboratory cages. Additionally, we aimed to assess how mice rank different types of shelters by conducting two-choice preference tests. Once we had established the preference for different shelter types, we also measured thermal properties of the nest site with different shelters. We hypothesised that mice prefer to be housed with underground shelters and that these are more insulating, as measured by the radiating temperature of the nest site.
Animals, material and methods
Animals and housing
We conducted a series of three studies assessing shelter use and preference under different conditions. In all studies, mice arrived at the facility from Janvier Laboratories, France. They were weighed on arrival, assigned individual IDs, marked on their lower back with a black non-toxic marker (Stoelting Co.), or a fur cut, and pair-housed in conventional Type III cages (Tecniplast) with sawdust (Puraflake, Labodia) in Study 1, or wood-chip bedding (Lignocel Safe) in Study 2, and nesting material (10g Sizzle nest, Datesand) until the experiment started. Animals were housed under 12:12 h reversed dark:light cycle, with lights off at 12:00 h, with ad libitum food (Kliba Nafag, #3430) and tap water. No special acclimation period was applied after arrival, as the animals were housed under standard husbandry conditions and left undisturbed except for routine weekly cage cleaning. All animals were cup- or tunnel-handled throughout the studies. Experiments were purely exploratory and the study was not powered to detect a prespecified effect size. Instead, we used the smallest number of mice that allowed complete counterbalancing of order effects, while at the same time providing sufficient heterogeneity to the study sample (multiple strains and both sexes) to assess external validity of the results.
Experimental design and data collection
Shelter use
For Studies 1a and 1b, 20 RjOrl:SWISS (SWISS) and 20 BALB/cJRj (BALB/c) mice of both sexes were used. Room temperature ranged between 21°C and 22°C and relative humidity between 43% and 65%. At 11 weeks of age (Study 1a), mice were housed with one of the following shelters to assess shelter use (Figure 1(a)): i) nesting material only (mice could use nesting material to build a cover, but no additional structural shelter was added), ii) a small house (red transparent triangular mouse house (Tecniplast)), iii) a large house (custom-made from red transparent acrylic, covering half of the cage area), iv) underground plastic tunnels (Ferplast, 8 FPI 4808), where mice could retreat, or v) a burrowing cage with deep bedding (15 cm high), where mice could freely burrow. All cages (i–v) were additionally provided with 10g of nesting material (Sizzle nest, Datesand) outside the shelter. The tunnel and burrowing cages were made by stacking two Type III cages, with four holes (6 cm diameter) in the floor area of the top cage. In the tunnel cage, the four holes led to plastic tunnels, with one tunnel extending along the longer side of the cage and connecting two entry/exit holes, while the other two holes led to shorter L-shaped tunnels with dead-ends (Figure 1(b)). In the burrowing cage, the four holes led into deep bedding (Puraflake, Labodia) mixed with straw (JR Farm Bergwiesenheu) to increase stability of burrows. Small quantities of straw were added to all cages to control for material and olfactory cues. In both tunnel and burrowing cages, the bottom cage was covered with red transparent PVC foil, so that the underground area was perceived dark by the mice but allowed visual inspection during daily checks (Figure 1(b)). All mice were housed with each of the five shelters for one week over five consecutive weeks. Shelter types and the order in which the shelters were provided was balanced across strain and sex. Due to aggression, two cages with male BALB/c mice were separated during Studies 1a and 1b and the four males were housed singly.

In the tunnel cage, mice often moved the bedding around and into the tunnels, therefore, the underground area was not always dark as the floor of the top cage was not fully covered by bedding. To test whether the use of underground tunnels was dependent on perceived darkness, we ran a follow-up study (Study 1b) with the same animals. Here, the bottom tunnel cage (outside of the tunnels) was filled with wood-chip bedding (Lignocel Safe), so that the underground tunnel areas remained dark even if mice moved the bedding in the top cage. Mice were housed with one of the three shelter types: i) a small house (red transparent triangular mouse house (Tecniplast)), ii) a dark tunnel cage, and iii) a burrowing cage with deep bedding for one week each, in a counterbalanced order, the same as in Study 1a.
Although the use of sawdust bedding in Study 1 allowed mice to make stable burrows, all mice nested underneath the top cage floor area, where they created a chamber, usually in the middle or to one side, but never deep in the bedding. Therefore, the aim of Study 2 was to test whether the animals’ use of underground shelter depends on the depth and type of the burrowing substrate. For Study 2, 12 C57BL/6NRj (C57BL/6) and 12 RjOrl:SWISS (SWISS) mice of both sexes were used. The animals arrived at the facility at four weeks of age, were pair-housed in Type III cages upon arrival and were first used in another study, where they were housed with a red transparent tunnel and a mouse house (Tecniplast). Between the completion of the previous study and the start of Study 2, the animals were housed with only nesting material for three weeks. The animals were six months old at the start of Study 2 and were housed with one of three shelters for one week. Shelter types used were: i) a small house (half of the animals were housed with the triangular Tecniplast mouse house and half with the round house from Bio-Serv, counterbalanced across sex and strain), ii) a burrowing cage with deep bedding, the same as in Studies 1a and 1b, and iii) a shallow burrowing cage, with bedding depth of 5cm (Figure 1(a)). All cages contained another commonly used type of bedding (wood-chip; Lignocel Safe) in which mice could dig but could not build stable burrows. Nesting material (Sizzle nest, Datesand) was provided in all cages, outside the shelter. Mice were housed in a room with ambient temperature ranging between 22°C and 28°C and relative humidity between 35% and 65%. In all studies, cages were recorded on Days 5 and 7 after cage change, for 24 h (Vivotek IR cameras).
Preference tests
Preference tests were conducted at the end of Studies 1a and 2, by connecting two cages with a clear polycarbonate tunnel (diameter 6 cm, length 7 cm; Figure 1(c)). In Study 1a, each connected cage contained one of the five shelters. There were 10 possible combinations of shelters and to counterbalance the shelter pairs across the two sexes and strains, only eight cages (i.e. pairs of mice) per strain were used for preference testing (n=16 cages in total). Each pair of mice was housed with five of the 10 possible shelter pairs over five weeks, ensuring that for each given week, each shelter pair was presented only once per strain, and there were no repetitions of pairs across the cages. In Study 2, all 12 cages were housed with all three shelter pairs over the three weeks, in a counterbalanced order, balanced across sex and strain. To control for side-bias, the two mice were always placed in either the left or the right cage, alternating at each cage change, while ensuring that they were not placed in the cage with the same type of shelter in consecutive weeks. Blinding was not possible as the housing conditions were obvious to the experimenters during video coding.
Data collection and analysis
Shelter use
For both shelter use and preference, mouse locations were determined from videos using Solomon Coder (Version 19.08.02). Videos were coded using scan-sampling with 10 min intervals during a 2 × 24 h period, yielding 288 scans per mouse. Locations coded for shelter use were ‘In shelter’ or ‘Outside shelter’. Although nesting material was available in all cages, our focus was the use of structural shelters and not general nest use. In cages without a structural shelter, however, the nesting material represented the only available form of cover, and therefore time in shelter was defined as time spent in the nest. In cages that contained a structural shelter, time in shelter was defined as time spent inside that structure, even if a nest was built outside of the shelter. We did not quantify time spent in the nest separately in those cases, because our goal was to assess whether mice used the provided structural shelter. This approach allowed us to directly compare conditions with and without a structural shelter, using a consistent reference point for ‘time spent in the main source of cover’ under each housing condition.
Ten per cent of all videos were re-coded by both coders (AB and JN for Study 1; ML and JN for Study 2). In addition, 5% of videos were re-coded by each coder for intra-observer reliability. Both intra- and inter-observer reliability were high, with Cronbach’s alpha of 0.96 and 0.98, respectively, for Study 1 and 0.90 and 0.88 for Study 2.
Statistical analysis was carried out using R Studio (Version 2023.06.2). For shelter use, time spent in shelter (relative to observed time) was our variable of interest. We used a generalised linear mixed effects model (glmer) followed by post hoc tests with Bonferroni adjustments (phia) to analyse the data. ‘Sex’, ‘Strain’ and ‘Shelter’ were included as fixed factors and ‘MouseID’ and ‘Day’ nested within ‘Cage’ as random factors. Strain and sex effects were ignored in the post hoc tests. Data for the dark and light phases were analysed separately. No specific inclusion and exclusion criteria were set and all animals were included in the study. Four males were singly housed because of aggression; however, they were retained in the dataset and treated the same as group-housed animals for statistical analysis. Model assumptions were assessed through visual inspection of diagnostic plots.
Preference tests
The coding was done by scan-sampling every 10 min, with instantaneous recording of a period of 24h, yielding 144 scans per mouse (Study 1) and every 2 min during the 12h light period only, yielding 360 scans per mouse (Study 2). ‘Time in cage’ was coded for each mouse in Study 1, and ‘Time in cage’ and ‘Time in shelter’ in Study 2. Shelters were ranked by calculating a half-matrix, using a generalised linear mixed effect model (glmer), with ‘Sex’, ‘Strain’ and ‘Shelter pair’ included as fixed factors and ‘MouseID’ nested within ‘Cage’ as a random factor. Proportions of time spent in each cage (Study 1) and shelter (Study 2) for different cage pairs were calculated using the logarithm of the estimate for each cage/shelter pair. Because shelter use on Day 1 and Day 2 was strongly correlated after controlling for cage effects (partial correlation between Day 1 and Day 2 shelter use, controlling for cage, dark phase: r = 0.67, p < 0.001, light phase: r = 0.86, p < 0.001), only one day was recorded for cage or shelter preference coding (Day 5 after cage change). In Study 1a, preference was measured based on time inside either of the two cages, rather than time inside the shelters, to facilitate outcome assessment. However, mice often made the nest outside the shelter. Because mice spent a significant amount of time in the nest, either inside or outside the shelter, we used nest position noted during cage changes as a proxy to estimate shelter use in the preference test in Study 1a.
Radiated temperature of the nest site
In Study 2, the cages were recorded with a thermal camera (FLIR T530) one day before each cage change, to assess radiated temperature of the nest site. Thermal images were taken during the light phase when the mice were inside the nest. The cage was removed from the rack, placed on a cart, the cage lid was taken off and a 10s video focusing on the nest site was taken. The position of the thermal camera was adjusted for each cage type so that the distance between the camera and the cage floor was 60 cm. We used the image analysis software FLIR Tools (Version 5.13.18031.2002) to measure the average temperature (°C) of the circular area around the centre of the nest site (5 cm diameter). Thermal images were adjusted for camera distance, ambient temperature and humidity. Emissivity was set to 0.92. 29 The mean reflected temperature of the nest site per cage was analysed using lm with ‘Sex’, ‘Strain’ and ‘Shelter’ as fixed factors and ‘Cage’ and ‘Week’ as random factors, followed by post hoc tests with Bonferroni adjustments. Model assumptions were assessed through visual inspection of diagnostic plots.
Results
Shelter use
In all cages, mice made nests either inside or outside a shelter. Shelter use varied between shelter types during both dark and light phases (Study 1a: F4,354 = 223.87, p < 0.001 and F4,333 = 307.39, p < 0.001, respectively; Figure 2), but the overall pattern of use was similar in both phases. Mice spent similar amounts of time inside the nest when only nesting material was available, inside the large house, and underground in the burrowing cage, while very little time was spent inside the tunnels or the small house.

Time spent (relative (rel) to observed time) in different types of shelter in Study 1a, Study 1b and Study 2.
In the follow-up study (Study 1b), with darker tunnels, mice again spent most of their time in the burrows during both phases (dark phase: F2,174 = 92.70, p < 0.001; light phase: F2,174 = 188.45, p < 0.001). While time spent in tunnels increased compared with Study 1a, dwelling time was still lower than time spent in the burrowing cage and very little time was spent in the small house.
In Study 2, mice were housed with wood-chip bedding where they could not make stable burrows. Time spent in shelter again differed between shelter types in both the dark and light phases (F2,125 = 18.41, p < 0.001 and F2,125 = 91.91, p < 0.001, respectively; Figure 2). Post hoc tests revealed that time spent underground was similar between deep and shallow burrowing cages, while time spent in the small house was much lower (see Supplemental material Table S1a online for statistics).
Shelter use was similar for males and females across all three studies, particularly for the small house (pairwise comparisons with Bonferroni correction; Study 1a: estimate = −0.018, p = 0.72, Study 1b: estimate = 0.014, p = 0.81, Study 2: estimate = −0.043, p = 0.73), and deep burrow (Study 1a: estimate = 0.05, p = 0.342, Study 1b: estimate = 0.06, p = 0.45, Study 2: estimate = 0.03, p = 0.81), which were included in all three experiments (Supplemental Figure S1). More males than females used the tunnels (Study 1a: estimate = 0.244, p < 0.001), and when the tunnels were made dark in Study 1b, tunnel use increased in both males and females, although some females still nested outside the tunnels (Study 1b: estimate = 0.254, p = 0.003). SWISS mice spent less time of the light phase in the small house compared with BALB/c (Study 1b: estimate = 0.34, p < 0.001) and C57BL/6 mice (Study 2: estimate = 0.29, p = 0.03), while time in deep burrows did not differ between strains (Study 1a: estimate = −0.05, p = 0.29, Study 1b: estimate = −0.05, p = 0.55, Study 2: estimate = 0.05, p = 0.66; Supplemental Figure S2; see Supplemental Table S1b for details). Estimates represent absolute differences in the proportion of observation time spent in each shelter.
Shelter preference
In Study 1a, shelters were ranked based on the time spent in either of the two connected cages. Mice preferred the deep burrow cage, spending at least 80% of their time in the cage, followed by the large house, small house and the tunnel cage, while the cage with only nesting material was least preferred (Figure 3(a) and Supplemental Table S2a). The burrowing cage was the only shelter where all mice nested inside the shelter, whereas in all other cages, up to 65% of cages had nests outside the shelter (Figure 3(b)).
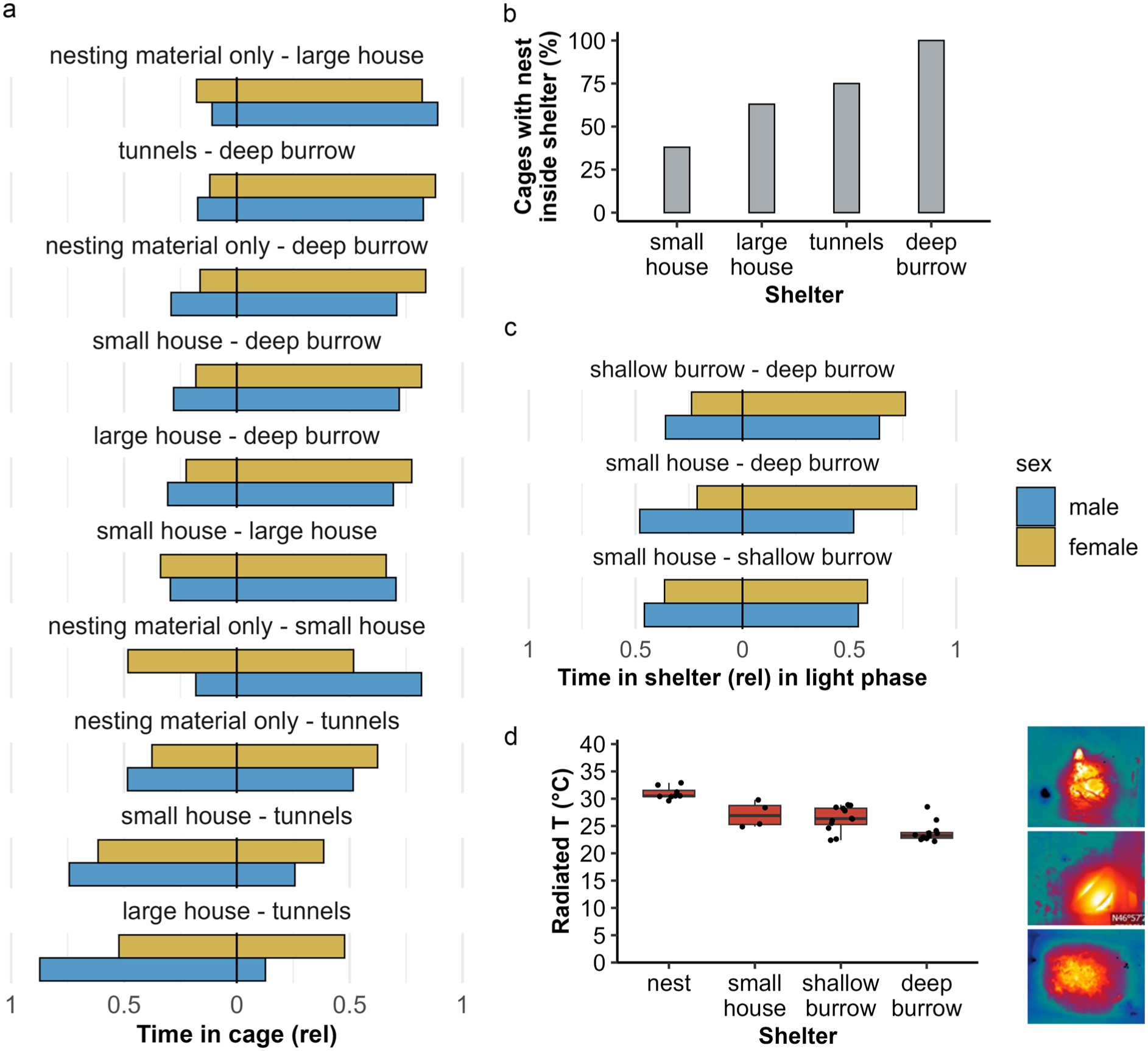
(a) Time spent in cage across 24h in Study 1a, relative (rel) to observed time, in each connected cage during preference testing. (b) Relative number of cages with the nest site inside shelter as recorded during cage changes in Study 1a. (c) Time spent in shelter during the light phase in Study 2 during preference testing, relative to time in both shelters. (d) Radiated temperature (T) of the nest site in Study 2. Thermal images show radiated temperature from a nest, a mouse house and deep burrow.
In Study 2, shelters were ranked based on time spent inside shelters in either of the two connected cages, relative to all shelter time. Here, the most preferred shelter was the deep burrow, where mice spent up to 76% of their time, followed by shallow burrows, and the least preferred shelter was the small house. Preference for deep and shallow burrows was more pronounced in females (Figure 3(c) and Supplemental Table S2b).
Radiated temperature of the nest site
Radiated temperature of the nest site differed between all shelter types (F3,21 = 43.40, p < 0.001) except small house and shallow burrow (pairwise comparisons with Bonferroni correction; estimate = −0.92, p = 1.000; Figure 3(d)). Mean radiated temperature of the nest site with only nesting material was higher (31°C ± 0.4°C) compared with when the nest was inside the shelter (ranging from 23.8°C ± 0.5°C for deep burrows to 27.1°C ± 1.2°C for the small house). This analysis included cages where the mice were sheltering inside the small house and cages where the nest site was outside the house (nesting material; Figure 3(d)).
Discussion
Our results show that when given a choice, laboratory mice spend most of their time in underground shelters, either artificial or self-made burrows, and this effect was seen in outbred and inbred strains with different propensities for burrowing, 14 both sexes and mice of different ages. Rodents prefer dark areas, 30 actively burrow for various reasons13,31–33 and even slightly deeper bedding triggers digging behaviour.34,35 Motivation and preference for burrowing in laboratory mice have been confirmed before,13–16 and burrowing behaviour has been used as an indicator of well-being and motor function;31,36 however, no studies have yet attempted to determine which aspects of burrows are important for mice and whether they prefer to dwell in self-made or artificial burrows. We find that mice used all underground shelters when made dark; however, neither stability of burrows nor bedding depth affected their use, as mice made nests directly beneath the floor of the top cage. Together with the increased preference for the large house compared with the small house, our data thus suggest that the tactile stimulus of being underground might be necessary to trigger retreat behaviour in mice, and these properties might not be sufficiently provided by conventional shelters. This finding is not surprising, as specific shelter shapes have also been shown to be important for gerbils. 32 However, we cannot completely ignore the fact that underground shelters still allowed mice to actively dig, as even in cages with pre-constructed tunnels, mice moved bedding into the tunnels. It is possible that retreating to tunnels containing bedding as well as underground shelters with wood-chip bedding were sufficient to fulfil the motivation to burrow. Both burrowing and retreating to a safe place are species-specific behaviours in mice31,32 and moving away from the exposed open space of a standard cage into an underground area could additionally enhance the animals’ perception of safety.
Interestingly, we found that mice spent very little time in plastic mouse houses, and instead built their nests outside them. Small houses (either polycarbonate or paper) are widely used as enrichment at research facilities 3 and have been reported to improve welfare in rats, 37 increase longevity and reduce wounding in BALB/c males 38 and affect motor behaviour and learning. 39 However, previous studies indicate that mice often prefer cardboard houses to plastic ones because they allow greater environmental control. 17 Poor use of red plastic houses has been reported for several mouse strains,17,21,40,41 a finding also observed in our study, suggesting that standard plastic houses might not adequately meet mice’s shelter preferences. There was some variability in tunnel and house use, which likely reflects individual differences in how mice interacted with these shelters rather than sex- or strain-specific effects. Some animals used the tunnels or houses consistently for resting and nesting, while others made the nest outside or used them only intermittently.
Beyond measuring the use of shelters when only one shelter was available, we also assessed shelter preference using pairwise preference tests. Tests using connected cages are commonly employed to detect preferences for resources, such as bedding, nesting and shelter types.42–44 In line with shelter use results, the small house and nesting material alone were also the least preferred shelters, while both shallow and deep burrowing cages were preferred. However, the preference for burrowing cages was much more pronounced in females, possibly due to territorial behaviour in males, as some male pairs used both connected cages. Male mice show preference for social contact, 45 but also demonstrate a tendency to establish separate territories46–49 when opportunities arise.
While digging behaviour or specific properties of underground shelters might satisfy motivation for burrowing and sheltering, 32 another important benefit could be better thermoregulation. At research facilities, mice are housed at temperatures well below their preferred range of 26–30°C.5,7,26 Nesting material can alleviate some degree of cold stress;26,50 however it might not be enough,51,52 and while group-housed mice can reduce cold stress to some extent by huddling, male mice are often housed singly. 49 To prevent hypothermia, mice rely on non-shivering thermogenesis by utilising brown adipose tissue.53,54 Therefore, improved insulation of the nest site could alleviate cold stress, which can affect the immune system and metabolism.53,55,56 We found that all shelter types, including the small house, insulated the nest site, while the radiated temperature from the nest site with only the nesting material was higher. This suggests that the insulating properties of nesting material alone are much lower than those of artificial shelters and while a small house might be sufficient for reducing heat loss, its provision as a shelter is meaningless if mice do not use it. Heat dispersal from the heat source can differ between shelters of varying size and volume. Because the shape and size of underground shelters in our study varied between cages, it was not feasible to determine their exact volumes. The 5 cm diameter circular area was chosen to standardize measurements across cages and to capture the region directly above the nest, where the animals were located at the time of recording and heat emission was most consistent. This provided comparable estimates of surface temperature above the primary heat source rather than integrating total radiated heat, which would depend strongly on shelter geometry. A larger shelter, however, might display a lower surface temperature simply because the same amount of heat is distributed over a larger area, not necessarily because it provides better insulation. Thus, our measure serves as a relative indicator of nest-site temperature rather than a direct quantification of total heat retention and provides preliminary data requiring further investigation. Nonetheless, mice continued to use deep burrows even when ambient temperature reached 28°C during the light phase (Study 2), indicating that factors beyond behavioural thermoregulation contribute to the use of deep bedding.
Housing conditions have been identified as the factor that most strongly influences the welfare of laboratory rodents. 57 Besides cold stress, the physiological effects of which have been well researched, 53 psychological stress from thwarted motivation for digging or retreat can also negatively impact well-being.58–60 Further research is needed to evaluate the impact of underground shelters on behaviour and physiological functioning. However, given that mice are the most used animals in research, and poor health and well-being might compromise the validity of the research, adequate shelter may have a substantial impact on both animal welfare and the scientific validity of mouse research.61,62
Supplemental Material
sj-docx-1-lan-10.1177_00236772251404062 – Supplemental material for A house is not a home: Shelter preferences of laboratory mice
Supplemental material, sj-docx-1-lan-10.1177_00236772251404062 for A house is not a home: Shelter preferences of laboratory mice by Alina Bonorand, Marie Leekens, Bernhard Voelkl, Hanno Würbel and Janja Novak in Laboratory Animals
Supplemental Material
sj-tif-2-lan-10.1177_00236772251404062 – Supplemental material for A house is not a home: Shelter preferences of laboratory mice
Supplemental material, sj-tif-2-lan-10.1177_00236772251404062 for A house is not a home: Shelter preferences of laboratory mice by Alina Bonorand, Marie Leekens, Bernhard Voelkl, Hanno Würbel and Janja Novak in Laboratory Animals
Supplemental Material
sj-tif-3-lan-10.1177_00236772251404062 – Supplemental material for A house is not a home: Shelter preferences of laboratory mice
Supplemental material, sj-tif-3-lan-10.1177_00236772251404062 for A house is not a home: Shelter preferences of laboratory mice by Alina Bonorand, Marie Leekens, Bernhard Voelkl, Hanno Würbel and Janja Novak in Laboratory Animals
Footnotes
Acknowledgements
We would like to thank the animal care takers, Myriam Siffert, Fabiana Kölliker and Georgina Hayoz, for their help with husbandry and animal care. Thanks to Michelle Gygax, Veit Cooper and Andreas Glarner for making the burrowing and tunnel cages.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study was funded by the Swiss Federal Food Safety and Veterinary Office (FSVO; Contract number 714001679).
Ethical considerations
This study was carried out in accordance with guidelines of the Swiss Animal Welfare Ordinance (TschV 455.1). It was approved by the Cantonal Veterinary Office in Bern, Switzerland (permit numbers BE 5/21 and BE 1/22). The reporting of the study follows the ARRIVE 2.0 guidelines for reporting animal research. 63
Supplemental material
Supplemental material for this article is available online.