
Abstract
Cognitive performance and cerebral hemispheric function are known to vary with fluctuating levels of estradiol and progesterone across the menstrual cycle in naturally cycling females. However, the literature is mixed with regard to how each hemisphere may be affected by elevated ovarian hormones. To better understand this, the current study employed a dual-task paradigm to examine potential shifts in hemispheric involvement for a verbal problem-solving task across the menstrual cycle in 30 right-handed, normally cycling young adult females (18–21 years old). To our knowledge, no study to date has utilized dual-task procedures to directly investigate the potential shifts in hemispheric function across the menstrual cycle. Specifically, participants were tested during both menses and their estimated midluteal phase where they engaged in repetitive unilateral finger-tapping while concurrently solving anagrams silently or aloud. Analysis of finger-tapping interference during the dual-task conditions revealed that solving anagrams silently was lateralized to the left hemisphere while solving anagrams aloud yielded a pattern of more bilateral hemispheric involvement, both of which were consistent across both menses and midluteal phases. Analysis of cognitive performance, however, revealed that silent anagrams performance while tapping with the right, but not left, hand significantly increased during the midluteal phase. Consistent with a number of other studies using different methodological approaches, the current dual-task findings suggest that when ovarian hormone levels are putatively elevated, there is enhanced recruitment of left hemisphere resources while performing a lateralized verbal task.
Keywords
Introduction
Sex differences on various cognitive tasks have been demonstrated across a considerable number of behavioral studies, with a female advantage observed for tasks involving language processing and a male advantage observed for tasks involving visuospatial processing (Andreano & Cahill, 2009; Hirnstein, Hugdahl, & Hausmann, 2019; Miller & Halpern, 2014). The reported cognitive sex differences are small to moderate, but most studies do not account for potential cognitive effects of circulating hormones (Hirnstein et al., 2019), which may contribute to increased variability and influence gender-specific outcomes in cognitive performance. For example, the menstrual cycle is one source of intraindividual variability in brain and cognitive function that may contribute to within-female variability in cognitive performance, which may influence the ability to detect and reliably capture the magnitude of potential cognitive sex differences.
The human menstrual cycle offers a natural paradigm to study activational effects of endogenous fluctuating ovarian steroids, such as estradiol and progesterone, on brain and cognitive function. However, the behavioral literature has generated mixed findings. For example, several studies have shown that normally cycling females demonstrate enhanced cognitive performance during phases of elevated estradiol and progesterone levels than during menstruation (when estradiol and progesterone levels are lowest) on various verbal tasks, including verbal articulation (Hampson, 1990), verbal fluency (Maki, Rich, & Rosenbaum, 2002; Solis-Ortiz & Corsi-Cabrera, 2008), and verbal memory (Maki et al., 2002; Rosenberg & Park, 2002). In contrast, enhanced cognitive performance during menstruation has been documented for various visuospatial tasks, such as mental rotation (Courvoisier et al., 2013; Hampson, Levy-Cooperman, & Korman, 2014; Hausmann, Slabbekoorn, Van Goozen, & Cohen-Kettenis, 2000; Maki et al., 2002; Phillips & Silverman, 1997; Silverman & Phillips, 1993), facial recognition (Mead & Hampson, 1996), spatial processing (Hampson, 1990), and spatial memory (Postma, Winkel, Tuiten, & van Honk, 1999). There are, however, a number of studies that did not detect menstrual cycle phase-related shifts in cognitive performance on various verbal or visuospatial tasks, which may be partially due to methodological differences and/or limited statistical power (for review, see Sundstrom Poromaa & Gingnell, 2014).
Given that verbal and visuospatial processing generally correspond to left and right (respectively) hemisphere specialization for the majority of individuals (Corballis, 2014; Pujol, Deus, Losilla, & Capdevila, 1999; Vogel, Bowers, & Vogel, 2003), fluctuations in verbal and spatial performance across the menstrual cycle may be partially mediated by concomitant cycle phase-dependent shifts in cerebral hemispheric activation. Behavioral studies examining hemispheric involvement for a given task across the menstrual cycle have employed divided visual field and/or dichotic listening techniques (Hausmann, 2017; Hodgetts, Weis, & Hausmann, 2015). These perceptual techniques manipulate the presentation of task stimuli to one or both hemispheres in order to generate perceptual asymmetries, with the assumption that task performance will be more proficient when stimuli are directly presented to the more appropriate hemisphere, indicating a functional cerebral asymmetry for that task (Hellige, 2000). Furthermore, changes across time in hemisphere-specific performance on a given task are assumed to infer changes in the involvement of one or both hemispheres for that task (Hausmann, Becker, Gather, & Gunturkun, 2002). Both behavioral and functional neuroimaging studies have shown enhanced left hemisphere activation for various verbal and visuospatial tasks during phases where estradiol and progesterone levels are elevated (Bibawi, Cherry, & Hellige, 1995; Cowell, Ledger, Wadnerkar, Skilling, & Whiteside, 2011; Dietrich et al., 2001; Fernandez et al., 2003; Hausmann et al., 2002; Hausmann & Gunturkun, 2000; Rode, Wagner, & Gunturkun, 1995; Sanders & Wenmoth, 1998; Wadnerkar, Whiteside, & Cowell, 2008; Weekes & Zaidel, 1996; but see Alexander, Altemus, Peterson, & Wexler, 2002; Weis et al., 2008). Studies have also demonstrated increased right hemisphere involvement for both verbal and nonverbal tasks during the menses phase (menstruation) when ovarian hormone levels are lowest (Cowell et al., 2011; Mead & Hampson, 1996; Sanders & Wenmoth, 1998).
These menstrual cycle-related differences in hemispheric activation may further reflect shifts in functional cerebral asymmetries (the differences between the left and right hemisphere for various cognitive processes). Indeed, some studies have demonstrated that hemispheric lateralization of verbal or spatial processing is greater during menstruation when ovarian hormone levels are lowest, whereas these asymmetries are reduced or eliminated when estradiol and progesterone are elevated (Hausmann, 2017; Hausmann et al., 2002; Hausmann & Gunturkun, 2000; Mead & Hampson, 1996; Sanders & Wenmoth, 1998). However, others have shown the opposite, with greater functional cerebral asymmetries observed during elevated ovarian hormone levels (Bibawi et al., 1995; Hausmann, 2017; Wadnerkar et al., 2008), or have shown no cycle effects (Can, Hahn, Ocklenburg, Ball, & Güntürkün, 2012; Hjelmervik et al., 2012). These disparate findings may partially depend on the type of cognitive task employed as well as test modality (Hausmann, 2017; Hjelmervik et al., 2012; , Hodgetts et al., 2015; Hodgetts, Weis, & Hausmann, 2017). For example, in dichotic listening studies where cognitive control demands are manipulated, normal cycling women tested during elevated levels of estradiol displayed improved cognitive control during a linguistic dichotic listening task (Hjelmervik et al., 2012; but see also Hodgetts et al., 2015, 2017) and slightly reduced cognitive control during an emotional prosody dichotic listening task (Hodgetts et al., 2017). These changes in cognitive control across the menstrual cycle were associated with a task-specific increase (Hjelmervik et al., 2012) or decrease (Hodgetts et al., 2017) in lateralization during elevated levels of estradiol. Furthermore, it has also been argued that task difficulty may influence the magnitude of menstrual cycle effects on lateralization, with more demanding tasks being more likely to reveal shifts in cerebral asymmetries across the menstrual cycle (Hodgetts et al., 2017).
Differences in cerebral functional asymmetries across the menstrual cycle may reflect increases or decreases in a single hemisphere’s activity or a concomitant change in both hemispheres (Hausmann, 2017). While there is evidence to suggest that hormonal effects act on a single hemisphere (Bibawi et al., 1995; Hausmann, 2017), it has also been proposed that high levels of ovarian steroid hormones may alter interhemispheric processes, which may ultimately lead to changes in functional cerebral asymmetries across the menstrual cycle (Hausmann & Gunturkun, 2000; Weis & Hausmann, 2010; Weis et al., 2008). For example, neuroimaging studies have shown that while completing a verbal task during periods of elevated ovarian hormones, the inhibition of the nondominant right hemisphere by the left hemisphere is significantly reduced and coincides with a concomitant decrease in functional cerebral asymmetry for that task (Weis & Hausmann, 2010; Weis et al., 2008).
Dual-task procedures are also useful for evaluating cerebral hemispheric asymmetries of verbal and spatial processing (Hellige & Kee, 1990; Kee & Cherry, 1990). In a dual-task paradigm, a hemisphere-specific manual activity (e.g., unilateral finger-tapping) is performed concurrently with a hemisphere-specific cognitive task. The fine motor movements of each hand (e.g., finger movements) used to perform the manual activity are controlled by the contralateral hemisphere. Thus, due to the sharing of the same cortical resources during the dual-task procedure, greater manual interference generally occurs with the hand controlled by the hemisphere that is more involved with the concurrent task (Hellige & Kee, 1990). For example, participants generally exhibit more right- than left-hand interference while performing a concurrent verbal task, suggesting asymmetric left hemisphere engagement (Kee, Bathurst, & Hellige, 1983; Kee & Cherry, 1990). However, if there is no change in the amount of manual interference, then an increase in cognitive performance associated with one hand over the other may also indicate the utilization of additional contralateral hemispheric resources (Hellige & Kee, 1990; Kee & Cherry, 1990).
To better understand how left hemisphere function may be affected by fluctuating ovarian hormones, the current study employed a dual-task paradigm to examine potential shifts in hemispheric involvement for a verbal problem-solving task across the menstrual cycle. To date, no study has utilized dual-task finger-tapping procedures to investigate the potential shifts in hemispheric function across the menstrual cycle, so the current study offers unique methodological insight. Within this dual-task paradigm, menstrual cycle-dependent shifts in the amount of hemispheric involvement for a given cognitive task may be indicated by shifts in the amount of left- or right-hand manual interference in the concurrent motor task or shifts in cognitive performance associated with one hand over the other in the concurrent verbal task.
Specifically, the current study employed dual-task procedures modeled after Kee et al. (1983) to assess shifts in left hemisphere involvement across estimated menses and midluteal phases of the menstrual cycle. These phases were chosen in attempt to maximize the difference in both estradiol and progesterone levels (Hausmann, 2005; Sundstrom Poromaa & Gingnell, 2014). In normally cycling women, estradiol and progesterone levels are typically lowest during the menses phase (menstruation); in contrast, the midluteal phase is typically marked by a second, more moderate peak in estradiol (the first larger peak having occurred just prior to ovulation during the late follicular phase) as well as a peak in progesterone (Hausmann, 2005; Sundstrom Poromaa & Gingnell, 2014). In the current study, participants were asked to engage in repetitive unilateral finger-tapping while concurrently solving anagrams silently or aloud during both phases. It was hypothesized that participants would show fluctuations in the magnitude of lateralized finger-tapping interference or cognitive performance across the two phases. Based on prior work (Bibawi et al., 1995; Dietrich et al., 2001; Hausmann et al., 2002; Hausmann & Gunturkun, 2000), we also hypothesized that potential shifts in lateralized finger-tapping interference or cognitive performance would be associated with increased left hemisphere involvement during the midluteal phase.
Method
Participants
A total of 30 female undergraduate students were conveniently recruited and participated in the current study to partially fulfill their introductory psychology course requirements. Participants were tested as part of a larger research project examining potential modulating factors of functional hemispheric asymmetries; those who met the inclusion criteria were included in the current study. Participants were 18 to 21 years of age (M = 18.87, standard deviation (SD) = 0.97), native English speakers, had normal or corrected-to-normal vision, and were right-handed. Left-handed and ambidextrous individuals were excluded due to increased variability in cerebral hemispheric lateralization of language in this subgroup compared to right-handed individuals (Pujol et al., 1999). During an initial screening, participants were first administered the Edinburgh Handedness Inventory (Oldfield, 1971) where they were presented with a list of 10 everyday activities and asked to indicate their hand preference when typically performing each activity. Scores on the Edinburgh Handedness Inventory, which can range from −100 (completely left-handed) to +100 (completely right-handed), confirmed that all participants were strongly right-handed (M = 93.93, SD = 8.82); those who scored lower than +70 were not included in the study. Participants also completed two 15-second unilateral finger-tapping tapping trials; participants who did not tap faster with their right hand on both trials were excluded from the study.
Participants who met the inclusion criteria for right-handedness then completed a brief menstrual cycle questionnaire to confirm the following additional inclusion criteria: a consistent menstrual cycle length varying no more than three days from cycle to cycle over the past year, no missed periods during the past year, no pregnancies within the past year, and no use of hormonal contraceptives or any other medication that would influence hormone levels for the past three months. All participants reported cycle lengths of between 28 and 31 days. During testing, two participants had cycle lengths that fell outside of this range, so their data were excluded.
All participants gave written informed consent after being given a full explanation of the study and were debriefed in person at the conclusion of their participation. The study was approved by the California State University, Fullerton Institutional Review Board.
Procedure and materials
Participants who satisfied all the inclusion criteria were individually tested on two sessions that coincided with their menses and midluteal menstrual cycle phases, which were chosen to capture the largest difference in both estradiol and progesterone levels (Hausmann, 2005; Sundstrom Poromaa & Gingnell, 2014). To schedule the testing sessions and confirm cycle lengths, participants agreed to inform the experimenter on the first day of their menstruation for two consecutive cycles. During testing, participants’ cycles ranged from 28 to 31 days (M = 29.13, SD = 1.31). Menses and midluteal phase length and timing estimates that were based on prior work (Asso, 1983; Hampson, 1990; Hausmann, 2005) were used to time each testing session. Specifically, menses phase testing sessions took place on day 3 to day 5 after the onset of menstruation, and midluteal phase testing sessions took place on day 20 to day 23 after menstruation. Because levels of testosterone are higher and are rapidly changing during the morning hours (Dabbs, 1990), participants were tested during the afternoon or early evening hours when testosterone levels were likely to have leveled off. The order of testing was counterbalanced across participants so that half of the participants began testing during the menses phase and the other half began testing during the midluteal phase.
On both testing sessions, participants were administered a dual-task paradigm task that consisted of finger-tapping while concurrently solving anagrams either silently or aloud (Kee et al., 1983). Anagram task stimuli were comprised of 16 different lists of four single-solution anagrams (e.g., the anagram “TYSLE” could be rearranged to form the word “STYLE”). Anagram solution words were chosen based on being previously rated for low imagery and medium-to-high familiarity (Gilhooly & Hay, 1977). High familiarity words were chosen to increase the probability that participants would recognize the word of the anagram solution. Each list of four anagrams was typed with a Times New Roman, 20-point font size in capital letters and was ordered vertically on a 22 cm × 28 cm sheet of white paper protected by clear cover sheets. A different anagram list was used for each dual-task trial (tapping while solving anagrams silently or aloud). For each participant, anagram lists 1–8 were presented in a random order during the first testing session, and lists 9–16 were presented in a random order during the second testing session. Thus, participants were presented with a total of 32 anagrams during each testing session where they were tasked with solving 16 anagrams silently and 16 anagrams aloud.
The tapping apparatus was a SiliconGraphics mouse, model M-UD43, which was mounted on a rectangular metal platform to prohibit lateral movement and to keep the tapping position constant for all participants. The tapping mechanism was run on a Dell Inspiron 5100 laptop computer using Tapstats, a program written specifically for recording finger-tapping measures in the lab. After a two-second tapping warm-up, Tapstats automatically records a participant’s mean number of taps per second derived from the number of taps performed during a 15-second trial period.
Participants were asked to complete a series of three tapping conditions on each hand: finger-tapping alone, finger-tapping while solving anagrams silently, and finger-tapping while solving anagrams aloud. Importantly, participants were instructed to limit their movement to only tapping the designated index finger during all trials. In the finger-tapping alone condition, participants were instructed to tap a computer mouse with either the left or right index finger as quickly and consistently as possible for 15 seconds while focusing on a white board positioned approximately 60 cm in front of them. In both dual-task conditions, participants completed the same tapping procedures while concurrently presented with a list of four different anagrams placed on a typing stand positioned 60 cm in front of them. In the anagrams silent condition, participants were asked to solve the anagrams silently while finger-tapping for 15 seconds. At the end of the 15-second trial, participants were asked to immediately verbalize the solutions to the anagrams that they just completed silently during the trial. The number of correct responses that were verbalized immediately after the trial was recorded for analyses. In the anagrams aloud condition, participants were instructed to solve each anagram aloud while finger-tapping for 15 seconds. To solve the anagrams aloud, participants were asked to verbally describe their steps while solving each anagram, such as verbalizing any transposition of letters in real time, sounding out a potential solution, and/or stating any guesses at the correct solution. Correct responses verbalized during the 15-second trial were recorded for analyses.
During each testing session, participants were given a brief series of practice trials, followed by the test trials. To familiarize participants with the tapping procedure, participants first performed two tapping alone trials (one for each hand, with hand order counterbalanced across participants), followed by one silent condition dual-task trial and one aloud condition dual-task trial (with hand assignment for practice dual-task trials counterbalanced across participants). After executing the practice trials, participants then completed a set of 8 test trials per hand for a total of 16 test trials: 4 tapping alone trials per hand (each paired with a dual-task trial), 2 anagrams silent trials per hand, and 2 anagrams aloud trials per hand. Importantly, each dual-task trial was paired with a tapping alone trial that served as the baseline tapping rate for that dual-task trial. Participants were administered the one of the two sequences of 16 test trials (consisting of eight consecutive trials per hand), which was counterbalanced across participants so that half of the participants experienced the first sequence and the other half experienced the second sequence for both testing sessions. In each sequence, the participants experienced the anagrams silent conditions before completing the anagrams aloud conditions. Tapping hand was counterbalanced so that half of the participants began the sequence with the right hand and the other half began with the left hand.
Analyses
For finger-tapping performance, tapping rates (measured in mean taps per second) were recorded for each trial. Percentage change scores in tapping rate for each dual-task trial paired with a tapping alone trial were then calculated as [(TA − D)/TA] × 100, where TA is the baseline tapping rate (tapping alone) and D is the tapping rate for the dual-task trial (Hellige & Kee, 1990; Kee et al., 1983). Thus, finger-tapping performance was assessed through tapping interference on dual-task trials, which was quantified by the mean percentage reduction from the baseline finger-tapping rate. Anagram performance was measured as the cumulative total number of correctly solved anagrams across all trials for each dual-task condition (silent and aloud).
Finger-tapping performance and anagram performance were analyzed separately, and the pattern of results for each was compared to rule out the possibility of a dual-task trade-off between the two measures and to investigate the possibility that variations in the cognitive measure may have occurred when tapping performance is held constant (Caroselli, Hiscock, & Roebuck, 1997; Hellige & Kee, 1990; Plummer & Eskes, 2015). To test for carryover effects, testing order (i.e., the phase during which participants were first tested) was initially included as a between-subjects factor in each analysis. However, for both finger-tapping and cognitive performance, there were no significant main effects of order and order did not significantly interact with any other variables. Thus, order was removed from further analyses and the data were pooled across the two counterbalanced testing order conditions.
Mean percentage change scores and total number of anagrams solved were subjected to separate 2 (Phase: menses vs. midluteal) × 2 (Hand: left vs. right) × 2 (Condition: silent vs. aloud) repeated measures analyses of variance (ANOVAs). Because we were primarily interested in evaluating the extent of right and left hemispheric involvement on this dual-task procedure across the menses and midluteal phases, separate planned comparisons between the left and right hands were performed within each phase as well as between menses and the midluteal phase within each hand. These comparisons were done separately for each dual-task condition (silent vs. aloud). An alpha level of .05 was set for all statistical tests. All analyses were performed using SPSS 24.0.
Results
Finger-tapping performance
Figure 1 presents the mean percentage reduction scores as a function of tapping hand and phase separately for each anagrams condition. The repeated measures ANOVA performed on finger-tapping interference revealed a significant main effect of Hand, F(1, 29) = 10.66, p = .003, η2 = 0.06, where participants had significantly larger percentage reduction scores overall while tapping with their right hand (M = 6.52, SD = 4.99) than while tapping with their left hand (M = 4.04, SD = 3.49). Thus, participants displayed an overall lateralized right-hand finger-tapping interference, indicating greater left hemisphere involvement overall for solving anagrams. There was also a significant main effect of Condition, F(1, 31) = 48.82, p < .001, η2 = 0.34, which was a large effect (Cohen, 1988) where tapping while solving anagrams aloud elicited more finger-tapping interference overall (M = 8.29, SD = 5.41) than tapping while solving anagrams silently (M = 2.27, SD = 3.20), suggesting that the added component of verbal production during the aloud condition may have required utilizing significantly more cortical resources overall (across both left and right hemispheres) than did solving anagrams silently while tapping.

Mean percentage reduction in tapping rate (tapping interference) for left and right hands while currently solving anagrams silently (a) or aloud (b) during the menses and midluteal phases. Error bars indicate standard error of the mean. *Significantly different from the corresponding menses or midluteal phase left-hand percentage reduction scores at p < .05.
Interestingly, there was a significant Hand × Condition interaction, F(1, 29) = 4.94, p = .03, η2 = 0.01, which was further examined via planned comparisons. During both menses and midluteal phase testing, participants had a significantly greater reduction in right-hand finger-tapping rate while concurrently solving anagrams silently than that of left-hand finger-tapping rate while concurrently solving anagrams silently (menses: p < .001, d = 0.79; midluteal: p = .013, d = 0.48; Figure 1(a)). In contrast, participants did not exhibit lateralized tapping interference while solving anagrams aloud during either menstrual cycle phase, as there were no differences in mean percentage reduction scores between the right and left hands for the aloud condition (Figure 1(b)). Thus, lateralized finger-tapping interference to the left hemisphere appears to only have occurred during the anagrams silent condition, while a pattern of more bilateral hemispheric involvement was observed for the anagrams aloud condition.
There was no main effect of Phase, F(1, 29) = 1.97, p = .28, η2 < 0.01, and Phase did not significantly interact with Hand or Condition (Fs < 1), indicating that tapping interference does not appear to vary as a function of menstrual cycle phase. Although there was a trend for greater left-hand tapping interference (i.e., increased right hemisphere involvement) while solving anagrams silently during the midluteal phase compared to menses (p = .06, d = 0.34), all planned comparisons between menses and midluteal phases in right- and left-hand tapping interference were not statistically significant for both the silent (Figure 1(a)) and aloud (Figure 1(b)) conditions.
Anagrams performance
Overall, participants correctly solved between 7 and 24 total anagrams during menses phase testing (M = 14.13, SD = 4.43) and between 7 and 22 total anagrams during midluteal phase testing (M = 13.80, SD = 4.05) across both silent and aloud trials. A repeated measures ANOVA on number of anagrams solved did not reveal a main effect of Phase, suggesting that anagrams performance overall was not altered by menstrual cycle phase. There was also no main effect of Hand or Hand × Condition interaction (Fs < 1). Importantly, the absence of these effects suggests that the main effect of Hand and the Hand × Condition interaction for tapping performance cannot be attributed to a dual-task trade-off with cognitive performance on the anagrams task. There was, however, a significant main effect of Condition, F(1, 29) = 6.79, p = .014, η2 = 0.02, which was a small effect (Cohen, 1988) where participants correctly solved significantly more anagrams across all aloud condition trials (M = 15.10, SD = 3.32) than across all silent condition trials (M = 13.63, SD = 3.13). Thus, the overall effect of Condition on finger-tapping performance may have been partially due to a dual-task trade-off with anagrams performance.
Figure 2 presents mean anagrams performance as a function of tapping hand and phase separately for each anagrams condition. The ANOVA for cognitive performance did reveal a significant Phase × Hand interaction, F(1, 29) = 6.38, p = .017, η2 = 0.03, which was further examined via separate planned comparisons for each anagrams condition. There were no significant differences in number of anagrams solved (silently or aloud) between the left- and right-hand dual-task conditions for either phase (Figure 2). Thus, in contrast to finger-tapping performance, cognitive performance associated with left- and right-hand finger-tapping did not appear to be strongly lateralized in either phase. Analyses further revealed that while tapping with the right hand, participants correctly solved significantly more anagrams silently during the midluteal phase than during the menses phase (p = .019, d = 0.45), which was a small-to-medium effect (Cohen, 1988). While tapping with the left hand, however, there was no difference between phases in number of anagrams solved silently (Figure 2(a)). In other words, anagram performance in the silent condition while tapping with the right (but not left) hand significantly increased during the midluteal phase. Because right-hand tapping interference during the anagrams silent condition was held constant across both menstrual cycle phases (see Figure 1(a)), these phase effects on silent anagram performance cannot be explained by a dual-task trade-off with tapping performance. Rather, these data suggest that additional left hemisphere resources were recruited during the midluteal phase (when ovarian hormone levels are elevated) while solving anagrams silently and concurrently tapping with the right hand. In contrast to the anagrams silent condition, menstrual cycle phase effects were not detected for the anagrams aloud condition, as there were no differences between phases in anagrams performance while tapping with the left or right hands (Figure 2(b)).
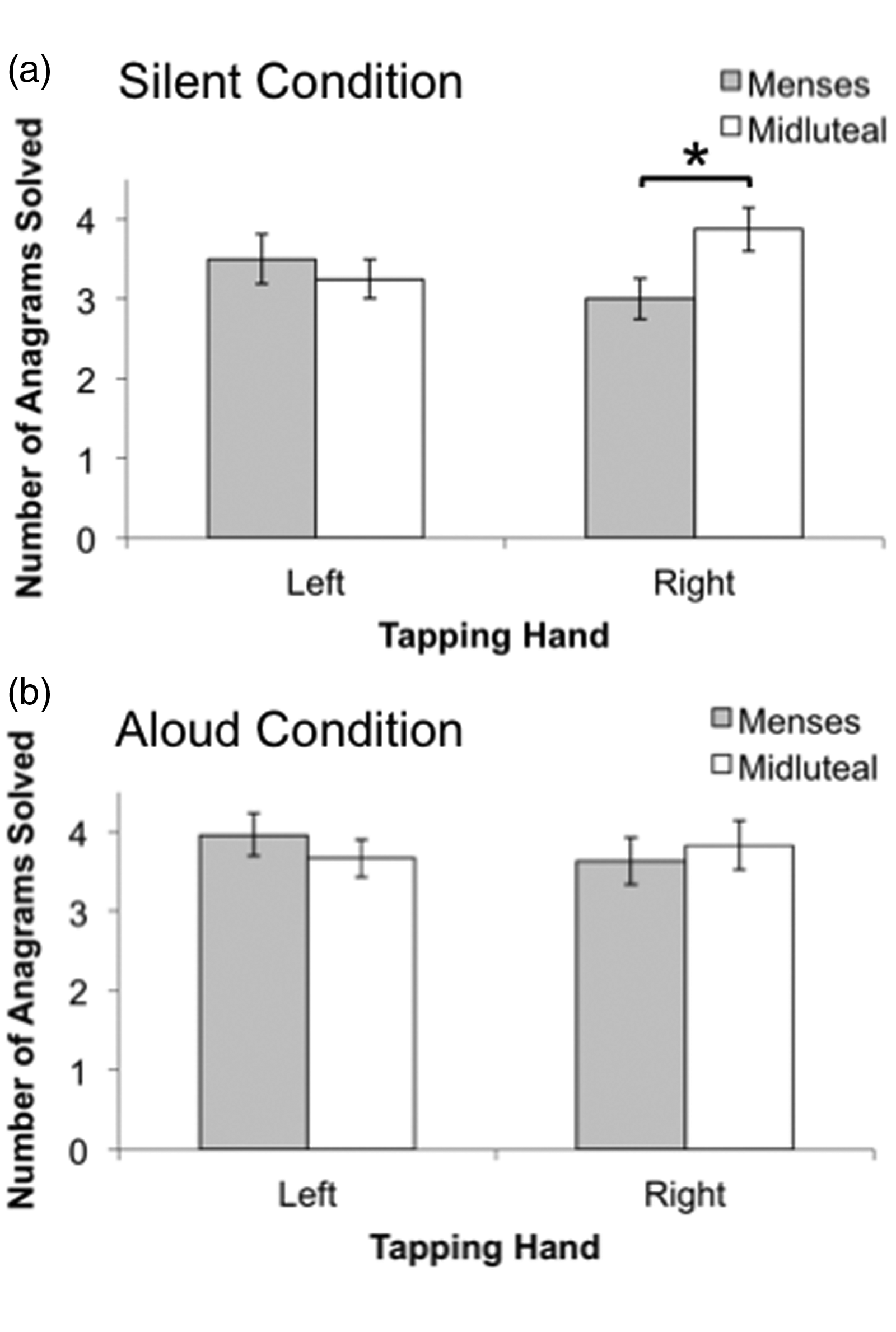
Mean total number of anagrams solved silently (a) or aloud (b) as a function of concurrent tapping hand (left vs. right) and menstrual cycle phase (menses vs. midluteal). Error bars indicate standard error of the mean. *Significantly different at p < .05.
Discussion
The current study employed dual-task procedures (unilateral finger-tapping while concurrently solving anagrams silently or aloud) to examine potential variation in cerebral hemispheric function across the menstrual cycle in normally cycling women. Specifically, hypothesized shifts in anagram-solving ability and associated cerebral hemispheric involvement were investigated across the estimated menses (low estradiol and progesterone) and midluteal (elevated estradiol and progesterone) phases of the menstrual cycle. These particular phases were chosen to maximize the differences in both estradiol and progesterone levels (Hausmann, 2005; Sundstrom Poromaa & Gingnell, 2014). While we did not detect shifts in finger-tapping interference or overall anagrams performance across the menstrual cycle, there was a modest increase in silent anagrams performance that was associated with right- (but not left-) hand finger-tapping during the midluteal phase compared to menses, suggesting an enhanced recruitment of left hemisphere resources for this verbal problem-solving task (while tapping with the right hand) during putative elevated estradiol and progesterone levels. This phase-related shift in left hemisphere activity was not detected for the anagrams aloud condition. Importantly, the current findings are, to our knowledge, the first to demonstrate the ability of this particular dual-task paradigm to detect shifts in hemispheric activation across the menstrual cycle.
It was initially hypothesized that shifts in left hemispheric activation across the menstrual cycle would be manifested by an increase in the magnitude of lateralized right-hand finger-tapping interference during the midluteal phase. However, analyses of finger-tapping performance did not support this hypothesis. Instead, finger-tapping interference results indicated that the pattern of tapping interference observed, while concurrently solving anagrams silently or aloud did not vary across the midluteal and menses phases. For both phases, greater overall finger-tapping interference was observed when anagrams were solved aloud than when anagrams were solved silently, which is consistent with previous dual-task studies demonstrating an increase in tapping interference (and thus an overall increase in utilization of cortical resources) when verbal production is required while solving anagrams (Kee & Cherry, 1990; Kee et al., 1986).
Surprisingly, however, the current study also revealed that during both menstrual cycle phases, lateralized tapping interference implicating left hemisphere specialization was observed only for the silent condition; in contrast, levels of finger-tapping interference while solving anagrams aloud were similar across both hands, suggesting more bilateral hemispheric involvement with the added verbal component while solving anagrams. This current finding partially contrasts with prior studies showing lateralized tapping interference for both the silent and aloud anagrams conditions (Kee et al., 1983; Kee & Cherry, 1990; Kee et al., 1986). The disparate results may be explained by differences in perception of task difficulty, in overall cognitive performance, and/or the inclusion of male subjects in prior studies. For example, when cognitive task complexity is increased, bilateral hemispheric processing becomes more advantageous for solving the task (Belger & Banich, 1992; Helton et al., 2010; Klingberg, O’Sullivan, & Roland, 1997), which also suggests that less demanding cognitive tasks may be more efficiently processed by a single hemisphere. In the current study, the added component of verbal production during the aloud condition may have enhanced the overall complexity of the task for these particular female participants; not only did participants have to engage in repetitive finger-tapping while concurrently solving anagrams, but they were also introduced with an additional cognitive demand of concurrently generating verbal expressions of their cognitive processes being used to solve the anagrams. Thus, similar levels of right- and left-hand finger-tapping interference during the aloud condition may have reflected a necessary recruitment of bilateral hemispheric processing during the more complex aloud verbal task, whereas the less demanding task of solving anagrams silently relied predominantly on the left hemisphere. In addition, unlike prior studies (Kee & Cherry, 1990; Kee et al., 1986), the current study revealed a small improvement in anagrams performance in the aloud condition compared to the silent condition, which may suggest a small dual-task trade-off where bilateral cortical resources were overallocated to the anagrams task at a small cost to finger-tapping performance (Plummer & Eskes, 2015).
Although menstrual cycle effects were not detected with the finger-tapping data, the hypothesized shift in hemispheric function across the menstrual cycle was detected by an analysis of cognitive performance in the anagrams silent condition. Importantly, the relative amount of hemispheric resources being utilized by the manual task of finger-tapping was held constant for each hand across both phases, as revealed by the absence of a Phase × Hand interaction for finger-tapping performance. Thus, any observed changes in anagram performance associated right- or left-hand finger-tapping could be attributed to the recruitment of additional hemispheric resources that enabled more efficient processing of anagrams and not explained by a dual-task trade-off with tapping performance (Hellige & Kee, 1990; Kee & Cherry, 1990).
The current study revealed that silent anagram performance associated with right-hand finger-tapping improved during the midluteal phase compared to the menses phase. In contrast, silent anagram performance associated with left-hand finger-tapping did not change across the two phases. This enhancement of silent anagram performance associated with right-hand finger-tapping during the midluteal phase likely stemmed from the recruitment of additional left hemisphere resources that were utilized while performing this particular dual-task condition. The silent anagrams task was lateralized to the left hemisphere (as revealed by the tapping interference data), which suggests that a sharing of left hemisphere resources was required when solving anagrams silently while concurrently tapping with the right hand (Hellige & Kee, 1990; Kee & Cherry, 1990). In contrast, silently solving anagrams while tapping with the left hand likely required minimal sharing of left hemisphere resources (Hellige & Kee, 1990; Kee & Cherry, 1990). The enhancement of silent anagrams performance during the midluteal phase occurred only for the dual-task condition that placed a higher demand on the left hemisphere (i.e., while tapping with the right hand), indicating that additional left hemisphere resources may have been recruited during the midluteal phase to lead to an improvement in cognitive performance on this particular dual-task condition. Thus, the cycle effects we observed in the current study were specific to solving anagrams silently while tapping with the right hand. It is possible that solving anagrams silently while tapping with the right hand also placed additional demands on cognitive control processes, which may have contributed to our ability to detect menstrual cycle-related shifts on this particular task condition (Hjelmervik et al., 2012; Hodgetts et al., 2017).
Importantly, the current dual-task findings corroborate other perceptual-behavioral studies that have also detected an enhancement of left hemisphere function during periods of elevated ovarian hormone levels for tasks involving verbal processing (Hampson, 1990; Sanders & Wenmoth, 1998; Wadnerkar et al., 2008; Weekes & Zaidel, 1996), as well as for nonverbal tasks (Bibawi et al., 1995; Hausmann et al., 2002; Hausmann & Gunturkun, 2000; Rode et al., 1995). The current findings are also consistent with neuroimaging studies that have detected increased left hemisphere activation during phases of elevated ovarian hormones for other types of verbal problem-solving tasks, including a word stem completion task (Dietrich et al., 2001) and a semantic decision task (Fernandez et al., 2003). Fernandez et al. (2003), however, reported increased neuronal activity in both the left and right hemispheres during the midluteal phase compared to menses, which is in contrast to the current study that demonstrated an enhancement in just left hemisphere involvement. Differences between the current study and Fernandez et al. may be partially explained by differences in the verbal task employed and the employment of dual-task parameters in the current study, which may have been less sensitive to changes in right hemisphere activity.
Interestingly, this menstrual cycle phase effect on left hemisphere function was only observed for the anagrams silent condition, which was also the condition that revealed lateralized tapping interference and indicated a left hemisphere specialization for solving anagrams silently. Most studies that demonstrate shifts in hemispheric activity across the menstrual cycle have used behavioral tasks that are generally lateralized to the left or right hemisphere (for review see Hausmann, 2017), which is consistent with the current findings on the anagrams silent task. In contrast, solving anagrams aloud while concurrently tapping yielded more bilateral hemispheric involvement in the current study, as demonstrated by similar amounts of tapping interference across both hands for the aloud dual-task condition. Thus, it appears that at least in the current study, the lateralized verbal task was more sensitive to changes in left hemisphere involvement across the menstrual cycle. However, one study using the divided visual field technique showed that the left hemisphere was more activated during the midluteal phase than during menses for a nonlateralized match-to-sample chair recognition task but not for a free-vision right hemisphere face processing task (Bibawi et al., 1995). This disparity between the current study and Bibawi et al. (1995) may be further explained by differences in task difficulty. As suggested by Hodgetts et al. (2017), studies that employ more demanding lateralized tasks appear to be more likely to reveal shifts in cerebral asymmetries across the menstrual cycle than that of studies that implement tasks with relatively low cognitive demands. In Bibawi et al. (1995), the face processing task, which required participants to make simple judgments about which of two chimeric faces looked happier, may not have been sensitive enough to detect changes in right hemisphere engagement across the menstrual cycle due to the task’s minimal cognitive demands on attention and working memory. In contrast, it is possible that the dual-task nature of solving anagrams silently while finger-tapping with the right hand placed sufficiently high enough cognitive demands (e.g., required more cognitive control) to observe a shift in left hemisphere involvement from the menses to midluteal phase. Taken together with previous studies, the current findings further highlight the complexity of menstrual cycle effects on hemispheric activity, highlighting that these cycle effects may largely depend on the type of cognitive task as well as task modality.
Because the current study focused on performance during the midluteal phase (when both estradiol and progesterone are elevated) relative to the menses phase, we were not able to make inferences about the individual effects of estradiol or progesterone on the measures analyzed. However, the current behavioral findings are consistent with Dietrich et al. (2001) who examined patterns of cortical activation associated with a lateralized motor and verbal task across the menses phase and preovulatory phase where only levels of estradiol were elevated. Using functional magnetic resonance imaging, Dietrich et al. showed that the magnitude of contralateral hemispheric activation of motor areas during a repetitive unilateral finger opposition task (touching each finger to the thumb in a repetitive sequence) did not vary across the menses and preovulatory cycle phase. While we did not examine motor or verbal performance during the preovulatory phase, the current study did find that the magnitude of lateralized tapping interference (i.e., motor task performance) while solving anagrams silently was similar across the menses and midluteal phases, which is consistent with Dietrich et al.’s findings that contralateral hemispheric activity supporting unilateral finger movements does not appear to fluctuate across the menstrual cycle. Moreover, Dietrich et al. also discovered that while females were solving a lateralized word stem completion task, the size of left hemisphere activity was significantly larger during the preovulatory phase than during menses. This finding is similar to the current dual-task findings that additional left hemisphere resources were putatively recruited while completing a lateralized anagrams task during elevated ovarian hormone levels. However, the findings by Dietrich et al. suggest that additional neural recruitment during verbal processing may be modulated by estrogen alone.
The main limitation of the current study was that we were unable to conduct hormonal assays to verify midluteal phase estimates in particular, which is why the inclusion criteria for normally cycling women that was implemented in the current study was very restrictive with regard to cycle length and regularity. However, using counting and self-report methods to accurately predict hormonal phases in young adult, women can be problematic due to variability in hormonal fluctuation between women, especially when predicting the preovulatory rise in estradiol levels (Blake, Dixson, O’Dean, & Denson, 2016; Hausmann, 2017; Wideman, Montgomery, Levine, Beynnon, & Shultz, 2013). It is worth noting, however, that our findings are consistent with other studies that also used midluteal phase estimates in the absence of hormone assays (Bibawi et al., 1995; Hampson, 1990; Sanders & Wenmoth, 1998) as well as with studies that did perform hormonal assays to verify midluteal phase estimates (Fernandez et al., 2003; Hausmann et al., 2002; Hausmann & Gunturkun, 2000; Rode et al., 1995). Future studies that include hormonal assays are needed to corroborate the current behavioral findings and to examine whether these menstrual cycle effects are also generalizable to women with less regular cycles and/or cycles that fall outside of the current study’s limited range as well as to dual-task performance during the late follicular phase when only estradiol levels are rising.
Similar to other menstrual cycle studies examining shifts in hemispheric activation (for review, see Hausmann, 2017), the menstrual cycle phase effects reported in the current study are relatively small to moderate in size as well as fairly specific in nature. However, it is worth noting that taken together with previous studies, the current findings demonstrate that the menstrual cycle is likely a source of intravariability in brain and cognitive function that may contribute to within-female variability in cognitive performance. Variability in brain function across the menstrual cycle may play a role in determining whether and to what extent cognitive sex differences exist, which may be particularly relevant for areas where findings on cognitive sex differences carry larger societal implications, such as job recruitment, hiring and promotion decisions, education approaches to learning, and so on (Hirnstein et al., 2019). Differences in hormonal states may influence overall cognitive sex differences, yet most studies examining cognitive sex differences do not account for potential hormonal fluctuations across the menstrual cycle in females (Hirnstein et al., 2019). Our current study findings offer additional evidence highlighting the importance of considering menstrual cycle effects as a potential source of variability in female cognitive performance.