
Abstract
A novel versatile glycoside hydrolase Cel5M was identified from Pectobacterium carotovorum and used in ramie degumming. The full-length Cel5M (GenBank: MH544226) containing the catalytic domain Cel5MD and its carbohydrate-binding module (CBM3) were expressed in E. coli. Both Cel5M and Cel5MD showed a broad substrate spectrum cleaving konjac glucomannan (KGM), carboxymethylcellulose (CMC) and oat spelt xylan. The optimal temperature of Cel5MD was 55oC with CMC and KGM, whereas the optimum temperature of Cel5M was 65oC with KGM and 60oC with CMC. The improved thermal activity with KGM was probably reflected also in the much lower K m with KGM than with CMC, indicating stronger binding of the full-length enzyme with KGM. Both enzymes were highly stable over a broad pH range from pH 5.0 to 9.0 with KGM. The use of the Cel5M enzyme together with the wild-type P. carotovorum HG-49 strain showed significant improvement when compared to using the mere strain HG-49 for application in ramie degumming. Moreover, the whole P. carotovorum Cel5M gene was over-expressed in P. carotovorum HG-49 itself to improve the ramie-degumming capacity. Altogether, this study indicates that in situ production of processing enzymes could be utilized in the textile industry.
Ramie (Boehmeria nivea) fiber has attracted increasing attention in recent years because of its sustainability, excellent hydroscopic property, special functional properties (comfort and anti-bacterial), biodegradability and exceptionally long single fiber range from 60 to 250 mm. 1 Ramie bast fibers (RBFs) contain 25–35% gum composed of amorphous cellulose, hemicellulose, pectin, lignin and other chemical components. Eco-friendly and textile-friendly degumming techniques are of major interest to improve the degummed ramie quality and economic feasibility, as well as to reduce pollution and protect the environment. 2 Since amorphous cellulose and hemicellulose in ramie fibers are heterogeneous components, the bio-degumming process requires the use of several enzymes to secure clean production and environmental protection. The degumming processes are generally carried out at a temperature range of 40–70℃ and alkaline pH (pH 8–11). 3
Pectinases, mannanases and xylanases have been studied in enzymatic ramie degumming. 3 Due to several fiber components in the gum materials, multifunctional enzymes are also an option for ramie degumming. One of the main multifunctional enzyme options are members of glycosyl hydrolase family 5 (GH5),4,5 including enzymes from Erwinia carotovora subsp. Carotovora LY34, Ruminococcus albus 8, Thermobifida fusca KW3, Thermotoga maritima and Bacillus subtilis 168, which randomly hydrolyze accessible internal β-1,4-glycosidic bonds of cellulose fibers. 6 Typically, the identification and biochemical characterization of GH5 enzymes suggests that this family shows hydrolysis ability against several substrates, such as carboxymethylcellulose (CMC), konjac glucomannan (KGM), locust bean gum (LBG), and xylan. 7 All GH5 enzymes share a common (α/β)8-barrel fold and retain the anomeric configuration at the cleavage site, while showing a diversity of protein sequences. 8 In addition, they usually exist as a single polypeptide chain containing non-catalytic accessory modules, such as carbohydrate-binding modules (CBMs). To date, more than 4000 different GH5 sequences have been released, and three-dimensional (3D) structures of 44 GH5 enzymes are available in the protein data bank (PDB) and show a very low sequence identity to each other. CBMs are generally classified into 84 families and non-classified sequences in the latest update of the CAZy database. 8 Their average size is in the range from 36 to 200 amino acids, and they play a crucial role in substrate recognition and hydrolysis by increasing affinity and proximity with the substrate to improve the hydrolysis capacity of the enzyme. 9 Furthermore, they increase the penetration abilities of the enzyme into the substrate. 10 CBM3, mainly used by bacteria, share as the dominant fold a β-sandwich structure composed of two β-sheets, and show affinity for soluble and insoluble cellulose. 11
Pectobacterium has been shown to be a promising bast fiber degumming microbe. 12 P. carotovorum subsp. Carotovorum (Pcc), classified previously as Erwinia carotovora subsp. carotovora (Ecc), is subordinate to the family of Enterobacteriaceae, and secrets several extracellular enzymes, including pectinases, proteases and cellulases, resulting in the degradation of plant tissue. Although the strain has been widely applied in bio-degumming and biopulping, it cannot completely replace conventional chemical degumming. Therefore, the most urgent technical challenge for improving degumming efficiency is to increase the secreted enzyme activity of P. carotovorum in the degradation of various gummy substances during the degumming process and, consequently, decrease the quantity of required degumming chemicals. 13
In this work, a novel glycoside hydrolase Cel5M gene from P. carotovorum HG-49 was cloned by utilizing the sequence information of the endoglucanase gene (accession no. CP001657.1; 2686424..2687941) from P. carotovorum subsp. carotovorum PC1. The cloned gene was then expressed in Escherichia coli BL21 (DE3). In addition, we performed a comprehensive biochemical characterization of Cel5M composed of an N-terminal GH5 catalytic domain including a C-terminal CBM3, and on the other hand, its single catalytic domain Cel5MD. Furthermore, homologous overexpression of the enzyme Cel5M in P. carotovorum subsp. carotovorum HG-49 was conducted for increasing the capacity of the strain to produce and secrete degumming enzymes for accelerating the ramie-degumming process, reducing the use of degumming chemicals and, therefore, lowering the negative environmental impact.
Materials and methods
Strains, vectors, media and chemicals
The P. carotovorum subsp. Carotovorum HG-49 strain was preserved at China Center for Type Culture Collection (CCTCC) under the accession number of M2017016. Escherichia coli Top 10 (Tiangen, China) and Escherichia coli BL21 (DE3) (Tiangen, China) were used as the host strains. The expression vectors pET-32a(+) and pUC18 of Escherichia coli were purchased from Novagen (Germany). RBFs and refined ramie fibers were obtained from Hubei Jinghua Textile Group Co., Ltd. All fiber samples were vacuum-dried at 60oC for 24 h before application. All other chemicals were of analytical grade.
Sequence information
A bioinformatics search of the P. carotovorum subsp. carotovorum PC1 complete genome sequence (accession no. CP001657.1) revealed only one gene for glycoside hydrolase family GH5 (accession no. ACT13369.1; 505 aa). The whole gene (1518 bp) includes one open reading frame (ORF) containing GH5 domain and an additional C-terminal region coding for CBM3. The signal peptide region was predicted by using the SignalP 4.1 server. 8
Cloning of different enzyme versions
The mature endoglucanase Cel5M gene (1431 bp) is from P. carotovorum subsp. Carotovorum HG-49 and the N-terminal region (915 bp) without CBM3 (named Cel5MD) were amplified using primers designed for these regions. The N-terminal region coding for the signal peptide was not included. Both genes of Cel5M and Cel5MD without the predicted signal peptide sequence (nucleotides 1–84) were separately cloned into the pET-32a(+) vector with a C-terminal His-tag using the ligation independent cloning technique. 14 Briefly, primers were designed to include 5′-Nco I and 3′-Hind III restriction sites to facilitate cloning. Recombinant plasmids were transformed individually into Escherichia coli BL21 (DE3) cells via heat shock, then plated and grown for 12 h at 37℃ on Luria–Bertani (LB) agar plates supplemented with ampicillin (100 µg/mL). A single colony from each transformation was inoculated into LB medium (10 mL) containing ampicillin (100 µg/mL) and grown to OD600 of about 0.5. Then, the DNA inserts in the recombinant plasmids were sequenced to confirm the correctness of the gene sequences by Tsingke Biotech (Beijing, China).
Modeling of Cel5M and Cel5MD
Protein modeling was done by using SWISS-MODEL. 15 The endoglucanase 5A (Cel5A, pdb entry 1e5j) was used as a template for the N-terminal domain (residues 7–305) of Cel5M and the family 3b′ CBM (pdb entry 2wob) was used as a template for the C-terminal CBM3 domain of Cel5M (residues 324–477). The PyMOL (2.7.6) program 8 was used for inspection of the protein structures and creation of the images.
Gene expression and protein purification
For gene expression, the two positive transformants were inoculated into 10 mL of LB medium supplemented with 100 µg/mL of ampicillin and grown at 37oC and 220 rpm for 6–8 h. The pre-cultures were then added to fresh LB medium (100 mL) with 5% inoculum size, supplemented with ampicillin (100 µg/mL) and grown at 37℃ and 220 rpm. At OD600 of 0.5, isopropyl β-D-thio-galactopyranoside (IPTG) was added to the culture at a final concentration of 0.1 mM. After 24 h of culturing, the cells and supernatants were collected by centrifugation (6000 rpm, 20 min, 4oC).
To purify the recombinant proteins with His-tag, Ni-chelating affinity chromatography was utilized according to the protocol of the manufacturer with a modified binding/wash buffer (20 mM Tris-HCl, 500 mM NaCl, 5 mM imidazole, pH 7.9) and elution buffer (20 mM Tris-HCl, 500 mM NaCl, 200 mM imidazole, pH 7.9). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was used to assess the protein purity. The protein bands were visualized by staining with Coomassie brilliant blue G-250. Purified proteins were quantified by measuring absorbance at 280 nm (OD280nm) using NanoDrop 1000 (Thermo Scientific, Waltham, MA).
Biochemical characterization of Cel5M and Cel5MD
Enzymatic activity assays were performed by a colorimetric method using the 3,5-dinitrosalicylic acid (DNS) method, 16 with glucose being a standard for the calibration curves. One unit (U) of enzyme activity was defined as the amount of enzyme releasing 1 micromole reducing sugars in 1 min reaction. The properties of optimal pH, temperature, pH stability and thermostability for Cel5M and Cel5MD were measured in triplicate experiments with 5 mg/mL medium-viscosity CMC or KGM as the substrate. For optimal pH, the reaction mixture containing 100 µL of enzyme solution, 900 µL of 5 mg/mL substrate in different buffers (50 mM Na2HPO4-citric acid buffer pH 4.0–7.5, 50 mM Tris-HCl buffer pH 7.5–9.0 and 50 mM glycine-NaOH buffer pH 9.0–10.0) was incubated at 50℃ for 10 min and stopped by adding 1.5 mL of DNS solution. Subsequently, the mixture was incubated for 5 min at 100℃ and the absorbance was measured at 540 nm with a spectrophotometer. The optimal temperature for endoglucanase activity was determined by measuring the activity at a temperature range of 40–80℃ in the optimal pH corresponding to each enzyme (Cel5M at pH 6.5 and Cel5MD at pH 6.0). To determine the thermostability, the enzymes were incubated at 37℃ and 50℃ in citrate buffer (Cel5M at pH 6.5 and Cel5MD at pH 6.0) for different periods of time (0, 5, 10, 20, 40, 60, 90 and 120 min). The pH stabilities were determined at 37℃ for each enzyme in various pH buffers, as described above, for 60 min. Then the residual activities were measured under optimal conditions.
The substrate specificity of the purified enzymes was investigated at the optimal conditions with the following substrates (5 mg/mL) in triplicate experiments: oat spelt xylan (Sigma X-0627, USA), sodium carboxymethyl cellulose (CMC-Na, Sigma 21902, USA), LBG (Sigma 91783, USA), KGM (Aladdin, Shanghai, China), guar gum (Aladdin, Shanghai, China) and filter paper (Aladdin, Shanghai, China). The substrate specificity assays were done with the assay conditions described above (pH assays) at 50oC and pH 7.0. The same amount of enzyme was used for each substrate. The kinetic parameters were determined after measuring the initial rates at various concentrations (1–8 mg/mL) of CMC and KGM under optimal conditions for 3 min. The Michaelis–Menten constant (K m ) and the maximum velocity (Vmax) were calculated using Lineweaver–Burk plots. The unit of K m is mg substrate/mL and the unit of Vmax is micromole glucose produced/min/mg protein. The specific activity is defined as U/mg enzyme.
Homologous overexpression of Cel5M
The gene encoding Cel5M (1518 bp) equipped with terminal 5′-EcoR I and 3′-Hind III restriction sites was amplified from P. carotovorum HG-49 and, subsequently, was cloned into the expression vector pUC18. Recombinant plasmid was transformed into P. carotovorum HG-49 cells via electroporation (Gene Pulser Xcell™ Electroporation System, Bio-Rad, USA), plated and grown for 16 h at 37℃ on LB agar plates supplemented with ampicillin (100 µg/mL). A single colony from each transformation was inoculated into LB medium (10 mL) containing ampicillin (100 µg/mL) and grown to about OD600 of 0.5. Then, the foreign DNA insert in the recombinant plasmid was sequenced to confirm the correctness of the gene sequence by Tsingke Biotech (Beijing, China). The positive transformant Cel5M-HG-49 and the wild-type HG-49 were inoculated into 10 mL of LB medium and grown at 37oC and 220 rpm for 6–8 h. The pre-cultures were then added to fresh LB medium (100 mL) with 5% inoculum size and grown at 37℃ and 220 rpm for 16 h. Finally, the fermentation supernatant was collected by centrifugation (6000 rpm, 20 min, 4oC). Enzymatic activity assays were performed by a colorimetric method using the DNS method and the growth of cell mass was determined every 2 h.
Assays on ramie degumming
The enzymatic degumming with Cel5M was carried out in a 250 mL flask containing 5 g of RBFs at pH 7 and 37℃ under 180 rpm of shaking frequency for 16 h with purified Cel5M in 60 mL of deionized water with the final activities of 0, 100, 200, 300, 400 or 500 IU/mL. The microbial and enzymatic degumming of 5 g of RBFs was performed also with 10 mL (OD600 = 1.8, about ∼109 cells per ml) of 6 h-old inoculum of wild-type HG-49 and 50 mL of deionized water with purified Cel5M for the final activities of 0, 100, 200, 300, 400 or 500 IU/mL.
The microbial degumming was carried out in a 250 mL flask containing 5 g of RBFs and 10 mL of 6 h-old inoculum of strain HG-49 or Cel5M-HG-49 to about ∼109 cells per milliliter with 50 mL of deionized water at pH 7.0, 37℃ and 180 rpm for 16 h. All experiments were run in triplicate. Glucomannanase and carboxymethylcellulase (CMCase) activities were determined during the process of ramie degumming every 2 h.
To evaluate the degumming ratio, the treated fiber sample was beaten with a rubber covered mallet while flushing with a large amount of water to remove the gum that has been degraded or loosened in the degumming process but still attached to the fibers. Then the fiber sample was vacuum-dried at 60℃ to a constant weight to gravimetrically determine the gum contents of the fibers. 3
Microscopy
The degumming efficiency evaluation were carried out as described by Fan et al. 3 The surface morphology and microstructure of ramie fibers were observed utilizing an FEI-Quanta200 scanning electron microscope (FEI, Hillsboro, OR, USA) at 20 kV.
Nucleotide sequence accession number
The nucleotide sequence encoding the glucomannanase Cel5M was deposited at the GenBank database under accession number MH544226.
Results
Sequence analysis
The cloned full-length gene (1518 bp) of Cel5M showed the highest nucleotide sequence identity (99%) to the glycoside hydrolase family GH5 gene from P. carotovorum subsp. carotovorum PC1 (accession no. CP001657.1; 2686424..2687941) and was predicted to encode a protein of 505 amino acids (aa). The deduced amino acid sequence (477 aa; Ala29 to Pro505) without the signal peptides (28 aa; Met1 to Ser28) showed the highest identity with several published translated nucleotides, such as 99% identity with endo-1,4-beta-glucanase celA from Erwinia carotovora subsp. carotovora LY34 (accession no. AAC02964.2), 97% identity with cellulase CelV from Erwinia carotovora subsp. carotovora (Ecc) SCRI193 (accession no. Q47096.1) and 97% identity with beta-glucanase CelN from Erwinia carotovora subsp. atroseptica (Eca) C18 (accession no. AAC37033.1) (Figure 1). Therefore, Cel5M appears to be an endoglucanase-like enzyme, but the further analysis reported in this study shows that it is rather a versatile glycoside hydrolase with main activity on the glucomannan substrate KGM.
Multiple alignment of protein sequences of glycosyl hydrolase family 5 (GH5) endoglucanases. The numbering of each sequence starts from the initiation amino acid. Identical amino acids are indicated by black shading. The candidates of endoglucanase include endo-1,4-beta-glucanase celA (AAC02964.2) from Erwinia carotovora subsp. carotovora LY34, cellulase CelV (Q47096.1) from Erwinia carotovora subsp. carotovora (Ecc) SCRI193 and beta-glucanase CelN (AAC37033.1) from Erwinia carotovora subsp. atroseptica (Eca) C18.
Sequence analysis indicates that the mature Cel5M (477 aa) contains an N-terminal GH5 domain (305 aa; Ala29 to Asn333) and a C-terminal CBM3 domain (154 aa; Gly352 to Pro505) with a linker between them (18 aa; Leu334 to Ser351). The predicted protein with the two domains shows a deduced molecular mass of 51.8 kDa and a calculated pI of 4.97. In addition, the constructed Cel5MD enzyme contains only the GH5 catalytic domain (305 aa; Ala29 to Asn333) with the same sequence as the cloned full-length Cel5M, and it has a theoretical molecular mass of 33.3 kDa and a calculated pI of 5.14.
Structural modeling
The catalytic domain Cel5MD shares 70% sequence identity with the catalytic domain identified in endoglucanase Cel5a from Bacillus agaradhaerens strain AC13 (NCIMB 40482) (PDB structure 1E5J_A).
17
The 3D structure of Cel5MD was modeled by homologous modeling with 1E5J_A as a template, and the results confirmed that Cel5MD is formed of an (α/β)8-barrel fold. The eight conserved amino acids of endoglucanase Cel5MD include Arg63, His102, Asn139, His200, Tyr202, Trp262 and the two catalytic residues Glu140 and Glu228, which belong to the structural features of the GH5 domain (Figure 2(a)). In addition, the 3D structure of CBM3 from the C-terminal region of Cel5M was modeled based on the family 3b′ CBM (PDB structure 2WOB_A) with a high GMQE (Global Model Quality Estimation) of 0.73, although CBM3 shares only 35% identity with the 3b′ CBM from the Cel9V glycoside hydrolase from Clostridium thermocellum YS (Figure 2(b)).
18
The 154 residues of CBM3 form a nine-stranded anti-parallel β-sandwich, and the ‘bottom’ β-sheet of the β-sandwich is formed by strands 1, 2, 7 and 4, while the ‘top’ β-sheet comprises relatively short anti-parallel strands 5, 6, 3, 8 and 9 (Figure 2(b)). In conclusion, the 3D structure of Cel5M predicted by modeling indicated that the N-terminal domain is formed of a (α/β)8-barrel fold and the C-terminal is formed of a β-sandwich containing two anti-parallel sheets of four and five β-strands, respectively (Figure 2(c)).
Overall structure of CelMD, CBM3 and CelM. CelMD and CelM are in complex with ligand molecules. Secondary structures are color-coded with strands in salmon, helices in pale cyan and random coils in purple. The ligand molecules, methyl-4II-s-alpha-cellobiosyl-4II-thio-beta-cellobioside, transferred from the 1E5j structure are shown in ball-and-stick, in yellow and gray. The two putative catalytic residues, Glu140 (acid/base) and Glu228 (nucleophile), are shown in blue. Schematic representation of the structure of the catalytic domain Cel5MD (a); schematic representation of the structure of CBM3 is shown with the β-strands labeled (b); schematic representation of the structure of full-length enzyme Cel5M consisting of the catalytic domain and CBM3 (c). (Color online only.)
The analysis of the modeled structure with ligands transferred from 1E5j confirmed the expected catalytic site and catalytic residues (acid/base Glu140 and nucleophile Glu228). The distance between the catalytic residues (4.5 Å) indicates that the enzyme functions by the retaining mechanism, as is also shown for the enzyme used as the template. 17 The catalytic canyon is quite shallow, which might be a reason for flexibility in respect to accepted substrates, especially when glucose-, mannose- and xylose-based polysaccharides have the same lengths in the polysaccharide backbone. Phylogenetic analysis indicated that dual endoglucanase–mannanase activity is widespread in the GH5 family of glycoside hydrolases. 5 Also, xylanase activity is included in GH5 enzyme activity profiles, making it a versatile enzyme when wide substrate specificity is required.
Gene expression and protein purification
The glycoside hydrolase Cel5M and its catalytic domain Cel5MD were secreted respectively by E. coli and purified with the His-tag method. The purified Cel5M and Cel5MD revealed apparent molecular mass of ∼70 and ∼51 kDa by SDS-PAGE, respectively (Figure 3), which was in good agreement with the predicted molecular weights from their amino acid sequences (∼52 and ∼33 kDa, respectively) fused with the peptide sequence (∼18 kDa) from the expression vector pET-32a(+).
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis of Cel5M and Cel5MD. Lane 1: purified Cel5M. Lane 2: purified Cel5MD. Lane M: molecular marker.
Biochemical characterization
The temperature-dependent activity was measured by using CMC as the substrate (Figure 4(a)). The full protein Cel5M exhibited optimum activity at 60℃ in contrast with the optimum activity of the mere catalytic domain Cel5MD at 55℃, indicating that the CBM3 domain increases the thermal performance of the enzyme. Both enzymes maintained more than 50% of their maximal activity in the range of 40–65℃ (Figure 4(a)).
Optimal reaction temperature and thermostability of Cel5M and Cel5MD. Optimal reaction temperature of Cel5M and Cel5MD toward carboxymethylcellulose (a); optimal reaction temperature of Cel5M and Cel5MD toward konjac glucomannan (KGM) (b); stability of Cel5M and Cel5MD as a function of time (assayed after incubations toward KGM) (c). All experiments were done at pH 7.0.
The temperature effect of the substrate on glucomannanase activity was demonstrated with KGM (Figures 4(b) and (c)). The results showed that the substrate KGM stabilized Cel5M and Cel5MD more strongly than CMC. KGM contains mannose and glucose in a molar ratio of 1.6:1.0 linked together by β-1,4 linkage. KGM contains also some β-1,3-linked branches and acetyl groups. 19 The enzyme was clearly more active on β-1,4-glucomannan (KGM) than β-1,4-galactomannan (LBG, guar gum). This was seen as higher activity in about the 65–80oC region, with both enzymes, and also as a higher temperature optimum (65oC) with KGM as the substrate for Cel5M when compared to CMC as the substrate (60oC). The effect of KGM was stronger with CBM, indicating that binding of the substrate to CBM3 increases the thermostability of the catalytic domain. Cel5M maintained more than 68% of its maximal activity in the range of 50–75℃. Although the optimal temperature for the enzyme Cel5MD activity was also found to be 55℃, the higher activity of Cel5MD above the temperature optimum was seen in the maintaining more than 66% of its maximal activity in the range of 40–70℃ (Figure 4(b)).
The investigation of the thermal stability showed that Cel5M and Cel5MD lost slightly the activity at 37℃ and 50℃ during 120 min incubation, except the mere catalytic domain (Cel5MD), which retained the best the stability during 120 min incubation at 37oC, but not at 50oC (Figure 4(c)). This indicates that the CBM3 domain decreases the stability of the full protein at 37oC but not at higher temperatures, in which CBM3 stabilizes the enzyme.
According to the results of the optimum pH assays (Figure 5), we found that Cel5M and Cel5MD showed catalytic activity in the pH range of 4.0–10.0, and their highest catalytic activity was observed at pH 6.5 and 6.0 in 50 mM sodium-citrate buffer, respectively (Figures 5(a) and (b)). This demonstrated that CBM3 had only a very small impact on the enzyme's optimum pH value. The two enzymes showed higher activity in the range of pH 4.0–9.0 toward KGM than CMC (Figure 5(a) and (b)), and were significantly stable in the range of pH 5.0–9.0 with KGM (over 82% activity) (Figure 5(c)). The stability ranges of Cel5M and Cel5MD were also similar.
Effect of pH on activity and stability of Cel5M and Cel5MD. Effect of pH on the activity of Cel5M and Cel5MD toward carboxymethylcellulose (a); effect of pH on activity of Cel5M and Cel5MD toward konjac glucomannan (KGM) (b); pH stability of Cel5M and Cel5MD toward KGM (c).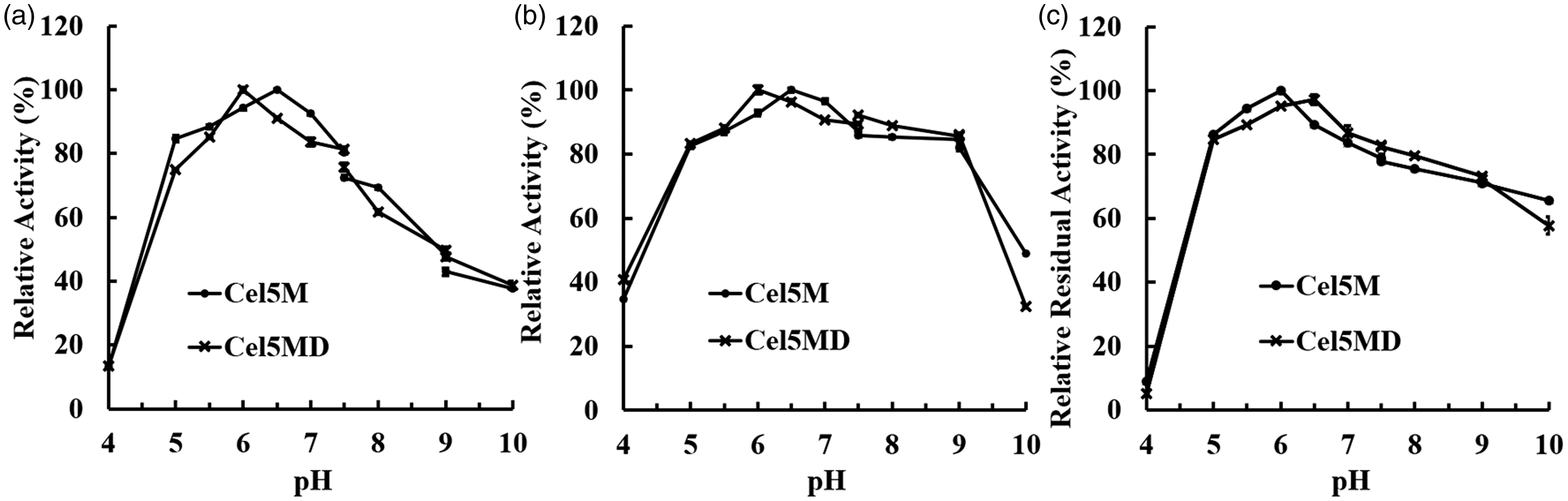
Effects of various chemicals and metal irons on Cel5M and Cel5MD toward activity with carboxymethylcellulose as the substrate
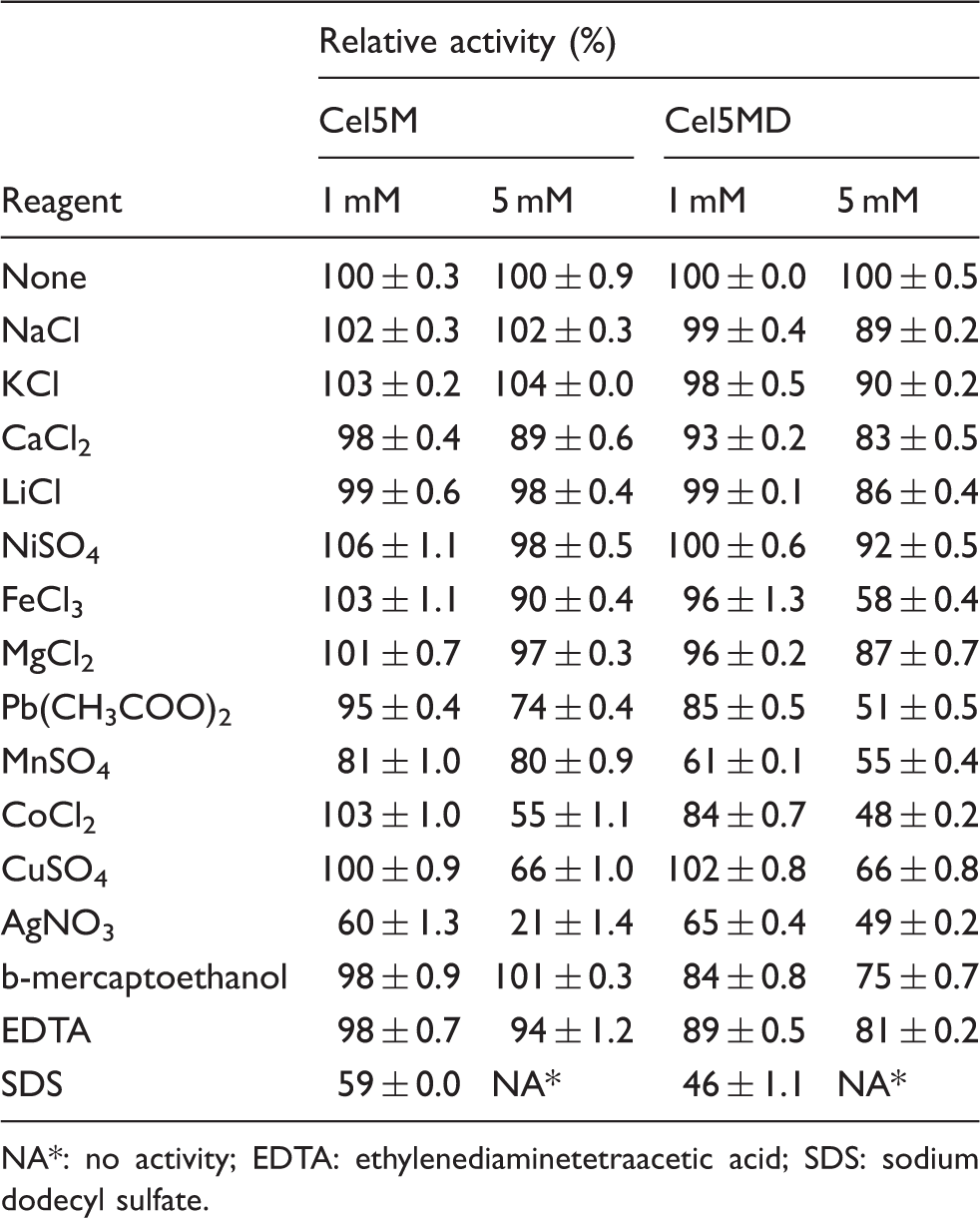
NA*: no activity; EDTA: ethylenediaminetetraacetic acid; SDS: sodium dodecyl sulfate.
Substrate specificity of Cel5M and Cel5MD

NA: no activity; KGM: konjac glucomannan.
The activity profiling of Cel5M and Cel5MD toward various substrates suggested that they react with a wide range of soluble substrates with β-1,4-glycosidic linkages. The enzymes displayed the highest activity toward KGM, followed by CMC, xylan, guar gum and LBG (Table 2). The high catalytic activity of Cel5M toward non-crystalline cellulose and hemicellulose make it a potential candidate for industrial application in bioscouring processes, such as ramie degumming.
Homologous overexpression of Cel5M
The recombinant strain Cel5M-HG-49 was created to increase the expression of the Cel5M enzyme from the HG-49 strains. The full-length enzyme Cel5M (1518 bp) containing the signal peptide was used in the cloning. The constructed recombinant vector Cel5M-PUC18 was transformed into strain HG-49 by electroporation. Thereafter, a positive recombinant strain named as Cel5M-HG-49 was successfully screened out and then cultivated in LB medium containing ampicillin (100 µg/mL) to check the enzyme secretion by the recombinant cells. Figure 6 shows the changes in cell concentration, glucomannanase activity and CMCase activity between strain HG-49 and engineered strain Cel5M-HG-49 over the liquor fermentation time of 16 h. Both strains showed similar growth curves. The cell concentration increased clearly during the exponential growth phase (2–8 h), the growth stayed on a plateau from 8 to 12 h and then decreased after 12 h, coming to a cell death period (Figure 6(a)). The glucomannanase activity increased with the increase of cell mass and reached the maximum at 8 h of fermentation time. The secreted glucomannanase activity by strain Cel5M-HG-49 was 41.2 U/mL higher than that by the original strain HG-49 (28.3 U/mL). Meanwhile, during the entire fermentation process, CMCase activity showed very slow growth for strain HG-49 but showed a remarkable growth profile for strain Cel5M-HG-49. After 16 h of fermentation, CMCase activity by strain Cel5M-HG-49 was 10.3 U/mL, whereas that by the strain HG-49 was extremely low (0.1 U/mL) (Figures 6(b) and (c)). These results showed that the recombinant strain overexpressing the GH5 enzyme has much improved capacity compared to the wild-type strain to produce and secrete the enzyme activity needed in the degumming process.
Growth of cell mass, glucomannanase and carboxymethylcellulase (CMCase) activity of strains HG-49 and Cel5M-HG-49 during 16 h in a shaker flash culture with Luria–Bertani medium. The cell mass of strains HG-49 and Cel5M-HG-49 (a); glucomannanase activity of strains HG-49 and Cel5M-HG-49 (b); CMCase activity of strains HG-49 and Cel5M-HG-49 (c).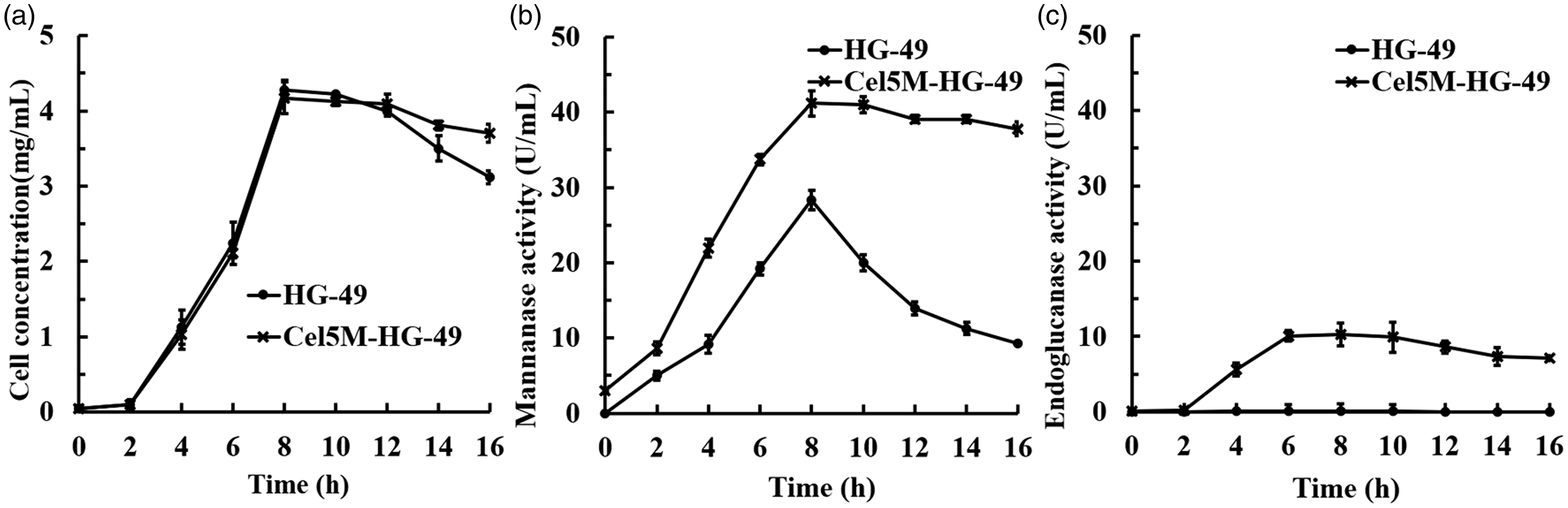
Application of Cel5M and strain Cel5M-HG-49 in ramie degumming
For an environmentally friendly degumming method, the primitive retting process has inspired people to screen the effect of microorganisms and their enzymes on ramie degumming. 20 After that, an increasing number of polysaccharide-degrading enzymes were investigated in the bio-degumming process. 21 The major hemicelluloses of ramie fibers were reported to be galactoglucomannan, glucomannan and xylan. 22 Therefore, hemicellulose-specific enzymes can be used in degumming. The efficiency of P. carotovorum in a microbial ramie-degumming process was highly satisfactory with expressed high activities of pectinase, mannanase and xylanase, and clearly revealed improved loosening of the fibers following the microbial treatment. However, there was still residual pectin, hemicellulose and non-crystalline cellulose on ramie fibers after bio-degumming by P. carotovorum. 23 Due to this finding, a comparison study was started to evaluate the effect of adding separately produced Cel5M enzyme into the process of microbial ramie degumming by strain HG-49. Based on the positive results, an overexpression recombinant strain was constructed secreting the Cel5M enzyme to the medium.
In our study, the ramie-degumming process was alkalic: the pH of the solution containing RBFs with single strain HG-49 was altered from 7.0 to 9.3 during the entire ramie-degumming process. The ramie-degumming processes utilizing the enzyme Cel5M alone or in combination with the degumming microbe were performed at pH 7 and 37℃ for 16 h. The gum content reduction was measured to be 10.1%, 13.1%, 13.7%, 13.7%, 14.2% and 14.9% for ramie fibers treated by mere Cel5M at concentration of 0, 100, 200, 300, 400 and 500 U/mL, respectively. A 10.1% gum content reduction was obtained for the negative control, which consisted of ramie fibers treated with only deionized water at pH 7, and the use of Cel5M increased the removal maximally only to 14.9%. This result shows that Cel5M alone caused only a partial removal of the gums. The respective weight reduction of ramie fibers was 82.2%, 84.5%, 85.1%, 86.0%, 88.9% and 87.3% by mixture of the consistent amount of strain HG-49 (10 mL, OD600 = 1.6, about 3.6*109 cells per mL) and added Cel5M at concentrations of 0, 100, 200, 300, 400 and 500 U/mL, respectively. The maximum achieved weight loss of ramie fibers treated by the enzyme-microbial method was 88.9% for the Cel5M concentration of 400 U/mL. The minimum gum removal ratio was 82.2% with strain HG-49 alone in the degumming process. The improvement by the addition of Cel5M enzyme into the enzyme-microbial degumming process was about 6.7%, which was higher than that affected by Cel5M alone.
The gum content reduction of RBFs was 87.7% after degumming by the engineered recombinant strain Cel5M-HG-49 at pH 7.0, 37℃ and 180 rpm for 16 h, which is higher than 82.2% with strain HG-49 alone. Since Cel5M was active at pH 5–9 and stable at pH 5–10, it is well suited to the conditions of ramie degumming, in which the pH is moderately alkaline. The results indicated that the added enzyme Cel5M improved the ramie degumming by strain HG-49, and quite similar results were obtained when the additional Cel5M enzyme was introduced into the degumming process by expressing it from the Cel5M-HG-49 strain.
Moreover, during the degumming process by strain HG-49 or Cel5M-HG-49, the cell concentration increased gradually during the first 2 h, and then reached the maximum concentration (10.8 × 109 and 14.4 × 109 CFU/mL, respectively) at 8 h, and finally slowly decreased to 8.4 × 109 and 10.8 × 109 CFU/mL, respectively, at 16 h. Both glucomannanase and endoglucanase (CMCase) activities increased during the degumming and achieved the maximum enzyme activity after 10 h. The highest glucomannanase activity was 67.8 and 79.7 U/mL for HG-49 and Cel5M-HG-49, respectively. The highest CMCase activity was 16.7 and 19.8 U/mL for HG-49 and Cel5M-HG-49, respectively. After 10 h, all activities of the two enzymes started to drastically decrease up to 16 h (Figure 7). The results indicated that the recombinant strain Cel5M-HG-49 showed a higher advantage in ramie degumming based on the higher cell concentration, glucomannanase activity, CMCase activity and gum content reduction than with the mere HG-49 strain. The effect of additional Cel5M expression is probably caused by the hydrolysis of the remaining linkages between hemicellulose and cellulose, pectin and lignin and, thus, by removing more hemicellulose and non-crystalline cellulose.
Growth of cell mass and glucomannanase and carboxymethylcellulase (CMCase) activity of strains HG-49 and Cel5M-HG-49 during ramie degumming of 16 h in shaker flash fermentation. Cell mass of strains HG-49 and Cel5M-HG-49 (a); mannanase activity of strains HG-49 and Cel5M-HG-49 (b); CMCase activity of strains HG-49 and Cel5M-HG-49 (c).
Electron microscopy
Scanning electron microscope studies revealed the changes in the morphology of ramie fibers after degumming when compared with the untreated ramie fibers (Figure 8). A large amount of gummy substance adhered to the surface of untreated ramie fibers (Figure 8(a)), whereas the degummed ramie was well-separated and almost all gummy substances were removed from the surface of the fiber bundle and the surface became clear and smooth (Figure 8(b)). Moreover, partial gummy substances were removed but the fiber bundle was dispersed insufficiently with purified Cel5M (500 U/mL) alone (Figure 8(c)). Although the fiber bundles were well-separated, some gummy substances still adhered to the bundle surfaces with the mere HG-49 strain (Figure 8(d)). The fiber bundles were well-separated and small amounts of gummy substances remained on their surfaces, and the exposed surfaces of the fiber bundles were much smoother with the mixture of strain HG-49 and purified Cel5M (500 U/mL) (Figure 8(e)), or with strain Cel5M-HG-49 alone (Figure 8(f)). These results indicated that the gummy substances on the surface of fiber bundles were mostly removed during the bio-degumming process, and the degumming efficiency of recombinant strain Cel5M-HG-49 was significantly higher than that of wild-type strain HG-49. The results showed that glucomannanase Cel5M is an efficient enzyme for ramie degumming and would be a good candidate for application in the textile industry. The results also showed that the removal percentage of gum substances by the enzyme does not fully show the actual effect of the enzyme, which was seen better in the electron microscopic images of the degummed ramie fibers. Electron microscopy indicates that even a small decrease in percentage amounts on the surface gums is significant.
Scanning electron microscopy images of degummed ramie fibers (×300 magnification). With deionized water (a); refined ramie fibers (b); with Cel5M (500 U/mL) (c); with strain HG-49 (d); with strain HG-49 and Cel5M (400 U/mL) (e); with the engineered strain Cel5M-HG-49 (f).
Discussion
The conventional chemical processes for ramie degumming consume large quantities of chemicals and energy, and result in the release of large amounts of wastewater with high COD (chemical oxygen demand) value, causing serious risk of environmental pollution. 3 In addition, after chemical degumming, the fineness, glossiness and breaking strength of natural fibers are partially weakened. 24 Bio-degumming greatly reduces the consumption of NaOH and coal, along with a reduction in the COD value, with significant improvement in fiber properties. 3 Therefore, developing a cost-efficient and environmentally friendly degumming process represents a positive benefit to the textile industry, in which a well-functioning biological technology is a key challenge.
P. carotovorum HG-49 was shown to be effective in ramie degumming, with a gum removal ratio of 82.2%. However, some problems, such as low glycosyl hydrolase activity toward hemicellulose and non-crystalline cellulose, limited the complete removal of gum by P. carotovorum HG-49. In a recent study, it was shown that the total hemicellulose content (14.50%) was formed mainly of a mixture of galactoglucomannan (6.78%), glucomannan (3.45%) and xylan (1.25%) in RBFs. 22 The use of endoglucanase, particularly acting on less ordered cellulose, enlarged the spaces between the crystalline region and partially hydrolyzed amorphous region, decreased fiber stiffness, enhanced softness and brightness and then reduced or even eliminated the itch feeling of the ramie fabric. 25
In this study, we used in ramie degumming a novel glucomannanase enzyme Cel5M from P. carotovorum HG-49 that has also endoglucanase and xylanase activity. The Cel5M and its catalytic domain Cel5MD were heterologously expressed in E.coli BL21, and characterized in terms of its potential suitability for bio-degumming application. The Cel5M exhibited maximum activity at 60℃ with CMC and 65℃ with KGM as the substrate. The KGM substrate clearly stabilized the full-length enzyme. Both substrate and the hydrolysis products can stabilize the enzyme. 26 Although Cel5MD showed an optimum temperature at 55℃ with both substrates, the enzyme showed higher activity with KGM than with CMC in the temperature region of 65–80oC. These findings showed that the slightly branched glucomannan substrate stabilized the enzyme better than linear CMC. Therefore, the full-length enzyme is likely to function well at temperatures up to about 50–60oC in ramie degumming. The optimum temperature of both enzyme forms is much higher than another two similar reported glycoside hydrolases: endoglucanase CelA (T opt 45℃) from Erwinia carotovora subsp. Carotovora LY3427 and cellulase CelV (T opt 42℃) from Erwinia carotovora subsp. Carotovora SCRI193, 28 but similar to that of endo-cellulase CelN (T opt 57℃) from Erwinia carotovora subspecies atroseptica (Eca). 29
CBMs are commonly found in glycoside hydrolases and may improve the hydrolytic activity of the enzymes toward their substrate by increasing affinity and proximity to the substrate. 9 We found out that the presence of CBM3 in Cel5M resulted in an increase in the capacity to hydrolyze a soluble substrate mainly composed of β-1,4-glycosidic bonds. A similar effect was observed for CBM of the Bifidobacterium animalis GH5 enzyme, which was shown to increase the activity of GH5 toward mannans. 30 In addition, Cel5M showed higher thermostability at high temperature, chemical resistance and stability to metal ions than Cel5MD. Since the presence of CBM3 increase the tolerance to inhibiting metal ions, that is, it partly relieved the inhibitory effect of metal salts, it is likely that the higher binding of substrate with CBM3 was the reason for the effect. This indicates that the metal ions inhibit the enzyme at least partly by a competitive inhibition mechanism.
CBMs binding to the substrate directed glucomannanase activity to the removal of the major pectic polysaccharides (homogalacturonan) from the plant cell wall structures, which are densely packed with cellulose, pectin and hemicellulose.31,32 Since Cel5M is able to cleave various fiber molecules (mannans, CMC, xylan), it thus has activity to a wide substrate spectrum, which is apparently a very important property in removing gum structures. The inspection by electron microscopy revealed that the additional Cel5M activity together with HG-49, despite the low increase in the removal percentage of the gums, is essential in removing gummy structures from the fiber surfaces.
It appears that a strategy for efficient bio-degumming is to find the right multiactivity enzymes to achieve the highest removal of gum substances from the fiber surfaces. Microbial strains with a high secretion of pectinase have been regarded as efficient ramie-degumming strains, such as Amycolata autotrobutylicun, Bacillus amyloliquefaciens, Bacillus cereus P05, Bacillus licheniformis, Bacillus pumilus, Bacillus subtilis, Bacteroides finegoldii, Clostridium acetobutylicum, Pseudomonas brassicacearum, Pectobacterium wasabiae, Rhizobium leguminosarum and Streptomyces lividans. 33 However, along with increasingly stringent environmental requirements and the quality of degummed fiber and the textile market demand, there is a need to design novel industrial degumming strains with an efficient enzyme system.
Conclusions
In this study, we managed to enhance the glucomannanase and endoglucanase activity of a degumming strain by transferring a recombinant expression vector pUC18 with endoglucanase Cel5M into P. carotovorum HG-49. We showed that the glucomannanase activity and CMCase activity of the new strain Cel5M-HG-49 were clearly higher than that of the wild-type strain HG-49 during the cultivation in LB medium. The maximal glucomannanase activity (41.2 U/mL) was found to be higher than that of the wild-type HG-49 strain (28.3 U/mL) and the maximal CMCase activity (10.3 U/mL) was found to be higher than that of the wild-type HG-49 strain (0.1 U/mL), which indicated that the Cel5M gene was over-expressed in the engineered Cel5M-HG-49 strain. In addition, after pretreatment by recombinant strain Cel5M-HG-49, the surface of the ramie fiber was smoother and the scratchiness of the fabrics was decreased.
Therefore, the improving of the ramie-degumming efficiency and fiber quality via the overexpression of Cel5M in P. carotovorum is likely to reduce the consumption of chemicals and energy in ramie degumming. The over-expression of versatile Cel5M made the engineered Cel5M-HG-49 strain more promising in ramie degumming than the wild-type strain. Therefore, the further development of application processes based on in situ production of processing enzymes by the engineered microbial strains could offer alternative technologies to the textile industry.
Footnotes
Authors' note
Yawei Wang is also affiliated to College of Life Science, South-Central University for Nationalities, China.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was financially supported by the Natural Science Foundation of China (Grant No. 21676111) and the Major Technological Innovation of Hubei Province of China (Grant No. 2018ABA093).