
Abstract
Injection of cannabis resin as a saline-Tween 80 solution to pregnant rats resulted in pups having lower body weight, higher emotionality, poorer learning capacity, and paradoxically higher brain weight and DNA content. The cannabis appeared directly to have caused the inferior growth rate of the pups which in turn affected emotionality and learning.
While a number of studies have been conducted to assess the direct effects of marihuana and related drugs on animals and humans, few have investigated the effects of administration of the drug during pregnancy on the offspring. Carakushansky, Neu, and Gardner (1969) and Hecht, Beals, Lees, Jolly, and Roberts (1968) have reported cases of syndactyly and absence of parts or all of the hand in a baby born to a mother who had been exposed to marihuana during pregnancy. Geber and Schramm (1969) studied the effect of marihuana resin injected subcutaneously in pregnant hamsters and rabbits and found embryocidal effects on rabbits. Persaud and Ellington (1968) reported teratogenicity of cannabis resin when administered daily at a dose level of 4.2 mg/kg body weight to rats on Days 1 to 6 of gestation. Teratogenicity took the forms of syndactyly, encephalocoete, phocomelia, hermia, and amelia. In another study, these same authors (Persaud & Ellington, 1967) administered a daily dose of 16 mg/kg body weight intraperitoneally to mice on Days 1 to 6 of gestation and observed complete fetal resorption. Intraperitoneal injection of cannabis resin evidently caused intrauterine death and reduced fetal body weight of survivors in mice. There is evidence that exposure of marihuana to mothers during critical stages of pregnancy or periods of most rapid differentiation in fetal organ systems can have severe physiological consequences in fetus and infant.
There have been no reports on the behavioral development of the surviving pups who were exposed to marihuana or an active ingredient during intrauterine and postnatal life. The reported presence of changes in brain characteristics by Persaud and Ellington leads one to an expectation that behavioral effects during later development ought to be found. This study was designed to investigate behavioral effects on offspring of cannabis resin injections to rat mothers during pregnancy. The behavioral measures were chosen to re-flea both learning capabilities and emotionality.
Method
Female Wistar rats (averaging 209 gm.) were purchased from Woodlyn Farms, Guelph, Ontario and mated in our laboratory. Upon confirmation of pregnancy by vaginal smears or presence of plugs, every other one of 16 mothers was put into an experimental or a control group. The pregnant animals were maintained in individual cages with Purina Lab Chow and water available ad libitum. They were housed in temperature- and humidity-controlled animal rooms in which the lights were turned on 12 hr. and off 12 hr. each day. Weight and food consumption were recorded daily. The mothers were placed in nesting boxes on Day 20 of gestation.
Cannabis resin (obtained from Tunney's Pasture, Ottawa, Canada) was used. The resin was composed of 54.9% Δ9-tetrahydrocannabinol, 19% cannabidiol, and 14% cannabinol. A suspension of cannabis resin in 0.9% saline containing 1 % Tween-80 was administered to eight pregnant rats. Each animal received a dose of 4.2 mg/kg body weight of cannabis resin in 0.2 ml of innoculum daily from Days 2 to 6 of gestation. A control group (N = 8) was given intraperitoneal injections of 0.9% saline containing 1% Tween-80 in 0.2 ml dosage.
When the pups were 18 days of age, five from each litter were randomly selected for testing. This was done to reduce the N to a manageable number. In order to qualify the pup's weight had to fall within ± 1 SD of the mean body weight of the litter.
The open-field test was conducted on Day 18 in a 48 cm × 48 cm Plexi-glas box lined with a cardboard sheet, marked with 7.6 cm squares. The animal was placed on the cardboard sheet facing the center of the box. The original placement was in one of the two center-most squares on either of the left or right edges of the box. The number of lines crossed within a 5-min. period was recorded along with a general outline of the animal's behavior. At the end of testing, the animal was returned to the litter and the cardboard sheet wiped clean with a damp sponge. This test was scheduled for the same time of day for each litter.
At 21 days the five selected pups from each litter were weaned and placed in individual cages. The first day of water-maze testing for each pup was scheduled for Day 22. The maze was constructed in the shape of two right turns followed by a left turn. The maze was 22.5 cm deep and constructed of 1.77 cm plywood. Each arm had an outside width of 14.6 cm. The first arm was 68.6 cm in length. The second arm, perpendicular to the first and completing the first T, was 63.5 cm long. The third arm was also 63.5 cm with the final arm being 59.7 cm in length. A 20.3-cm ramp was placed at the end of the last arm allowing the animal to climb out of the water (Mottin, 1967).
On testing days the maze was placed in a child's plastic swimming pool and filled with room temperature water to a depth of 12.7 cm. Placement of the maze in the outer pool allowed for the practical construction of the maze such that it could be less than watertight. Each pup was placed in the water at the beginning of the maze and timed as he traversed to the end, where he could climb up a ramp and terminate the noxious stimulation of the water. The use of a water maze for animals of this age was recommended by J. Mot-tin (Personal communication, 1973), who had earlier observed that this was a useful technique to assure that the animals persevered at the task and did not spend undue time exploring the maze. The criterion for mastery of the maze was defined as three consecutive error-free trials. Each animal was given a maximum of 10 trials on up to three consecutive days. (The first trial of Day 2 was Trial 11, etc.) Only one animal did not master the maze by the end of the third day. This performance was recorded as 30 trials to criterion. If an animal spent 5 min. in the maze without completing the trial or was judged to be in danger of drowning, it was removed and the performance for the given trial was recorded as 5 min. This procedure was employed in order to guard the animal against exhaustion. It had to be applied on only two occasions. A plastic sheet covered the maze during each trial in order to prevent the pup from climbing out. The number of trials to criterion, the average speed of swimming the first arm of the maze for each trial (speed), the total time to complete each trial, and the number of errors for each trial were recorded for each subject.
On the 36th day the pups were sacrificed by an overdose of ether and their brains were removed and weighed immediately. DNA determinations were conducted according to the method of Enesco and Leblond (1962). The only modification was in the final step when the DNA extract and diphenyl-amine reagent were incubated in the dark overnight (18 hr.) to develop color (Burton, 1956). The optical densities of the DNA extracts were read the next day at 595 mμ against the reagent blank in a Coleman spectrophotometer.
Results
Table 1 summarizes the major findings of this study. On the one hand, with respect to physical development, experimental pups weighed significantly less than the controls on Day 36. On the other hand, the brain weight of the experimental pups was significantly higher on Day 37.
Comparison of Experimental and Control Groups on Maternal Measurements, Pups' Growth Measurements, and Pups' Behavorial Measures
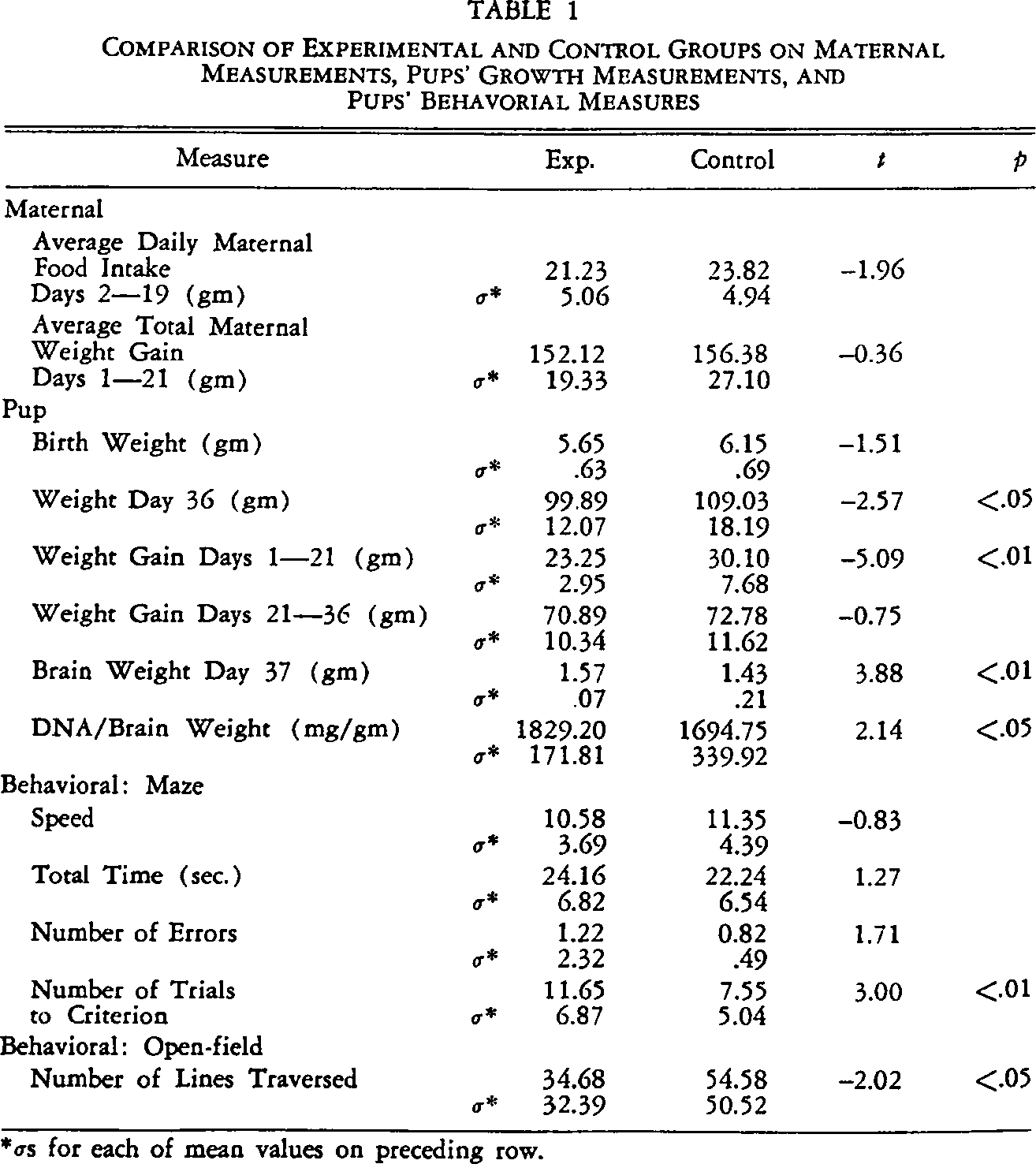
σs for each of mean values on preceding row.
Given the significantly higher brain weight for the experimental pups, all DNA assessments were divided by the corresponding brain weight to assure that any observed differences could not be accounted for by disparities in brain weights. The measures of DNA/brain weight were significantly higher for the experimental pups.
Examination of the behavioral measures reported in Table 1 shows that the control pups reached the criterion for learning the maze significantly faster than the experimental pups and traversed significantly fewer lines in the open-field task.
Inasmuch as it could be argued that the weight of the animals might influence performance in the maze, weight on the day of testing was correlated with each of the measures of learning. Weight did correlate significantly with the number of trials to criterion (r = −.38, p < .01) but not with any of the other measures.
Experimental and control pups differed in both weight and trials to master the maze. Weight correlated with number of trials to mastery of the maze. To assess whether the observed difference between experimental and control pups differed after removal of the effect of weight, the relationship between group membership and number of trials to criterion was first expressed as a point biserial correlation (r = .33, p < .01) which was then adjusted by means of a partial correlation. The correlation between membership in the experimental or control group and the number of trials to criterion remained significant but dropped from .33 to .19 when the effect of weight was partialled out (.05 < p < .10).
The significant difference in number of lines traversed in the open-field was also expressed as a point biserial correlation (r = −.23, p < .05). Again, to determine the effect of weight on day of testing, weight was correlated with open-field performance. This correlation was significant (r = .24, p < .05). Statistical removal of the influence of weight by means of partial correlation analysis reduced the correlation between treatment membership and number of lines traversed to −.14 (p > .10).
Discussion
Many of the physical effects of prenatal exposure to cannabis reported by Persaud and Ellington (1967, 1968) were not observed in this study. This may have been due to use of a different strain (e.g., Werboff, 1978). There were, nonetheless, four significant differences with respect to physical development: a difference in weight on Day 36, a difference in weight gain during Days 1-21, a difference in brain weight on Day 37, and a difference in the ratio of DNA to brain weight. The relative increase in weight of the control pups was not due to differences in litter size or food intake inasmuch as these were equal for the two groups. A more likely possibility is a difference in efficiency of utilization of food.
Since the direction of the differences in brain weight and DNA/brain weight ratio is contrary to what one might expect, it is important to bear in mind that the number of brain cells is not an indication of either emotionality or intelligence (Munn, Fernald, & Fernald, 1969). Inasmuch as cannabis is fat soluble, the greater brain weight may simply reflect storage of the agent. It is interesting to note that Carlini and Kramer (1965) reported that injection of marihuana to pregnant rats significantly increased brain DNA in their progeny.
With respect to the behavioral development of the pups, there were two statistically reliable observations. First, the experimental pups took a significantly greater number of trials to master the maze than did the control pups while manifesting similar scores on speed, total time, and number of errors. That these groups did not differ on speed (time to traverse the first arm of the maze) or total time (average time to complete a trial) suggests that the longer time that it took the experimental animals to master the maze was not due to any motor damage. On the other hand, there does seem to be some contribution of weight of the animals to mastery of the maze. The conversion of this finding to a point biserial correlation followed by an adjustment for the effect of weight by means of partial correlation showed a drop from .33 to .19. This latter figure is still significant, although marginally so. This suggests that both maternal exposure to cannabis and weight contribute, at least in a statistical sense, to the number of trials to criterion.
The exact significance of the role of weight in these findings is not clear but is intriguing. It appears as a significant covariate in the second statistically reliable behavioral observation: the difference in number of lines traversed in the open-field task. Removal of the influence of weight from the correlation between the animal's treated or untreated condition and its open-field behavior reduced the correlation from —.23 (p < .05) to —.14 (p > .10). In this case, the statistical correction for the influence of weight did reduce the effect of experimental treatment below the level of significance, suggesting that the increased emotionality among the experimental pups may not have resulted directly from the injection of cannabis. However, the significant correlation between the animal's weight and its experimental or control condition (r = −.48, p < .01) suggests that the cannabis resin influenced the weight of the animal, which in turn influenced performance through an effect on a mechanism presently measured indirectly by the weight of the animal.
These findings suggest that prenatal exposure to cannabis resin may, indeed, effect the behavioral development of offspring, although work will need to be done in order to spell out the mechanisms involved. The combined effects on learning, emotionality, and weight may reflect an effect on brain metabolism. Experimental work to date has suggested among other effects, exposure to cannabis and its derivatives can have an effect upon the hypothalamus, the hippocampus, and the amygdala, any or all of which might have contributed to the above observations (Drew & Miller, 1974; Miller & Drew, 1974).