
Abstract
Seamless integration of visual, vestibular, and somatosensory (VVS) systems is key to human movement control. Emerging evidence suggests autonomic nervous system (ANS) function may influence this integration, yet no studies have examined the relationship between ANS activity, indexed by the pupil light reflex, and VVS function. 90 healthy adults (aged 20 – 88 years old) were recruited for this prospective cohort study. ANS function was quantified using pupil light reflex, where response delay, maximum pupil constriction and dilation velocities were recorded. VVS system function was quantified via visual-vestibular testing, including smooth pursuit and voluntary saccades, while somatosensory function was quantified using an Active Movement Extent Discrimination Assessment protocol. Multivariate multiple regression analysis was conducted in RStudio with significance set at p < .05. Follow-up univariate analysis of variance was conducted. VVS performance under differing autonomic profiles – Coactivation, Co-inhibition, Parasympathetic-Dominant and Sympathetic-Dominant – was assessed using one-way analysis of variance. Pupil response delay, maximum pupil constriction and age all significantly predicted combined VVS function (p = .01, p=<.01, p=<.01, respectively), with follow-up univariate analyses demonstrating small-to-moderate effects across individual VVS outcomes (ηp2=0.05 to 0.25). ANS profile of Co-inhibition was significantly correlated with worse VVS performance. This research highlights not just the need to consider ANS function in human movement control but also the ANS profile of the individual at rest. The identification of ANS Co-inhibition as a maladaptive profile in the context of VVS integration challenges existing frameworks that treat VVS and ANS as independent contributors to human movement control.
Keywords
Introduction
The perception-action theory of human movement control posits individuals directly perceive environmental information necessary for movement and act upon it without requiring extensive internal processing (Gibson, 1966). Emerging evidence indicates that the autonomic nervous system (ANS) may modulate the efficiency of this internal processing (Gordon et al., 2023), yet the relationship between sensorimotor systems and ANS activity, as indexed by the pupil light reflex (PLR), has not been examined. Understanding this interaction could provide valuable insight for the development of interventions targeting complex multisystem conditions that involve both autonomic and sensorimotor dysfunction, including ageing.
Optimal human movement arises from the continuous coupling between perception – sensory input from the visual, vestibular and somatosensory (VVS) systems – and action, which involves the motor systems’ response to that input (Gibson, 1966). The visual system provides spatial orientation and environmental context (Davids et al., 2005). The vestibular system detects changes in angular and linear acceleration, while also coordinating eye-movements in conjunction with the visual system (Cabolis et al., 2018). Tactile and proprioceptive inputs, components of somatosensation, inform the encoding of the body’s position in space relative to itself and the environment (Goodman & Bensmaia, 2020). These sensorimotor, or VVS, systems work in concert to continuously inform the central nervous system of the body’s position and motion in space.
Beyond the VVS systems, ANS function plays a role in human movement control by preparing and regulating the physiological states necessary for movement execution (Gordon et al., 2023). Despite established anatomical links between ANS and VVS systems (Bronstein, 2013; Cabolis et al., 2018; Vidal et al., 2015), visually depicted in Figure 1, few studies have examined collective functional interaction, with previous research limited to specific-subsystem relationships, such as ANS and vestibular function connections (Bronstein, 2013; Raphan, 2020). Recently, Lustig et al. (2025), explored broader VVS-ANS interactions, reporting that incongruences between visual cues and actual movement elicited changes in heart rate variability (HRV), suggesting autonomic adjustments to resolve visuomotor discrepancies. These findings indicate that ANS and VVS system functions are interdependent rather than discrete (Lustig et al., 2025). However, to date, no studies have examined ANS function using PLR-derived measures in conjunction with collective VVS performance, despite the substantial anatomical and functional overlap of these systems. Simplified schematic of overlap between autonomic nervous system and sensorimotor (visual, vestibular and somatosensation) pathways within the brain and spinal cord for visualisation purposes. Adapted from Bronstein (2013); Vidal et al. (2015) and Cabolis et al. (2018)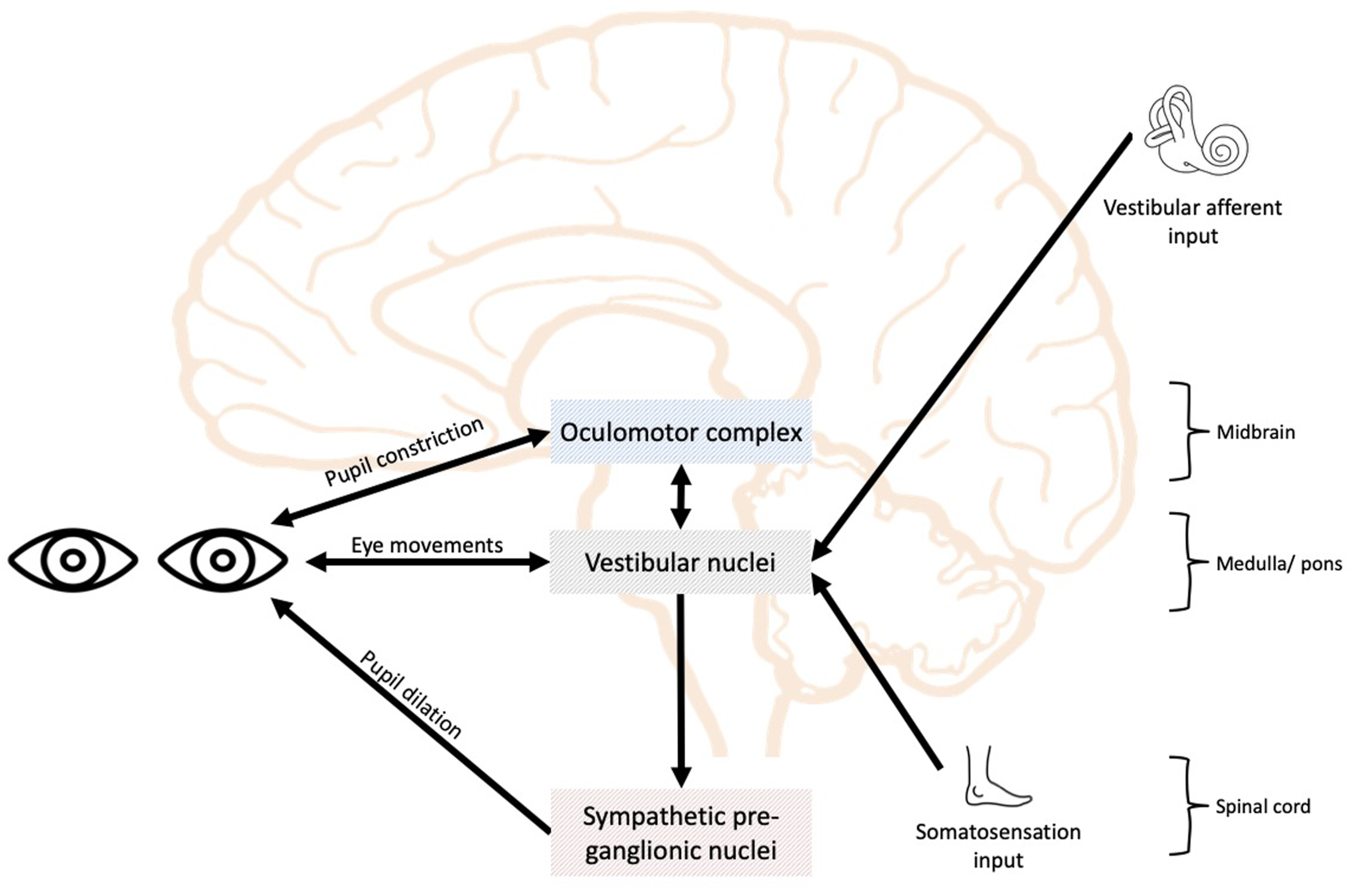
While HRV is a widely used marker of ANS function, it primarily reflects cardiac modulation, is influenced by numerous external factors, and does not independently capture sympathetic activity (Hayano & Yuda, 2019). The ANS comprises two distinct systems – the sympathetic nervous system (SNS) and the parasympathetic nervous system (PNS) – whose interaction produces four possible ANS profiles – SNS-Dominant, PNS-Dominant, Coactivation, and Co-inhibition (Lustig et al., 2025; Weissman & Mendes, 2021). Increased resting HRV, often interpreted as indicative of a PNS-Dominant profile, has been associated with better aerobic and coordination performance in athletes (Shaffer & Ginsberg, 2017; Tekin et al., 2025). However, while some studies attempt to infer ANS profiles using HRV measures, SNS and PNS contributions cannot be fully separated (Billman, 2013). Consequently, HRV may not be the most comprehensive measure of ANS function, offering limited insight into these distinct autonomic profiles, at a single time point (Billman, 2013). In contrast, the PLR may provide a more direct assessment of ANS function through measures of pupil response delay (PRD), maximum pupillary constriction velocity (MPCV), and pupil dilation velocity (PDV) (Hall & Chilcott, 2018). PRD reflects the latency between light stimulus and pupil constriction, serving as a proxy for central neural processing speed across midbrain and brainstem circuits (Hall & Chilcott, 2018). In contrast, MPCV reflects the rate of pupil constriction following response onset, representing the strength of the PNS efferent response (Hall & Chilcott, 2018). Finally, PDV reflects the SNS-driven recovery phase of the PLR, arising from the concurrent inhibition of the PNS constriction pathway and activation of the SNS innervated iris dilator muscle, with these pathways of control depicted in Figure 2 (Hall & Chilcott, 2018; Mathôt, 2018). This suggests that PLR could offer a more comprehensive marker of ANS function, particularly in contexts involving VVS integration, though its superiority over HRV has not yet been established. Simplified schematic of the pupil light reflex. Where the blue pathway represents the parasympathetic activation, and the red pathway represents the sympathetic activation. Adapted from Mathôt (2018)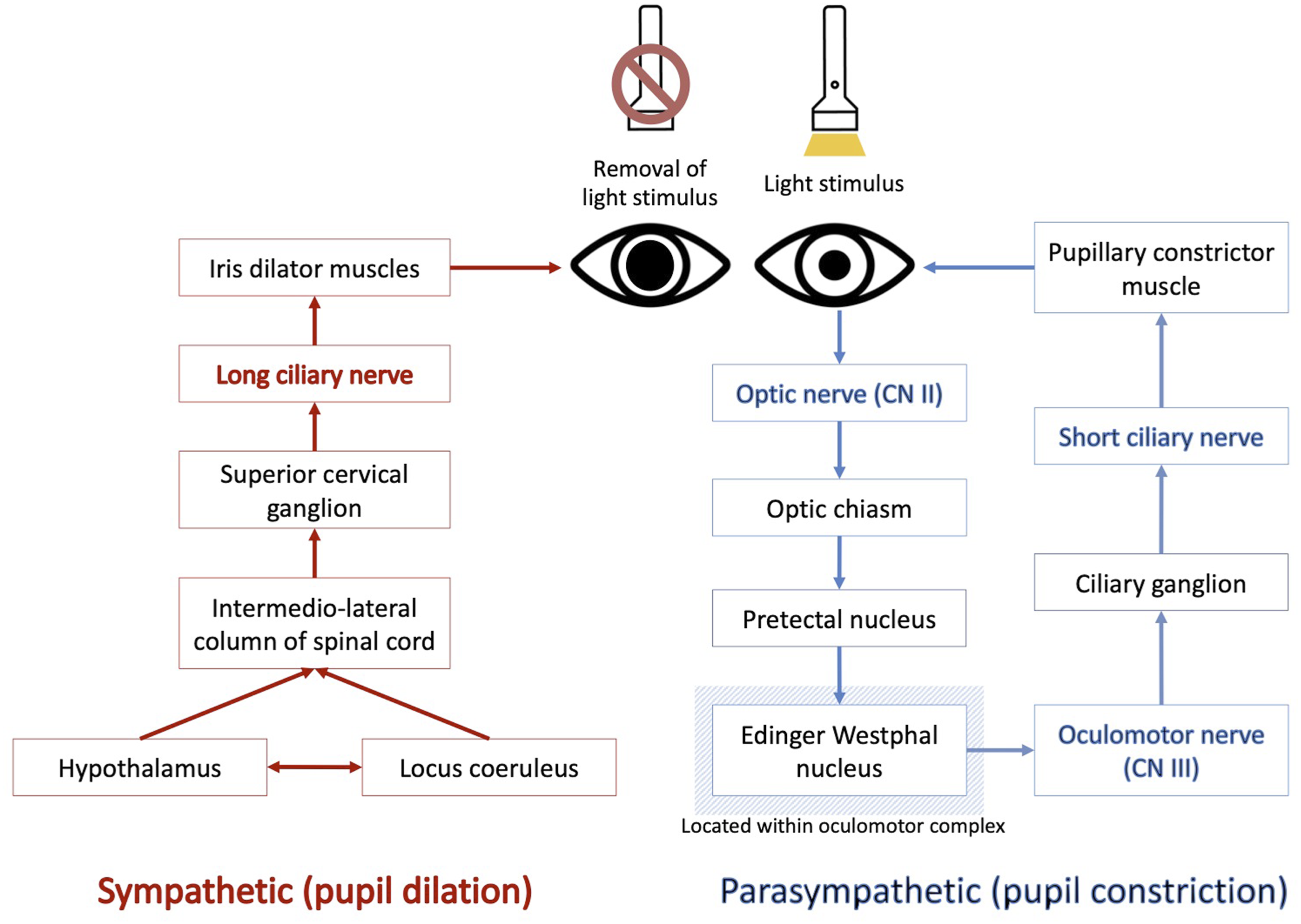
Establishing the relationship between ANS function as measured by PLR, and VVS performance, may be important to advance our understanding of human movement control. This is particularly relevant in conditions such as ageing that are already known to affect both VVS and ANS function (Peltsch et al., 2011; Yang et al., 2019). Similarly, ANS dysfunction is a prevalent feature in many multi-system diseases, such as Long COVID (Ferreira et al., 2023), where symptoms consistent with VVS dysfunction are also often reported. It is therefore plausible that abnormalities in movement control seen in these contexts may stem from disruptions in the integration of these systems. Quantifying the simultaneous contribution between ANS and VVS function is therefore important to develop comprehensive intervention strategies that address both systems concurrently. A more integrated approach could lead to improved outcomes, both in age-related declines and populations with complex multisystem conditions.
This study is the first to investigate the relationship between ANS function, as measured by the PLR, and VVS performance assessed through central pathways. The primary aim of this study is to examine how PLR-derived ANS metrics influence VVS ability while controlling for age-related differences. The secondary aim is to investigate whether autonomic profiles i.e., Coactivation of PNS and SNS activity, Co-inhibition of PNS and SNS activity, PNS-Dominant (activated PNS and inhibited SNS), or SNS-Dominant (activated SNS and inhibited PNS), are associated with differences in VVS performance. We hypothesised that elevated PNS function and faster PRD would be associated with increased VVS performance, while heightened SNS function would not be independently associated with increased VVS performance. For the secondary aim, we hypothesised that individuals with Co-inhibition of PNS and SNS activity will demonstrate reduced VVS performance. This research is novel, offering a more nuanced understanding of how ANS function contributes to movement control, potentially opening new avenues for integrated clinical interventions.
Method
Participants and Ethical Considerations
Participant Demographics and Descriptive Data
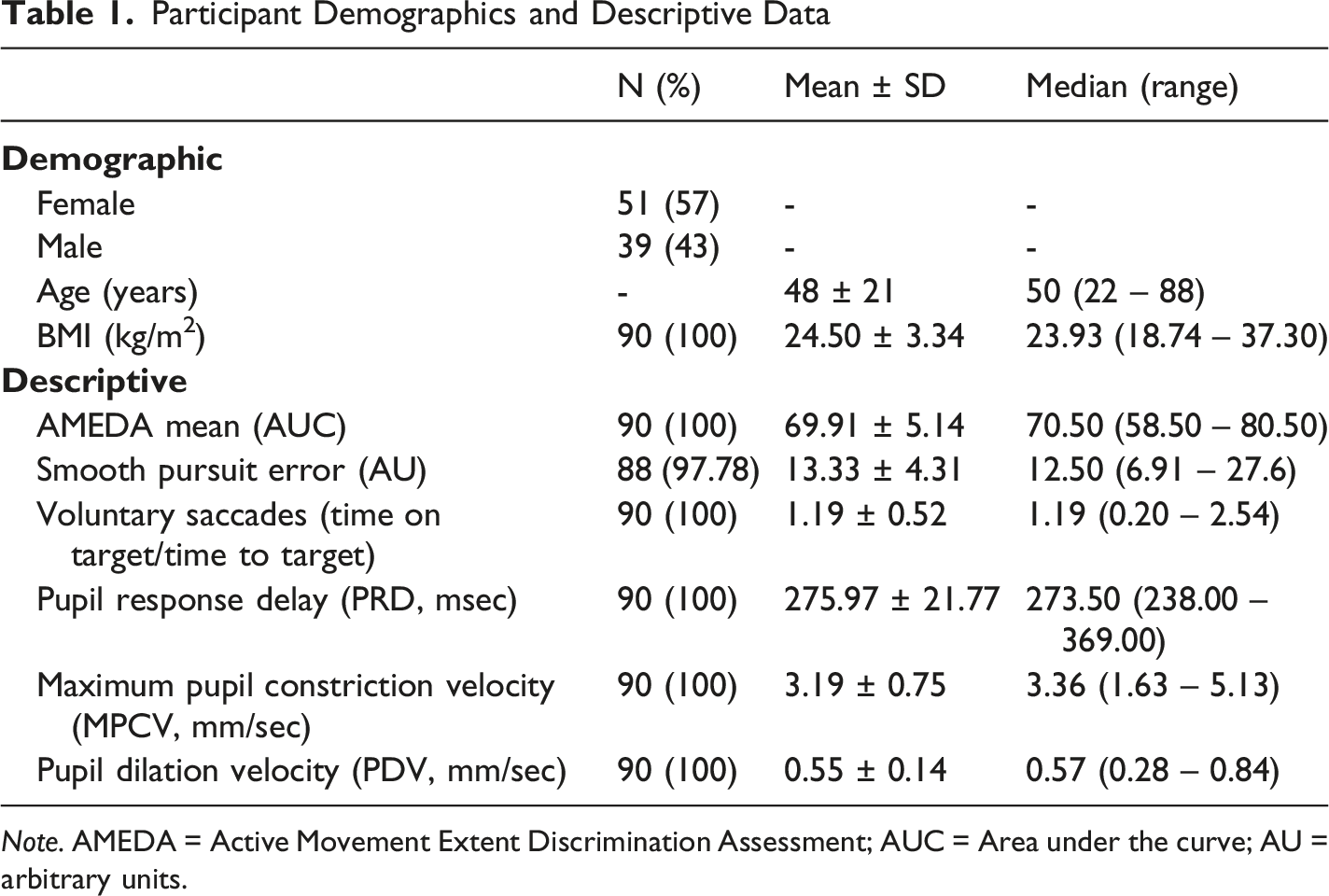
Note. AMEDA = Active Movement Extent Discrimination Assessment; AUC = Area under the curve; AU = arbitrary units.
An a priori power analysis was conducted using G*Power version 3.1 (Faul et al., 2007) for a univariate regression model, with four continuous predictors (PRD, MPCV, and PDV and age). A medium effect size (f2 = 0.15 (Cohen, 1988)), power = 0.80 and alpha = 0.05 were specified. The required sample size was estimated at n = 85. To account for potential attrition or unusable data, we therefore aimed to recruit 90 participants.
Procedure
Participants attended a single laboratory-based testing session, where they provided written informed consent, completed the Exercise and Sports Science Australia Pre-Exercise Screening tool, and had their height and weight recorded. Participants then completed the VVS protocol outlined in detail in McGrath et al., (2025), which consisted of two 5-min assessments: an eye-tracking assessment and the Active Movement Extent Discrimination Assessment (AMEDA Figure 3). Noise and environmental distractions were removed by conducting all testing sessions in a quiet, closed door, temperature-controlled laboratory, with consistent lighting. Sensorimotor and autonomic testing equipment where A. is the headset assessment and B. is the Active Movement Extent Discrimination Assessment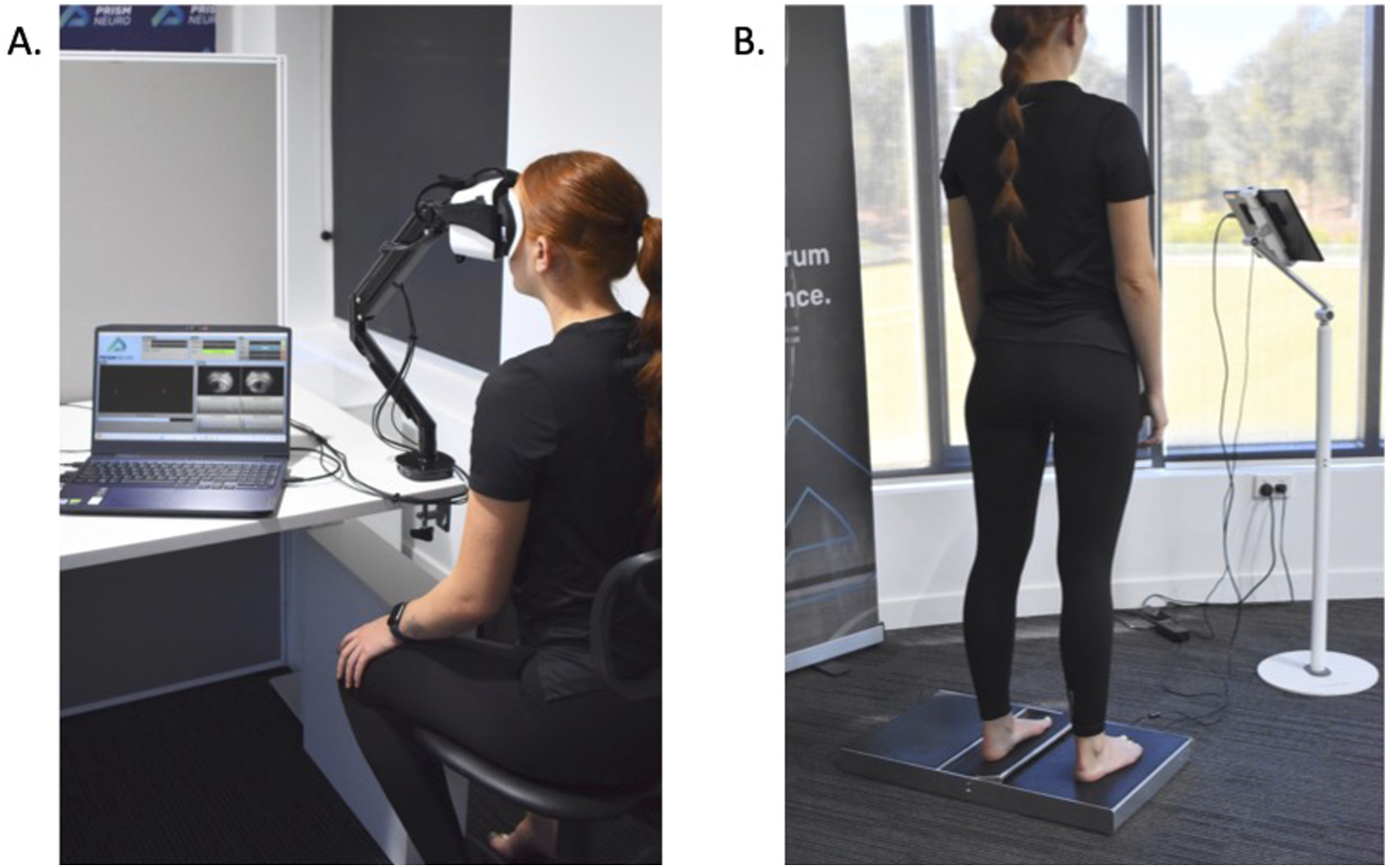
The first half of the VVS protocol utilises a virtual reality headset (Figure 3(a), Prism Neuro®, Canberra, ACT), and measures both pupillometry and eye-tracking metrics with three tests. Prior to testing, participants underwent a calibration within the headset, with testing commencing immediately thereafter. The first test assessed circular smooth pursuit, where participants were asked to track a small red dot moving in a circular path, with their eyes. Smooth pursuit performance was calculated as the root means square error between the participants gaze position and the stimulus, with higher scores (reported in arbitrary units, AU) indicating greater error. The second test measured ANS function using the PLR. Participants gaze fixated on a red ‘x’ while lights flashed, and pupil constriction and dilation velocities were recorded. PRD was calculated as the time taken for the pupil to react to the light, PNS function was measured via MPCV and SNS function was collected via average PDV (Hall & Chilcott, 2018). The final test measured voluntary saccades, where participants followed a rapidly moving red ‘x’ across the screen. The voluntary saccade metric was calculated as the proportion of time spent accurately on the target relative to the time taken to reach it. All data collected via the headset underwent proprietary automatic blink filtering and signal smoothing using a low-pass filter (McGrath et al., 2025).
The second half of the VVS protocol assessed somatosensory system performance using AMEDA (Figure 3(b), Prism Neuro®, Canberra, ACT), which evaluates a participant’s ability to discriminate between different ankle inversion angles (Waddington & Adams, 1999a, 1999b; Witchalls et al., 2014). Participants stood with one foot on a stable platform and the other on a tilting platform capable of five controlled inversion angles (10 – 14° in 1° increments). Testing was conducted on the left side first, followed by their right. Participants were familiarised with the five movement extents in shallow-to-deep order across three trials before formal testing. During testing, each level was randomly presented 10 times, totalling 50 trials per side. After each tilt, participants returned the platform to the start position and immediately reported the perceived level. Discrimination accuracy was calculated using the Area Under the Curve (AUC) from receiver operating characteristic analyses for each adjacent pair (1 vs. 2, 2 vs. 3 etc.). The final score was the average AUC across all four comparisons, converted to a percentage. A score of 100% represents perfect joint position sense discrimination between all positions, while a score of 50% indicates performance equivalent to chance (McGrath et al., 2025).
Statistical Analysis
All statistical analyses were conducted using RStudio (Version 2024.09.0 + 375), and statistical significance was set at p < .05. Descriptive statistics, including means and standard deviations (SD), were calculated for all variables. To address the primary research question – how ANS function, indexed by PRD, MPCV (PNS activity), and PDV (SNS activity), influences VVS performance (AMEDA mean, voluntary saccades, and smooth pursuit) – a multivariate multiple regression (MMR) was conducted (Alexopoulos, 2010). This approach, modelling multiple intercorrelated dependent variables simultaneously, has been previously applied in physiological research (Wang et al., 2012). Age was included as a covariate to account for potential age-related effects on both ANS and VVS function.
Prior to analysis, assumptions were evaluated. Normality was assessed using the Henze-Zirkler test and Anderson-Darling test (MVM package), which indicated a violation of normality. Smooth pursuit was non-normally distributed, and was subsequently log-transformed to reduce skewness, after which normality assumptions were satisfied (Appendix 1). Multicollinearity among predictors was assessed using variance inflation factors, where values below the threshold of 10 indicate acceptable collinearity. Correlation matrices and scatterplot panels were generated using the psych package to visualise correlations among variables. Pillai’s Trace was selected as the MMR test statistic (Ateş et al., 2019). The MMR analysis was run using the base stats package. To further explore individual dependent variables, separate univariate analyses of variance (ANOVAs) were conducted, with partial eta squared (ηp2) also calculated to evaluate effect size.
To address the secondary research question, whether categorical ANS imbalance affects VVS performance, participants were classified – after completion of all VVS testing – into one of four ANS profiles based on standardised z-scores of MPCV, and PDV. The classification included: (1) Coactivation, defined by both z-scores > −1 and an absolute difference ≤0.5; (2) Co-inhibition, defined by both z-scores < −1; (3) PNS-Dominant, defined by a PNS z-score exceeding SNS by > 0.5 or when PNS z-score was positive and SNS z-score negative; and (4) SNS-Dominant, defined by a SNS z-score exceeding PNS z-score by > 0.5. This approach, while ensuring all participants were assigned to a category without exclusions, was based on arbitrary z-score thresholds rather than established physiological cut-off points. These thresholds are consistent with those used in prior work (Schober et al., 2021).
One-way ANOVAs were performed for each VVS outcome to compare across ANS profiles. Assumptions of normality and homogeneity of variances were tested using the Shapiro-Wilk and Levene’s tests, respectively. For significant main effects, post hoc pairwise comparisons were conducted using Tukey’s Honestly Significant Difference (HSD) test, with adjusted p-values reported to control for Type I error. Box plots were generated to visualise differences in VVS outcomes between ANS profiles. Boxplot showing A. AMEDA mean scores across autonomic nervous system profiles, where higher scores represent better somatosensory performance; B. smooth pursuit performance by autonomic profile groups, where higher values indicate worse visual-vestibular tracking ability and C. voluntary saccades performance by autonomic profile groups where, higher values indicate improved visual-vestibular targeting ability.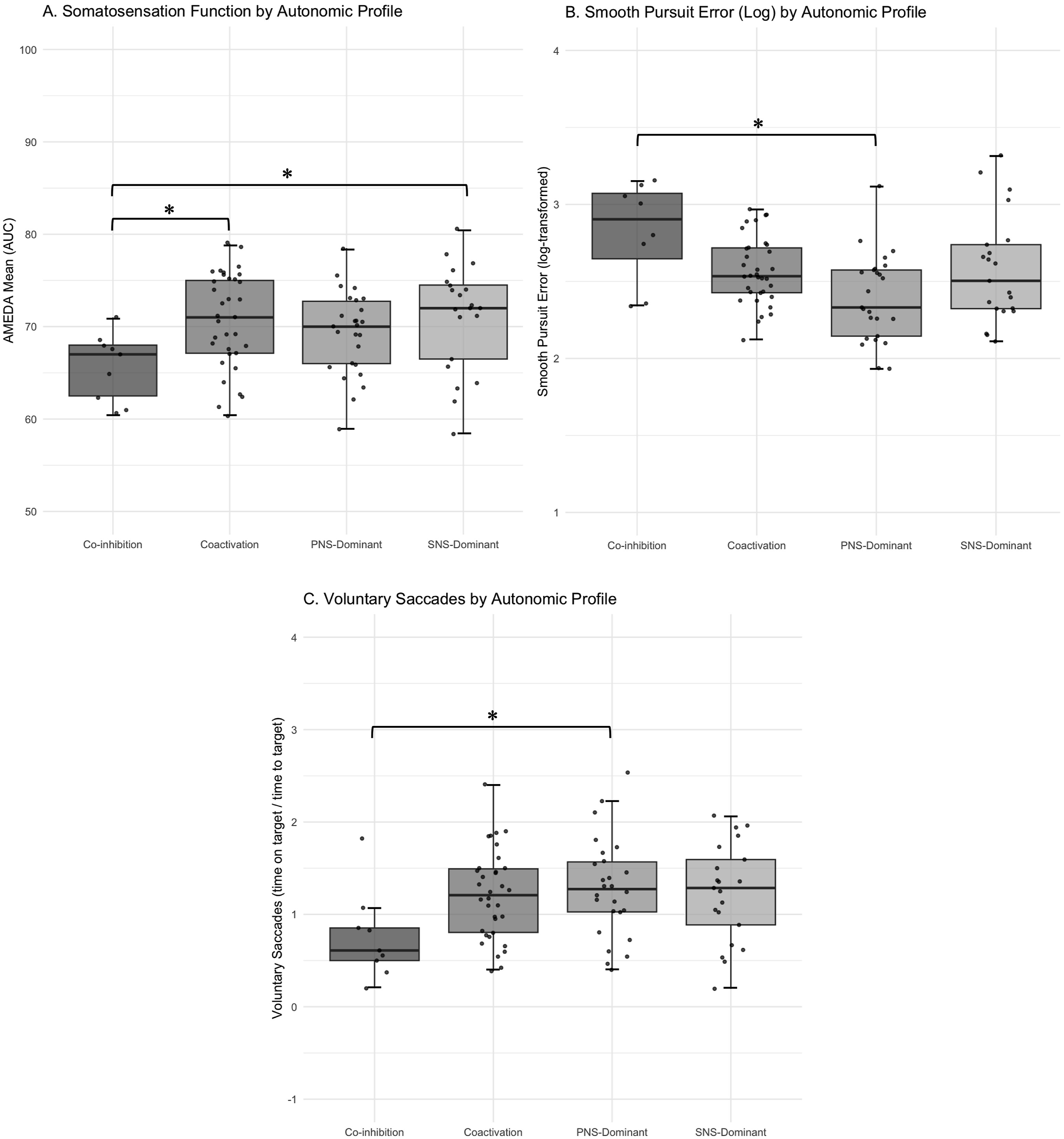
Results
Participants’ demographic descriptors and VVS scores are provided in Table 1.
Prior to conducting the MMR, multicollinearity among predictors was assessed using variance inflation factor. All values were below the accepted threshold of 10, indicating acceptable collinearity: PRD = 1.39, MPCV = 2.10, PDV activity = 1.84 and age = 1.38. Normality was evaluated using the Anderson-Darling and Henze-Zirkler tests. Following log-transformation of the smooth pursuit variable, all variables met normality assumptions (Appendix 1). Relationships among VVS and autonomic measures were visually assessed using a Pearson correlation matrix (Appendix 2). Most variable pairs demonstrated weak to moderate correlations (r = 0.10 – 0.69, (Schober et al., 2021)), consistent with recommendations to avoid excessive multicollinearity, which can obscure independent effects in subsequent analyses (Tabachnick et al., 2013). PRD and AMEDA mean showed negligible correlation (r = 0.02), indicating little shared variance.
Multivariate Multiple Regression Analysis Examining the Effects of Autonomic Nervous System Function (Pupil Response Delay, Maximum Pupillary Constriction Velocity, and Pupil Dilation Velocity) and Age on Combined VVS Performance
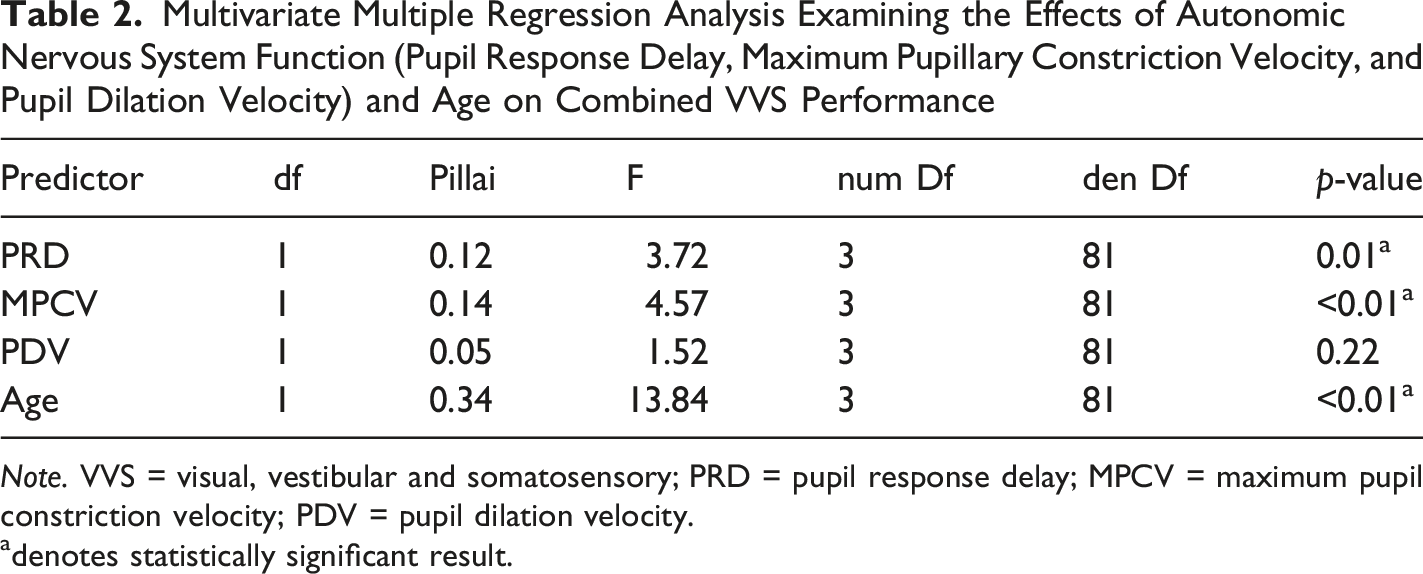
Note. VVS = visual, vestibular and somatosensory; PRD = pupil response delay; MPCV = maximum pupil constriction velocity; PDV = pupil dilation velocity.
adenotes statistically significant result.
Results From Follow-Up Univariate Analysis of Variance
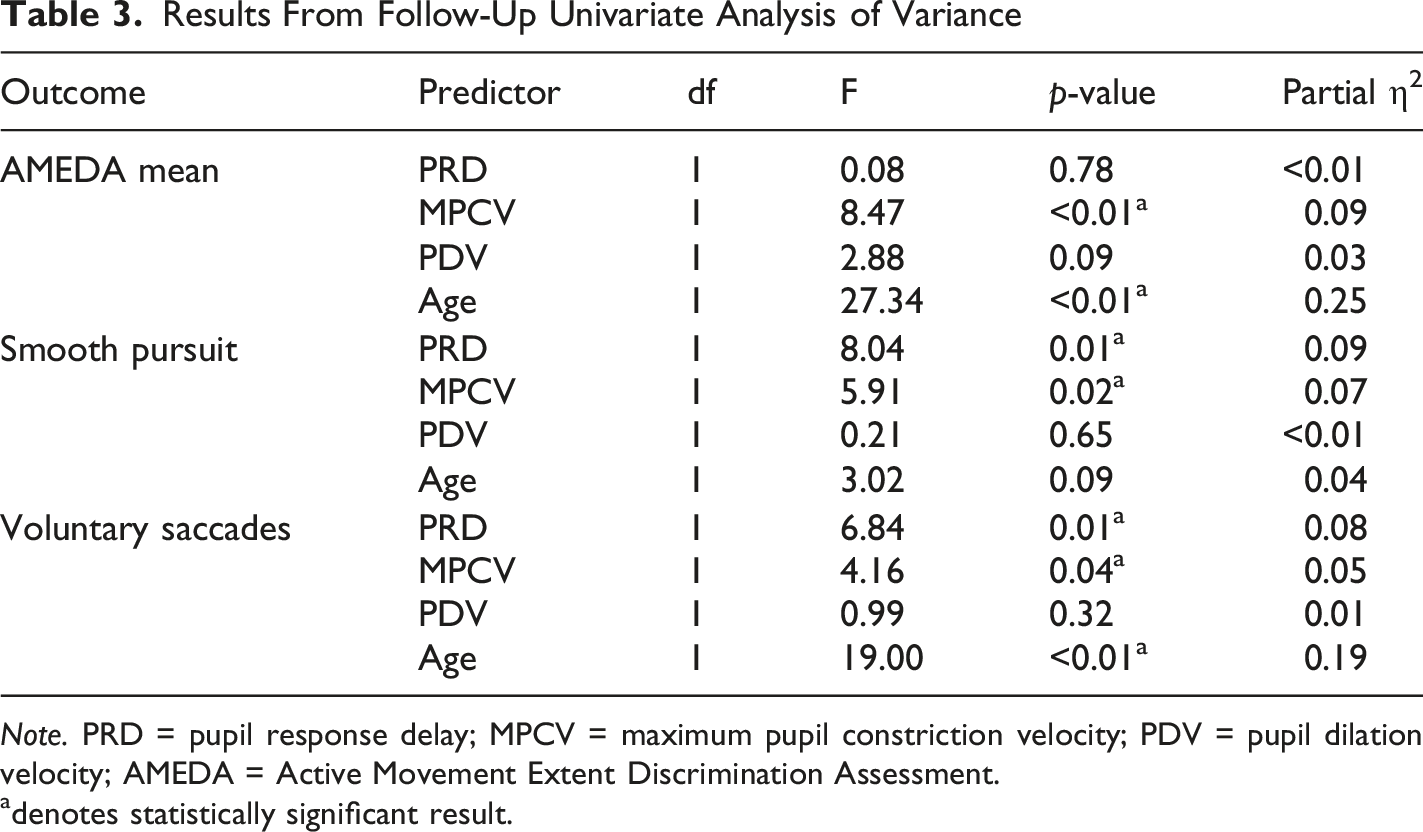
Note. PRD = pupil response delay; MPCV = maximum pupil constriction velocity; PDV = pupil dilation velocity; AMEDA = Active Movement Extent Discrimination Assessment.
adenotes statistically significant result.
One-Way ANOVA Results for VVS Performance Across Autonomic Nervous System Profiles
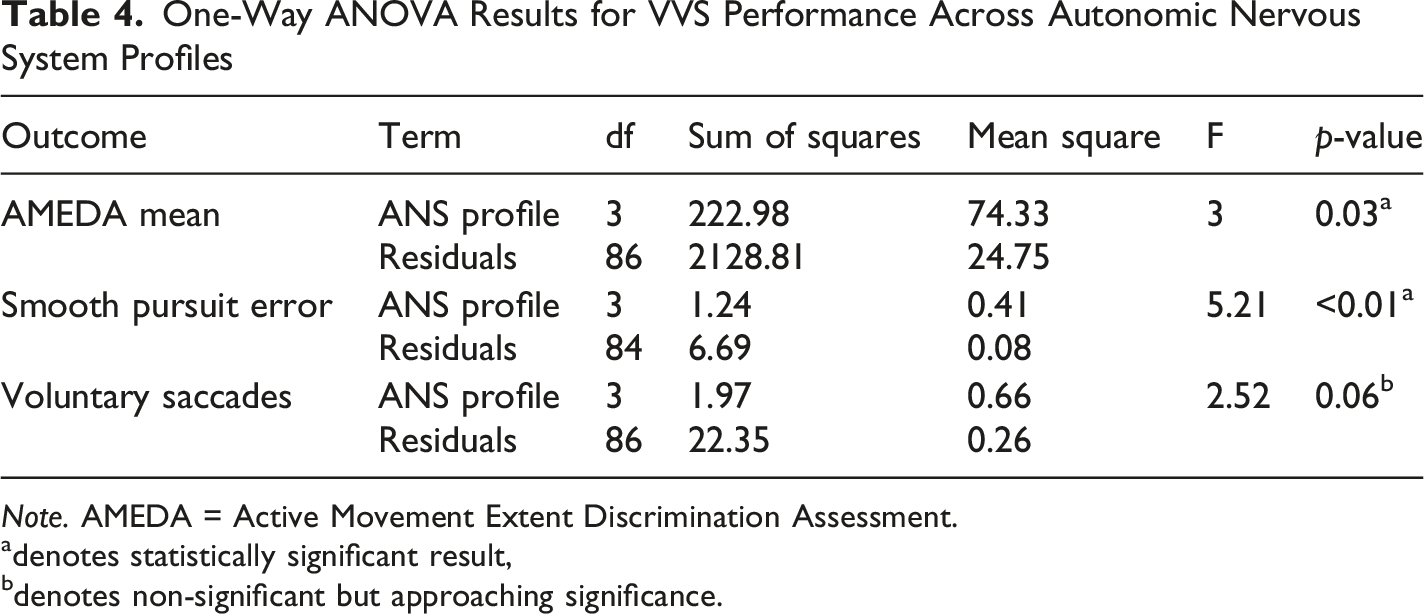
Note. AMEDA = Active Movement Extent Discrimination Assessment.
adenotes statistically significant result,
bdenotes non-significant but approaching significance.
Tukey HSD Post hoc Comparisons of Sensorimotor Outcomes Between Autonomic Nervous System Profiles
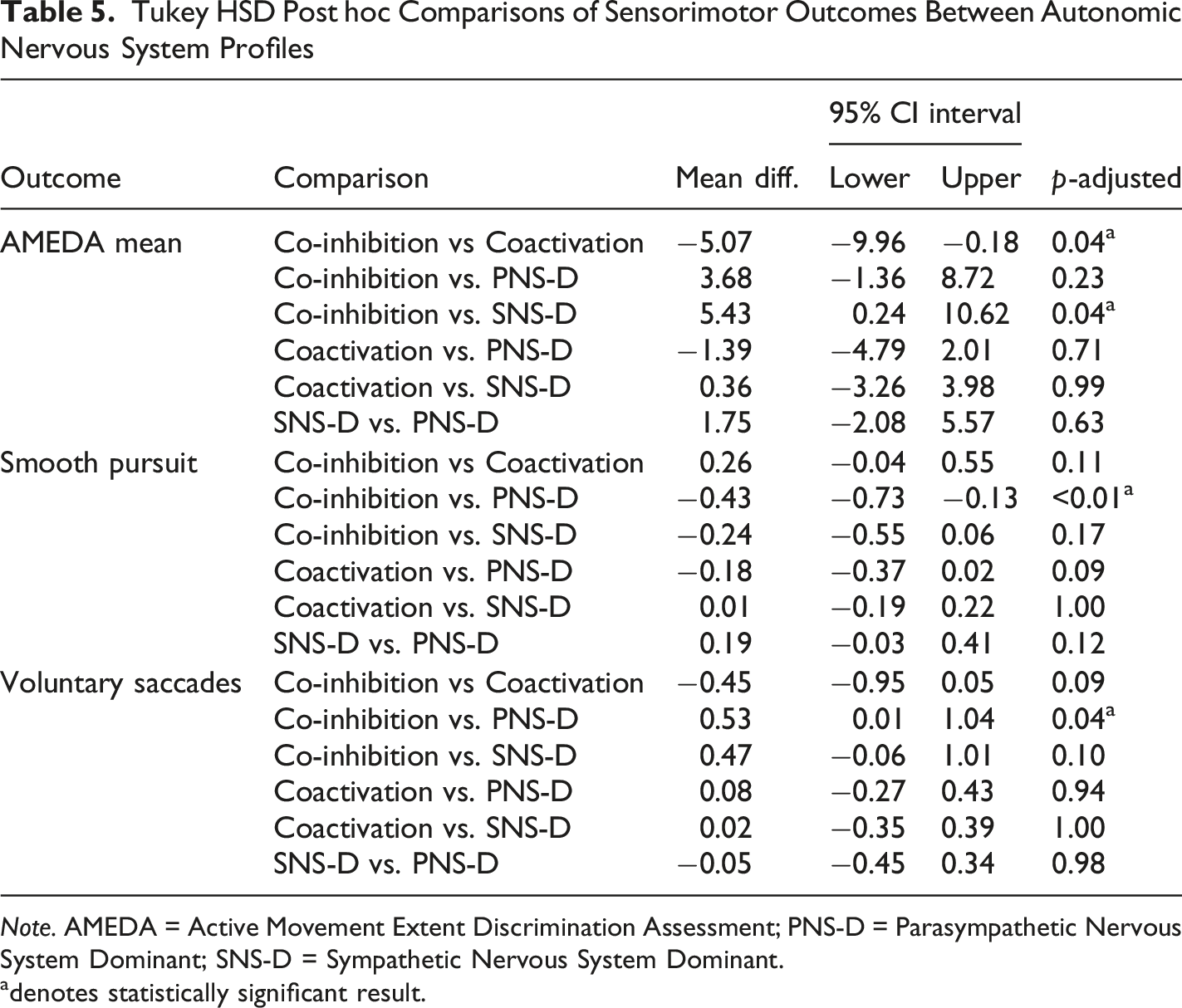
Note. AMEDA = Active Movement Extent Discrimination Assessment; PNS-D = Parasympathetic Nervous System Dominant; SNS-D = Sympathetic Nervous System Dominant.
adenotes statistically significant result.
Discussion
This study was the first to investigate the relationship between ANS function, via PLR measures, and VVS performance. In support of our hypothesis this study found that specific ANS metrics – PRD and MPCV– were significantly associated with VVS performance in healthy individuals. Further analysis revealed that VVS outcomes were most affected when both PNS and SNS systems were underperforming, or Co-inhibited, demonstrating that coordinated ANS function is important for optimal VVS integration.
The MMR revealed significant effects of PRD and MPCV on combined VVS outcomes. Follow-up univariate ANOVAs further clarified these effects; PNS activity, indexed by MPCV, was a significant predictor across all three VVS measures and PRD was significantly associated with smooth pursuit error and voluntary saccades. These findings suggest that stable PNS function provides a physiological context that supports sensory integration, while central neural processing speed (PRD) enables the efficient translation of sensory input. This interpretation is consistent with emerging models of motor cortex organisation, which show that effector-specific motor regions are integrated within a boarder somato-cognitive action network (Gordon et al., 2023). This framework supports the idea that effective movement control depends not only on isolated motor pathways but integrated systems linking somatosensory processing and autonomic regulation. These findings also align with previous research that links reduced HRV, an indicator of PNS function, to movement control disorders (Maurer et al., 2016). Studies have also shown the reverse, that increased resting HRV is associated with improvement in coordination tasks (Tekin et al., 2025). Our results build on this relationship, by demonstrating that PNS activity and PRD significantly predict VVS performance, extending beyond HRV studies by leveraging PLR to better isolate ANS contributions.
One-way ANOVAs based on categorical ANS profiles further supported the multivariate findings. Significant group differences were observed for AMEDA mean and smooth pursuit, with voluntary saccades approaching significance. Post hoc comparisons revealed that ANS profiles with heightened PNS activity (Coactivation and PNS-Dominant) were associated with improved VVS measures. Specifically, AMEDA performance was significantly lower in those with ANS profile of Co-inhibition compared to those with Coactivation or SNS-Dominant profiles. Together, these findings reinforce the role of ANS balance in facilitating precise and adaptive movement control by demonstrating that reduced function across both ANS branches may limit the capacity of the broader somato-cognitive action network (Gordon et al., 2023). Future research could extend this work by examining how interactions between ANS and VVS function relate to clinical measures of movement control, such as the Timed-up-and-Go.
This study did not find consistent associations between PLR derived measures of SNS function and VVS performance. SNS arousal has been linked to heightened alertness and preparatory motor states, however, PDV within the PLR reflects the functional efficiency of the SNS pathway under controlled conditions, rather than moment-to-moment arousal (Collet et al., 2013; Hall & Chilcott, 2018). Accordingly, our findings suggest that the isolated contribution of SNS pathway function to VVS measures may be less critical, particularly when PNS function is also considered. Post hoc analysis demonstrated that AMEDA scores were significantly higher in the SNS-dominant group, suggesting that this specific VVS measure may be more sensitive to SNS pathway contribution than smooth pursuit or voluntary saccades. This result aligns with known anatomical pathways of SNS innervation of skeletal muscle spindles, the origin of body position afferents (Radovanovic et al., 2015). This may be one possible explanation as to why heightened SNS function pathway efficiency appears to correlate with improved somatosensory performance but appeared to have limited effect on visual-vestibular measures. However, the role of the SNS in precise movement control remains incompletely understood, potentially due to limitations in commonly used ANS metrics, like HRV, which cannot fully isolate SNS from PNS contributions.
Age also emerged as a significant predictor across VVS ability, particularly for AMEDA mean and voluntary saccades. This is consistent with previous research showing age-related declines in these systems (Peltsch et al., 2011; Yang et al., 2019). For example, Peltsch et al. (2011) examined eye movements in 81 healthy older adults and found that the oldest participants were the slowest to initiate voluntary saccades and exhibited more directional errors. Furthermore, Yang et al. (2019), the first to apply the AMEDA method across the lifespan, demonstrated that AMEDA scores reached their peak in young adults and were at their worst in adults aged 70 years and above. This study was the first to examine these VVS systems in combination across the lifespan and collectively suggest that age significantly and independently affects VVS performance.
Study limitations must be considered when interpreting findings. The cross-sectional design limits causal inference between ANS and VVS function but allows direct comparison under the same physiological conditions – valuable given the ANS’s ability to fluctuate rapidly (Bronstein, 2013). Longitudinal studies could clarify directionality, and interventions (e.g., vagal stimulation) may help establish causality. The fixed assessment order may represent a minor limitation, as fatigue or learning effects cannot be fully excluded; however, this sequence was intentionally used to ensure ANS-function was assessed in a rested state. Future studies may benefit from counterbalancing test order, given adequate rest periods. Another limitation relates to statistical power. While an a priori power analysis was conducted for a univariate regression, the study ultimately employed a multivariate approach, and no power analysis was conducted for the secondary research question. This pragmatic decision reflects both the limited availability of formal tools for multivariate designs and the fact that ANS profiles were not pre-determined recruitment targets. Although these factors may increase the risk of a Type II error, the presence of significant findings suggests that statistical power was adequate. This approach also preserves ecological validity by reflecting real-world variability in ANS profiles. ANS profile classifications used a consistent z-score threshold of −1 to define below average activity, aligning with conventional norms (Schober et al., 2021). While somewhat arbitrary, this threshold was applied consistently across all participants, allowing reproducibility and interpretability. Future studies could refine these thresholds using physiological benchmarks.
These findings have clinical implications, particularly in the context of assessing and managing patients with VVS impairments. Individuals exhibiting Co-inhibition of PNS and SNS function demonstrated reduced VVS performance, a profile that may be clinically relevant given established associations between impaired somatosensory function and increased falls risk (Antcliff et al., 2023). This suggests that sensorimotor rehabilitation may benefit from concurrent consideration of ANS function rather than targeting VVS components in isolation. In contrast to HRV-based studies that primarily index PNS activity (Tekin et al., 2025), PLR-derived measures enable concurrent assessment of PNS and SNS function, allowing identification of Co-inhibitory ANS-profile that may be overlooked when relying on HRV alone.
Conclusion
This study demonstrates that ANS function, particularly PNS activity and response delay indexed via PLR, is a significant predictor of VVS performance in healthy individuals. ANS profiling revealed that Co-inhibition of PNS and SNS emerged as a maladaptive profile in this context, associated with poorer outcomes across multiple VVS measures. These findings extend previous research linking ANS profiles to functional performance by highlighting the role of ANS function in sensorimotor integration. Future studies could investigate whether integrated rehabilitation protocols that concurrently address ANS and VVS function enhance movement control performance.
Supplemental Material
Supplemental Material - Do Measures of Autonomic Nervous System Function Correlate With Sensorimotor Performance?
Supplemental material for Do Measures of Autonomic Nervous System Function Correlate With Sensorimotor Performance? by Michaela McGrath, Bernie Bissett, Angie Fearon, Jeremy Witchalls, Gordon Waddington in Perceptual and Motor Skills
Footnotes
Acknowledgments
We would like to thank all participants for their time and contribution to this study. Your involvement has been invaluable to our research.
Ethical Consideration
This study was approved by the University of Canberra Human Research Ethics Committee (HREC – 13197). All procedures performed in this study involving human participants were in accordance with the ethical standards of the institutional and with the 1964 Helsinki declaration and its later amendments.
Consent to Participate
Informed consent to participate in this study was obtained in writing from all individual participants included in the study.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication ofthis article: This research was supported by an Australian Government Research Training Program (RTP) Scholarship ![]() . This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Declaration of Conflicting Interests
The authors declare the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Professor Gordon Waddington is a founding shareholder in Prism Neuro Pty Ltd an Australian perceptual neuroscience equipment company. The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The lead author, Michaela McGrath, shares the surname of founders of Prism Neuro Pty Ltd however receives no financial support from Prism Neuro Pty Ltd nor holds shares in this entity.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Supplemental Material
Supplemental material for this article is available online.
Author Biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.