
Abstract
Although overeating is the root cause of many diseases and even premature death in laboratory animals, ad libitum feeding is still the prevailing practice in rodent husbandry. There is a need for practical methods of food restriction compatible with group housing. The diet board was developed as a mechanical solution to food restriction of group-housed rats. It tightly embeds food pellets in grooves cut into aspen boards, forcing the rats to gnaw wood to reach their food. In this two-year experiment, 146 nine-week-old outbred Hsd:Sprague Dawley® rats were divided into four equal groups: diet board-fed males and females, and ad libitum-fed males and females. Blood samples for clinical chemistry were collected at regular intervals during the study, and times of spontaneous death or euthanasia were recorded. Triglyceride and cholesterol values were significantly lower in diet board males than ad libitum males. The restriction of food intake via a diet board significantly increased the 24-month survival rate for both female and male groups. The better observed survival rates amongst diet board-fed rats makes it possible to start long-term studies with far fewer rats than with ad libitum feeding. Thus, using a diet board appears to be a promising method for the reduction of animal numbers in long-term rat studies that is compatible with group housing.
Introduction
Compared with restricted feeding regimens, ad libitum feeding, where rats have constant access to food, typically results in an increased incidence of premature death in long-term studies. 1 Yet ad libitum feeding is still the prevailing practice in rat husbandry, with no change in sight.
In rats, studies have shown that dietary restriction reduces overweight, improves health and extends their lifespans, the effects being relative to the extent of the restriction.2–4 In mild to moderate dietary restriction (up to 25% reduction from the ad libitum level), the beneficial effects are achieved without substantial drawbacks to animal welfare or experiments.5,6 With more severe dietary restriction, animals are likely to suffer from hunger, and their growth might be retarded. Severe dietary restriction might even hinder the disease processes under investigation, for example, tumour formation in long-term carcinogenicity studies.7,8
Obesity can lead to an increased use of animals in studies in two ways. First, overfeeding-related metabolic problems and background diseases might add result variation, leading to an exponential increase in the number of animals required for the same study outcome. This has been demonstrated with organ weights but not with other variables.9–12
Second, in long-term studies, obesity can result in poor survival rates unrelated to the actual procedure or disease being studied. Rodents have been used in most chronic toxicity studies and in almost all carcinogenicity testing. The Organisation for Economic Co-operation and Development guideline states that for a negative result to be acceptable in a rat carcinogenicity bioassay, survival in the study should not be under 50% in any group at 24 months, when rats are 25–26 months old. 13 Reaching this survival rate is challenging with ad libitum feeding. Thus, any animal husbandry practice that significantly enhances longevity possesses tremendous potential for reduction in animal numbers.
Dietary restriction is commonly carried out by giving the animals a measured amount of food once or twice a day. A major reason this practice is not widely adopted is the high workload, which would be unfeasible in large animal facilities. Another hurdle is the European regulatory requirement to group-house social animal species unless there are welfare or scientific reasons not to. 14 Limited feeding of group-housed animals might lead to unequal food intake among cage-mates due to group hierarchy, with a risk of undernourishment of subordinate individuals. A dietary restriction method that allows continuous social housing would reduce the number of animals required, the personnel workload and the financial costs. The ideal situation would be to reach these objectives with as mild a dietary restriction level as possible to avoid malnutrition.
The diet board was developed for the feeding of group-housed rats. The diet board consists of grooved aspen boards embedded with food pellets, which forces rats to gnaw wood in order to eat. They can choose when and how much they eat, so that it is, in effect, very laborious ad libitum eating. Working for food restricts the rats’ food intake and increases their energy expenditure, thereby limiting the development of obesity.15,16
The effects of dietary restriction on rat physiology have been documented in numerous studies, but it remained unclear whether the effects of diet board feeding would be similar. The feasibility and safety of the diet board had earlier been assessed in a 10-week experiment with young male Wistar rats.15,17 Additionally, the effects of diet board feeding over two years on male and female Sprague Dawley rats’ food and water intake, growth and adiposity, as well as behaviour, stress and welfare have already been published by us elsewhere.5,16 The objective of this study was to compare the clinical chemistry values and survival of diet board-fed and ad libitum-fed Sprague Dawley rats in the above-mentioned two-year experiment and assess the consequent reduction potential of diet board feeding.
Animals, material and methods
The study protocol was approved by the National Animal Experiment Board of Finland (licence ESLH-2008-03667/Ym-23) and the animal care and procedures complied with EU and Finnish legislation.14,18
Animals and housing
The study used 146 outbred Hsd:Sprague Dawley® male and female rats. These were born at the Oulu Laboratory Animal Centre, University of Oulu, Finland and were nine weeks old at the start of the study. The initial mean (SD) body weight was 303 (24) g in males and 202 (11) g in females. Health monitoring was done according to FELASA (Federation of European Laboratory Animal Science Associations) recommendations, with the only finding being serum antibodies of the rat minute virus. 19
Two weeks prior to starting the experiment, the rats were moved from the breeding barrier to the research unit, ear-marked and regrouped into three rats per cage. During the experiment, if two of the three rats in a cage died, a companion rat for the remaining one (not included in the analyses) was added to the cage.
The cages were solid bottom type IV (59.5 cm× 38 cm× 20 cm) polycarbonate cages with wire lids (Tecniplast 1354G, Buguggiate, Italy). Aspen shavings (PM90L) were supplied as nesting material and aspen chips (4HP) as bedding (Tapvei, Kiili, Estonia). Untreated municipal tap water was continuously available in polycarbonate bottles. Cages and bottles were changed twice a week between 09:00h and 10:00h.
Room temperature was 21 ± 1°C, relative humidity 40–60%, ventilation rate 15 ACH and illumination 350 lx at 1 m height. Full fluorescent tube lighting was on from 07:00h to 18:00h, with lights turned on gradually from 06:00h to 07:00h and dimmed from 18:00h to 19:00h.
Feeding
Autoclaved natural ingredient food (Lab For R 36, Lantmännen, Kimstad, Sweden) was available continuously either in diet boards (experimental groups) or ad libitum in cage lid hoppers (control groups). The ingredients were wheat, barley, soy, potato and vegetable oil, with added vitamins, minerals and trace elements. The food contained 18.5% crude protein, 4.0% crude fat (of which 1.0% linolic acid), 3.5% vegetable fibre, 55.7%, nitrogen-free extract, 6.3% ash, maximum 12% water and 1200 kJ/100 g metabolizable energy.
Filled diet boards, control group food and control cage boards were autoclaved simultaneously (45 min, 121°C). Both ad libitum food and diet boards were changed once a week at cage change. The diet boards and control cage boards were turned upside down on the fourth day of the week to make access to what remained in the boards easier.
The diet board
Two 2.7 cm× 14 cm× 36 cm aspen boards (Tapvei, Kortteinen, Finland) were joined in the middle to form a cross-shaped wall structure that divided the cage into four semi-closed compartments (Figure 1(a)). Each board contained 10 vertical grooves (diameter 12 mm, 6 mm wide gap) parallel to the 14 cm edge on both sides, filled with tightly-fitting food pellets (Figure 1(b)). Diet board rats received food only from the boards, which were always in the cage. The 6 cm× 6 cm rectangular passages at the ends of the boards and the space between the diet board and the lid allowed the rats to move throughout the whole cage. Control cages were supplied with similarly structured plain boards without grooves or food pellets. The diet boards and plain boards served as the mandatory cage complexity items. 14

(a) Diet board in use; (b) half of the board, illustrating the structure.
Study design, randomisation and blinding
The study groups were diet board males and females (experimental groups), and ad libitum males and females (control groups). Four 36-rat birth cohorts entered the study at one- to two-month intervals. Birth cohorts comprised five litters each: four litters of four males and four females, and one litter of two males and two females. Littermates were divided randomly but evenly into diet board and ad libitum groups. Within the study groups, rats were randomised into the cages, separating siblings from each other. Randomisation was achieved by grouping the rats’ ID numbers using the Excel random number function. Each cohort was housed in its own cage rack, with groups divided evenly on the shelves.
For inclusion in the experiment, rats had to be clinically healthy and outwardly normal. There were no pre-set exclusion criteria.
Sample size
The experimental unit for quantitative endpoints was the cage (absolute values) or the rat (variability). This study included both qualitative and quantitative endpoints, but the qualitative endpoint (survival) requires higher numbers of animals and is thus decisive. The total number of rats required to detect a 35% change in survival at significance level p = 0.05 and statistical power 0.8 was 66. 20 Since males and females were analysed separately, the total number required was thus 132. An additional 10% safety margin was added to account for losing animals or samples for reasons unrelated to the experimental setup, making the original sample size 144. An extra cage of diet board males was added after excluding a diet board male because of an injury just two weeks after the experiment began, making the final sample size 146 (38 diet board males, 36 rats in other groups).
Blood sampling and clinical chemistry analysis
Blood samples were collected 0, 3, 6, 12, 18 and 24 months from the study start. The respective ages of the rats were 9, 21, 33–34, 60–64, 85–87 and 113–114 weeks old. Cohorts 1 and 2 were sampled from Monday to Wednesday, cohorts 3 and 4 from Tuesday to Thursday, all between 13:00 and 15:00h. Rats were not fasted before sampling.
Cages were brought in random order into the operation room, one at a time. Rats were pre-warmed in a 40 ± 2°C chamber for 2 min to dilate the tail veins and then immobilised in a transparent acrylic Tailveiner tube (Braintree scientific, Inc., Braintree, MA, USA). Blood was sampled with a 20 G × 1½" vacuum needle (Terumo Venoject, Leuven, Belgium) with the rubber sheath removed to let the blood drip freely into the tube (Terumo Venosafe 3.5 ml, silicone coated, with gel plug, Leuven, Belgium). Glucose was measured from the first drops with a quick-test (Accu-Check Aviva, Roche Diagnostics GmbH, Mannheim, Germany). The aim was to collect 1–2 ml of blood. Occasionally this failed, some samples being too small for a complete serum panel.
Blood tubes stood at room temperature for 30–120 min before centrifuging (3000 rev/min, 10 min). Serums were stored in vials (Sarstedt Micro tube 1.5 ml, Nümbrecht, Germany) at ⩽ −25°C. Samples were transported in an EPS box with dry ice to the laboratory (Orion pharmaceutical company, Espoo, Finland) for analysis. Clinical chemistry values were determined using an ADVIA 1650 Chemistry Analyzer (Siemens, Germany).
The clinical chemistry variables assayed were K (potassium), Na (sodium), Cl (chloride), ALAT (alanine transaminase), AFOS (alkaline phosphatase), ASAT (aspartate transaminase), crea (creatinine), urea, bil (bilirubin), prot (protein), alb (albumin), glob (globulin), alb/glob, glu (glucose), chol (cholesterol), D-HDL (direct high-density lipoprotein), LDL (low-density lipoprotein), trig (triglyceride), CK (creatine kinase), Ca (calcium) and Pi (phosphate).
Survival analysis
Times of spontaneous death or euthanasia were recorded and survival times calculated. The study length was two years, and the rats alive at termination of 730 days were euthanised.
Health, welfare and humane endpoints
Health problems were documented and a clinical examination was performed if needed. Study-specific humane endpoints included tooth problems and excessive reduction or inadequate increase in body weight with age-specific weight limits, as described earlier. 16 The maximum average diameter of superficial tumours was set at 25 mm. Any other welfare disturbance that could not be quickly ameliorated was an indication for euthanasia.
Rats were euthanised by heart puncture for blood collection under isoflurane anaesthesia, and death was ensured with carbon dioxide in a pre-filled chamber.
Statistical analysis
The data were analysed using IBM® SPSS® Statistics 20 (IBM Corporation, Somers, NY, USA).
The main outcome variables were clinical chemistry values and time to death. The main explanatory variable was feeding method. Males and females were analysed separately.
Repeated measurements were examined using multivariable linear regression analysis with generalised estimating equations (GEEs). Time point was designated as a within-subject factor. 21 Birth cohort was included in the models for adjustment. Repeated measures time points were 0, 3, 6, 12, 18 and 24 months from the study start.
Mean clinical chemistry values for each cage were compared between groups by a repeated measures analysis.
Absolute and relative within-group variations of the clinical chemistry values of individual rats were compared between feeding groups. At each time point, the deviation of every original datum from the study group median was calculated to illustrate the absolute variation. For relative variation, this deviation was divided by the group median. Absolute and relative deviations from the group median were compared between feeding groups using multivariable linear regression analysis with GEEs.
The two-year survival rates of feeding groups were evaluated with the chi-square test. Kaplan–Meier survival analysis was used to obtain the estimated survival curves over the two-year study period. The statistical significance of the difference in survival curves was evaluated using the log rank test.
In the parametric tests, the normality of residuals was evaluated graphically with histograms and normality plots.
p-values < 0.05 were considered statistically significant. In clinical chemistry analyses with multiple comparisons, statistical significances were adjusted using the Bonferroni method.
A sensitivity analysis was performed by conducting significance tests with and without extreme outliers. The results of cases in which findings did not change are presented including the outliers.
Results
Blood and serum clinical chemistry values
Trig and chol values were statistically significantly lower in diet board than in ad libitum males (Figure 2). Chol: model-based effect −0.29 mmol/l, 95% confidence interval (CI) (−0.44, −0.14), p < 0.001, Bonferroni-adjusted p < 0.04. Trig: model-based effect of diet board feeding −0.37 mmol/l, 95% CI (−0.46, −0.28), p < 0.001, Bonferroni-adjusted p < 0.04.
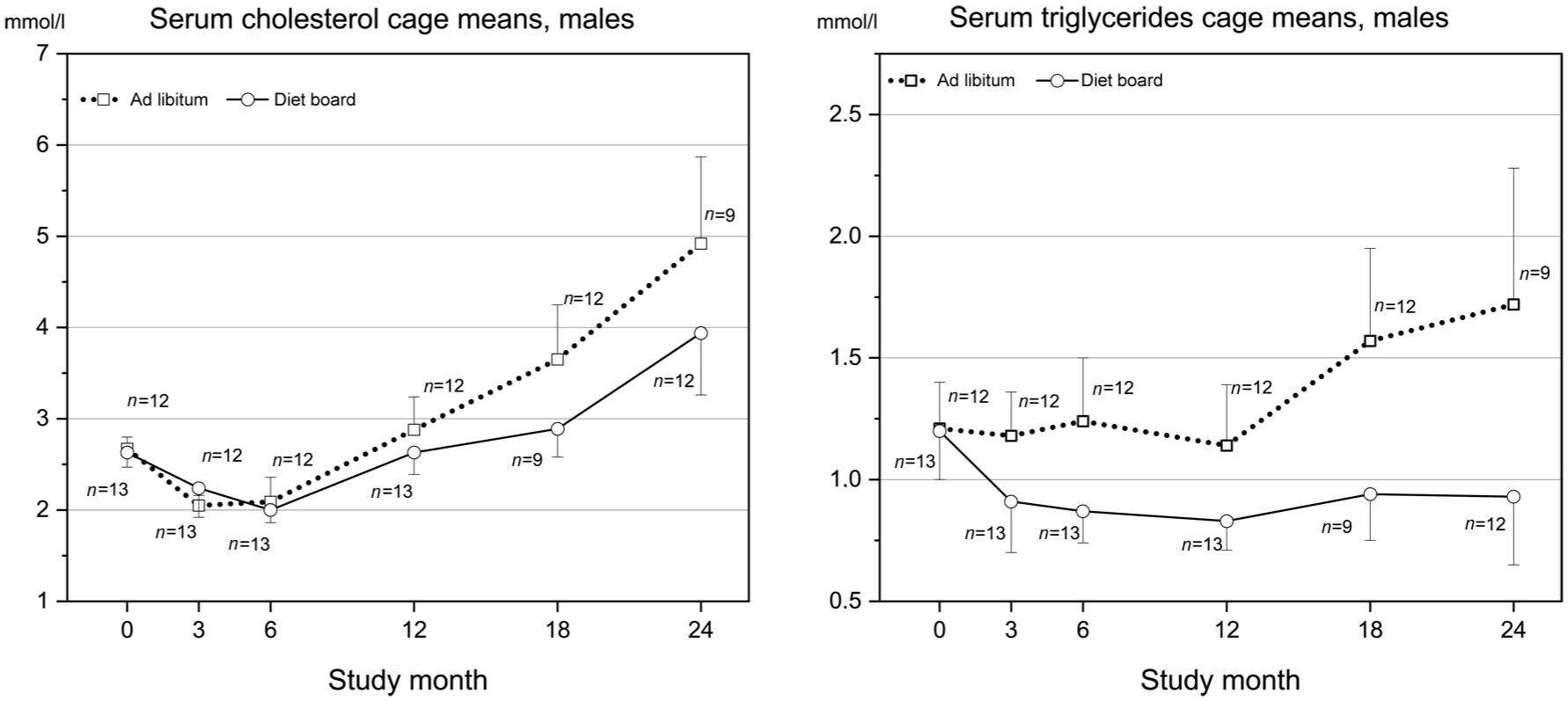
Serum cholesterol and triglyceride cage mean (SD) values of males. Both triglyceride and cholesterol values were significantly lower in the DB group than in the AL group (p < 0.001 and Bonferroni-adjusted p < 0.04 for both). The ages of the rats at the blood sampling time points were 9, 21, 33–34, 60–64, 85–87 and 113–114 weeks-old.
There were no statistically significant differences in Cl levels between feeding groups in either sex in the whole dataset, but when extreme outliers of individual rats’ results were removed, Cl values were higher in diet board than in ad libitum males. Whole dataset: model-based effect 1.31 mmol/l, 95% CI (0.39, 2.22), p = 0.005, Bonferroni-adjusted p = 0.2. Analysis without extreme outliers: model-based effect 1.51 mmol/l, 95% CI (0.70, 2.32), p < 0.001, Bonferroni-adjusted p < 0.04.
There were no other statistically significant differences between the feeding groups for either sex.
Variability
There were no statistically significant differences between feeding groups in either sex, in either absolute or relative variability of the clinical chemistry results.
Survival
The number of rats that survived to the end of the experiment is presented in Table 1. The difference between feeding groups in the two-year survival rates was statistically significant in the chi-square (χ2) test in the overall comparison (χ2 (1) = 15.03, p < 0.001) and separately in both males (χ2 (1) = 7.74, p = 0.010) and females (χ2 (1) = 8.13, p = 0.009). The odds ratio of surviving to the end of the experiment with diet board feeding as compared with ad libitum feeding was 3.87 overall, 3.86 for males and 4.96 for females.
Survival of rats at 24 months.
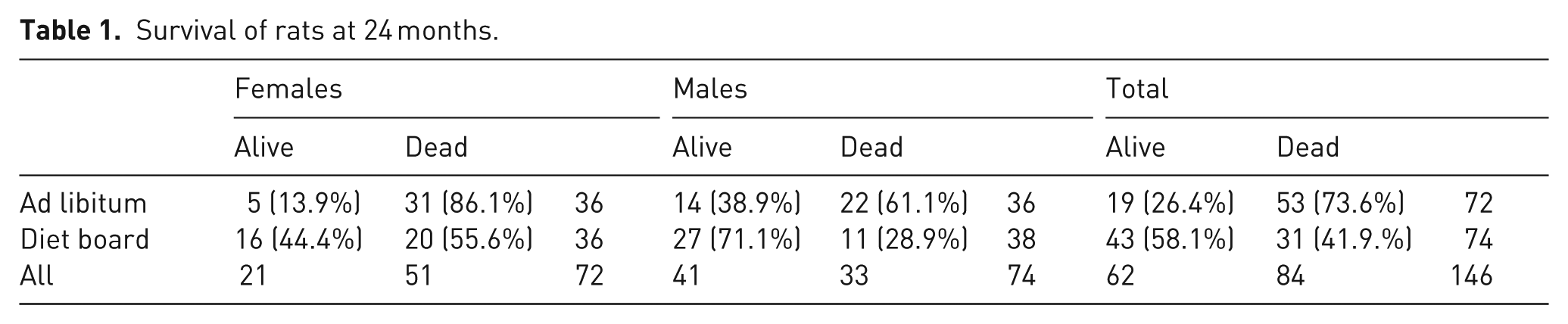
In the Kaplan–Meier log rank test, which weights all time points equally, the survival distribution was statistically significantly higher for diet board than for ad libitum-fed rats in both the overall comparison and separately for both males and females (Figure 3).
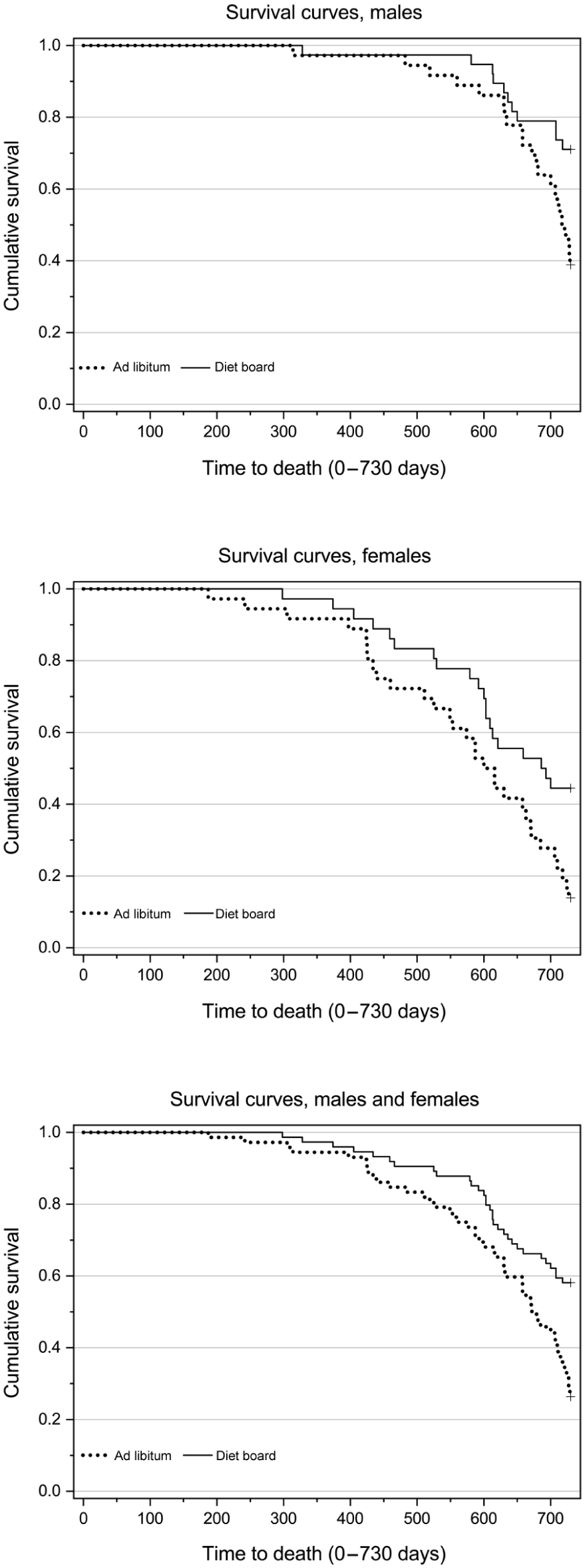
Cumulative survival in males (N = 74), females (N = 72) and both sexes combined (N = 146). Log rank test of Kaplan–Meier survival analysis gave statistically significant difference between the feeding methods in the overall comparison (χ2 (1) = 12.14, p < 0.001) and separately in both males (χ2 (1) = 6.53, p = 0.011) and females (χ2 (1) = 6.44, p = 0.011).
The projected number of animals needed at the end of a 24-month study can be drawn from Figure 4 for all four groups of rats when the number of rats needed at the end is known. The estimate is based on the survival data of 146 rats.
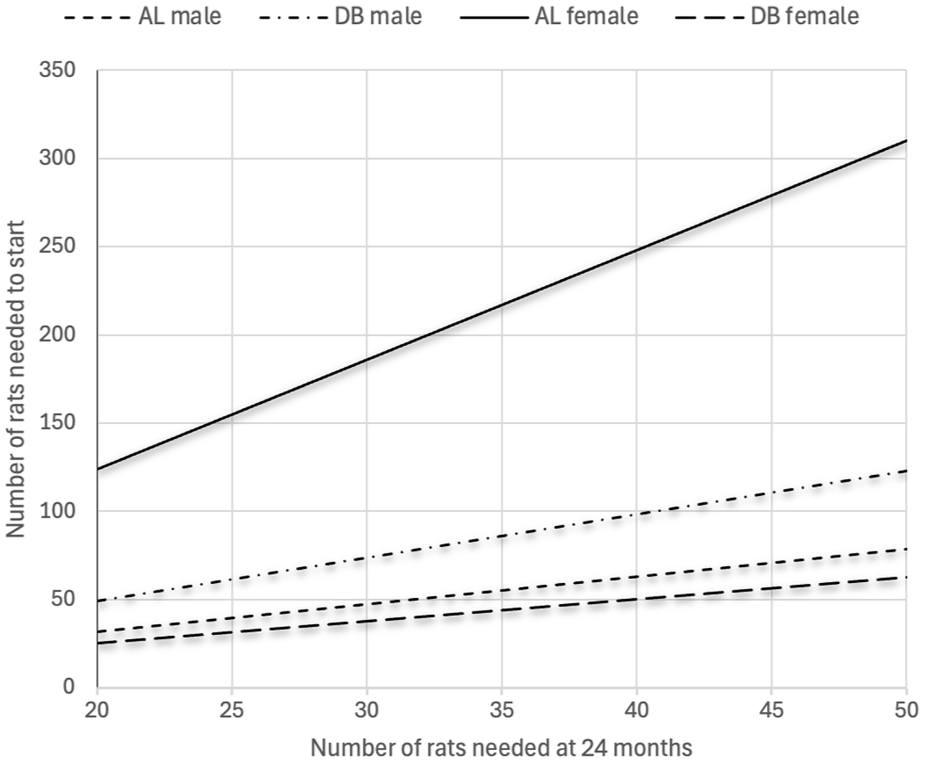
Projection of the number of rats needed at the start of a 24-month study in order to have a chosen number of rats alive at the end of the study. The projection is based on mortality data from four groups of Sprague Dawley rats in this study (N = 146): ad libitum (AL) males, diet board (DB) males, AL females and DB females.
Discussion
This two-year study evaluated the effects of diet board feeding on the serum and blood clinical chemistry and survival of Sprague Dawley rats, as well as the variability of clinical chemistry values and the potential reduction value of diet board feeding compared with traditional ad libitum feeding.
The earlier published parts of our study presented the results of diet board feeding on body weights, food intake and adiposity, as well as behaviour, stress and welfare.5,16 Diet board feeding reduced body weight by 11–19% in males and 6–11% in females over the course of the study. On average, food intake was 7% lower in diet board-fed than in ad libitum-fed males, but there was no difference between female groups. Diet board feeding reduced adiposity in both sexes, based on the epididymal fat weights of males (mean difference 32%) and body mass indexes of both sexes (a difference of 11–14% in males and 6–8% in females). The faecal corticosteroid metabolite levels were higher in diet board females compared with all other groups. In cage behaviour analysis and elevated plus-maze test, there were no evident behavioural signs of distress or impaired welfare due to diet board feeding, and it did not notably change the circadian distribution of behaviours. There were 30% more eating observations in diet board males and 85% more in diet board females than in the corresponding ad libitum groups. Diet board males were seen resting 4.2% less frequently than ad libitum males, but no difference was recorded between female groups.
The major clinical chemistry effects that we observed were associated with lipid and energy metabolism, consistent with the reduced energy intake and/or increased physical activity in diet board groups, and the results were largely comparable to findings in other dietary restriction studies examining mild to moderate dietary restriction and moderate physical exercise. Statistically significant findings included lower trig and chol levels in diet board than in ad libitum-fed males.
There were no significant differences in the Cl values between feeding groups in the whole dataset, but with extreme outliers of individual rats’ values removed, Cl values were significantly higher in diet board than in ad libitum males. There was no obvious explanation for the deviant values that would justify their removal. Considering the small effect-size and sensitivity of the result to outliers, the finding seems biologically irrelevant.
Trig levels in ad libitum-fed males were stable until 12 months and then increased considerably at 18 and 24 months. In diet board males, trig decreased from 0 to 3 months and remained stable thereafter. Aging rats have both an increase in the production of, and decline in the removal of, trig, leading to rising serum levels with age, especially during the second year. 22 We found this rising trend in all other groups except diet board males. Trig values in ad libitum males doubled towards the end compared with diet board males, whereas in females there was no difference. Our result is in line with earlier dietary restriction studies, considering that diet board feeding reduced food intake only in males (on average 7%), whereas food consumption did not differ between the female groups. 16 However, physical exercise (voluntary wheel running) can also prevent the rise in trig level. 22 As reported before, diet board-fed females in this study had lower body weights and body mass indexes and spent twice as much time eating compared with ad libitum-fed females, suggesting that diet board feeding increases physical exercise and energy consumption.5,16 Yet this exercise effect alone was not large enough to induce a reduction in trig.
In all study groups, chol levels first declined until six months and then rose towards the end. In diet board males, chol levels were significantly lower compared with ad libitum males, the difference showing from 12 to 24 months and peaking at the end. No difference was detected in females. Chol values have been reported to increase considerably in male and female ad libitum-fed Sprague Dawley rats by the age of 24 months. 23 Liepa et al. 24 reported a similar development as in our study between six and 24 months of age in male F344 rats with ad libitum feeding and 40% dietary restriction. In a 10-week diet board study with young male Wistar rats, Kasanen et al. 15 also reported significantly lower chol levels in diet board than in ad libitum-fed rats already at 10 weeks (17 weeks of age). By contrast, in some earlier studies with Sprague Dawley rats, no effects of dietary restriction on serum chol were found, with dietary restriction levels ranging from 25% to 55%.1,25
In other long-term experiments with mild to moderate dietary restriction in rats, the only consistent clinical chemistry finding has been a decrease in trig in both males and females.1,7,22,24–26 However, most other dietary restriction studies have been considerably shorter and the degree of food restriction higher than in this study, complicating the comparison of results.
The effects of dietary restriction on clinical chemistry variables are affected by a host of factors, including rat stock, sex and age, the degree and method of food restriction, food composition, the effects of meal timing on metabolic circadian rhythms, fasting before sampling and physical activity. All these might induce differences between the results of studies with seemingly similar experimental setups. It is noteworthy that the circadian rhythms of feeding, general activity and rest were similar in diet board and ad libitum-fed rats, and therefore there was likely no bias due to desynchronisation of circadian rhythms between groups in our study.5,17
As a limitation of our study, housing conditions and cage layout with diet boards could have influenced the rats’ activity and metabolism. Further factors such as non-fasted sampling, activity and handling-related stress might also have affected the results. Consequently, the interpretation of clinical chemistry findings should be treated with caution.
Dietary restriction has been claimed to decrease variation. Overfeeding-related metabolic problems and diseases are believed to increase variation in ad libitum-fed animals. A closer look at the studies on the topic shows that increased variation has been shown only with organ weights.9–12 Possible effects on variation are variable-specific, and this study assessed 21 clinical chemistry values with no significant variation differences attributable to diet board. Unlike our study, previous dietary restriction experiments used adjusted daily food quantities for rats. It could therefore be argued that the observed variation effects were due to precisely fixed quantities of food, rather than dietary restriction per se.
In this study, survival rates at 24 months for males were double for the diet board (71%) group compared with the ad libitum (36%) group, and over threefold for females in the diet board (42%) group compared with the ad libitum (11%) group (Table 1). These differences are surprisingly large considering that diet board feeding reduced food consumption by only about 7% in males, whilst in females no reduction at all was observed. 16 In other two-year dietary restriction experiments with a mild to moderate 10–28% restriction level, dietary restriction increased the two-year survival rate up to over double compared with the ad libitum-fed controls.4,9,27 In some previous studies, lifespan extension was relative to rising dietary restriction levels, whilst in others the greatest effect was already achieved with very mild dietary restriction.1,9,27,28 This discrepancy probably results from differences in rat strains or stocks, diet and animal care practices between institutions and studies. The deleterious effects of ad libitum feeding are not constant between institutions, and thereby the effect of a certain percentage level of dietary restriction compared with ad libitum feeding is not universally constant, either.2,4,29,30 In diet board feeding, the exercise effect of gnawing wood presumably adds significantly to the dietary restriction effect in survival. This conclusion accords with the studies of Holloszy, who found that voluntary wheel running increased the average lifespan in both male and female rats.31,32
The reduction potential with diet board feeding is illustrated in our projection (Figure 4). If one needs 25 male rats alive at 24 months, using a diet board one can start with 25 fewer rats than with ad libitum feeding. For female groups the difference is 125 rats, which stems from the exceptionally poor survival of ad libitum-fed females. In both cases, by using diet boards the reduction in the number of unnecessary animals in studies is remarkable.
This study is a two-year applicability evaluation of diet board feeding. Clinical chemistry results showed decreased values of trig and chol in males during the second year, attributable to diet board. We also evaluated reduction possibilities connected to survival and variable-specific changes in variance. We found no reduction potential in variances, but diet board feeding showed major efficacy in survival, reducing mortality in both sexes. It is especially noteworthy that diet board prevented the devastating mortality detected in ad libitum-fed female rats. Diet board certainly involves more animal husbandry work than standard laboratory practice, but it comes with a notable reduction potential. In conclusion, diet board has the potential to become a practical method for reduction in long-term rat studies which is also compatible with group-housing.
Footnotes
Ethical considerations
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study was supported by the ECLAM (European College of Laboratory Animal Medicine) and ESLAV (European Society of Laboratory Animal Veterinarians) Foundation, the Finnish Cultural Foundation, and Orion–Farmos Research Foundation.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The data this study is based on is freely available by contacting the corresponding author.