
Abstract
Informed by recent developments in the field of evolutionary psychology, we elicit novel insights about the causal relationship between the activation of mating motives and gender-specific preferences for risk taking in tourism. In this endeavor, across four experimental studies, we illustrate that the effect of activating mating motives on the propensity for risk-taking tourism will be greater for men than it is for women. That is, when mating motives were salient, men consistently displayed a greater preference for the choice of high-risk travel activities (studies 1A and 1B), adventurous destination choices (study 2), and adventurous appeals in travel advertising (study 3). Our findings offer important theoretical contributions to research into the role of gender in risk-taking tourism behaviors from the perspectives both of evolution-based theorizing and managerial implications for tourism practice.
Introduction
It is time for tourism behavior and consumption researchers to look beyond the traditional model of Economic Man and to think instead in terms of Darwinian Man. If . . . genes keep culture on a leash, then an understanding of when, how, and why that leash influences tourist behavior has the potential to produce profound insights into the social science of tourism and the policy development that arises from it. (Crouch 2013, 587)
In the excerpt above, Crouch (2013) draws attention to the potential of evolutionary psychology to advance tourism research and practice, calling for further investigations in this area. In response to this call, our article uses the evolution-based theorizing approach and illustrates how the activation of fundamental mating motives (Griskevicius et al. 2007) sheds a novel perspective on the gendered differences in risk-taking tourism behaviors (Carr 2002; Lepp and Gibson 2008; Pizam et al. 2004).
Across diverse fields in the social sciences, evolutionary psychology has emerged as an important lens for the study of human behaviors (Saad and Gill 2000; Fennell 2006; Hill and Buss 2006; Griskevicius and Kenrick 2013; Crouch 2013). The key tenet of this view is that social cognitions and behaviors have an evolutionary “sociobiological” (E. O. Wilson 1975) underpinning—that is, all motivational systems are conceived to be developed to increase the survival and reproductive success of our ancestors. Therefore, rather than concerning itself only with the direct consequences of particular psychological systems per se (hereinafter referred to as “proximate” causes), an evolution-based theorizing approach aims to understand the underlying sociobiological explanations as to why these systems exist in the first place (hereinafter referred to as “ultimate” causes) (Saad and Gill 2000; Griskevicius and Kenrick 2013).
Within this rapidly growing body of research, mating acquisition is understood to be as one of the fundamental motives that addresses the sociobiological challenge of successfully attracting and reproducing with mates (Kenrick et al. 2010; Griskevicius and Kenrick 2013). Previous literature in this domain shows that mating motives can be primed temporarily or activated by external cues, such as by showing images of attractive opposite-sex individuals (Griskevicius and Kenrick 2013). Importantly, it has been found that when mating motivations are activated, they lead people to want to be noticed; however, the ways in which men and women draw attention to themselves are different (Buss 1989; Saad and Gill 2003; Griskevicius and Kenrick 2013). Simply put, activating mating motives can result in an increase in explicit gender-specific behaviors.
Against this backdrop, Crouch (2013, 580) postulates that the ultimate explanation of tourism behavior resides in its ability to signal an individual’s mastery of exploring new territories, which are “both useful attributes in determining sexual selection and status.” This proposition provides an important basis for advancing extant knowledge about risk-taking tourism behaviors (Lepp and Gibson 2008; Pizam et al. 2004; Reichel, Fuchs, and Uriely 2007), where it has been suggested previously that the difference in men’s and women’s propensity for risk taking could determine their preferred tourism activities and destination choices (Pizam et al. 2004; Reichel, Fuchs, and Uriely 2007). From an evolution-based perspective, men rather than women are likely to be more risk taking when their mating motives are salient (Baker and Maner 2008). Therefore, it is possible that activating mating motives can result in distinct and predictable gender-specific propensities for risk-taking tourism behaviors. The main purpose of our article is, therefore, to explore this proposition.
In this endeavor, across four experimental studies, we have established a causal relationship between activating mating acquisition motives (Kenrick et al. 2010; Griskevicius and Kenrick 2013) and the elicitation of gender-specific preferences for risk taking in tourism (Pizam et al. 2004; Reichel, Fuchs, and Uriely 2007). We found that, when mating motives are active, men consistently favor risk-taking travel activities, destination choices, and adventurous tourism advertising appeals more than women do. Our findings offer important theoretical contributions to research into the role of gender in risk-taking tourism behaviors from the perspective of evolution-based theorizing, as well as managerial implications for tourism practice.
Literature Review and Theoretical Justification
Evolution-Based Theorizing Approach
The application and advancement of the evolution-based theorizing approach to developing novel insights about social behaviors relies on two main principles—ultimate functionality and domain specificity (Saad 2007; Kenrick and Shiota 2008; Griskevicius et al. 2009). Ultimate functionality refers to the existence of fundamental ancestral challenges that human beings have needed to surmount, and to the psychological systems that have helped them to achieve this (Scott-Phillips, Dickins, and West 2011). Griskevicius and Kenrick (2013) have recently summarized the fundamental motives framework that details these major ancestral challenges and the associated motivational systems that functioned to resolve them. The fundamental motives include self-protection, disease avoidance, affiliation, status, mate acquisition, mate retention, and kin care (Griskevicius and Kenrick 2013). These motives provide a sociobiological theoretical basis to develop subsequent novel insights regarding the specific instances of behaviors across different domains (e.g., tourism). In other words, when adopting an evolution-based theorizing approach, researchers may start by asking the following question: “How does a particular behavior, cognition, emotion and/or perception constitute a functional solution to an adaptive problem in our evolutionary past?” (Saad and Gill 2000, 1006).
The second key principle is domain specificity (Griskevicius et al. 2009; Kenrick et al. 2010). This principle asserts that, even though there may be some perceived similarities between the expressions of different fundamental motives, they are qualitatively distinct. That is, each fundamental motive evokes a particular set of domain-specific cognitive and behavioral responses. As Griskevicius et al. (2009, 386) explains, this is because “mental mechanisms well-suited to solving one adaptive problem are often ill-suited to solving another.” In other words, different adaptive problems require distinct, and often incompatible, solutions—and are solved more effectively by independent psychological systems (Kenrick et al. 2010).
Thus, the evolution-based theorizing approach draws on sociobiology to identify fundamental motives that serve distinct functions within the meta-purpose of increasing reproductive success and survival. When activated, each motive is expected to evoke a distinct set of consequences for cognition and behaviors in relation to the specific kinds of survival-relevant stimuli (Kenrick et al. 2010). Accordingly, compared to proximate explanations of social behaviors that tend to characterize motives according to willingness as opposed to evasion (Griskevicius et al. 2009), the ultimate evolution-based approach conveys a more holistic perspective—that is, it draws attention to the underlying survival and reproductive motives that guide social cognitions and behaviors (Kenrick et al. 2010). Thus, by establishing what particular type of fundamental motives orchestrate a given behavior, this approach fosters a more complete understanding and thus improves the predictability of this behavior.
In tourism studies, the application of sociobiology and evolution-based theorizing was pioneered by Fennell (2006), who illustrated how a sociobiological theory of reciprocal altruism (Trivers 1971) offers an important perspective to evaluate the nature of short-term and long-term collaborative interactions in tourism, advocating for a need to foster ethics in securing long-term symbiosis in tourism space. Following Fennell (2006), several other studies have endeavored to apply the tenets of reciprocal altruism in developing insights regarding travel behaviors across different settings (Plummer and Fennell 2007; Paraskevaidis and Andriotis 2017). However, with the exception of this emerging stream of research, other evolutionary motives and their relevance to the field of tourism have received limited attention (Crouch 2013).
Therefore, the purpose of this study is to use the principles of evolution-based theorizing to discuss the relevance of one particular fundamental motive—acquisition of a mate—to advance our knowledge regarding the gendered differences in risk-taking tourism behaviors.
Mate Acquisition Motives
Acquisition of a mate is one of the fundamental motives that underpins humans’ needs to attract partners and reproduce (Griskevicius et al. 2009; Griskevicius and Kenrick 2013). Previous studies have shown that this motivational system can be activated by a range of external cues that elicit sexual desires, such as photos or videos that depict attractive people of the opposite sex (Griskevicius et al. 2009). The evolution-based perspective postulates that, when activated, the urge to acquire a mate promotes people to adopt strategies that are likely to maximize their mating success, such as engaging in “salient positive differentiation”—attracting the attention of the opposite sex and differentiating themselves from the same-sex rivals (Griskevicius et al. 2007).
While both men and women engage in salient positive differentiation, they tend to adopt different gender-specific strategies for choosing their mating partners and to maximize their mating success. For instance, some studies report that, when selecting long-term mating partners, men tend to prioritize physical attractiveness, whereas women may put more emphasis on status and resources (Li and Kenrick 2006; Miller 2009). Li and Kenrick (2006) attribute such differences to particular sets of sociobiological functions, whereby women adaptively allocate more effort toward ensuring resources for their offspring, and men toward ensuring sufficient fertility (Griskevicius et al. 2006; Grammer et al. 2003). Informed by this perspective, a number of studies have explored how mating motives influence behavior in various areas, such as conspicuous consumption (Griskevicius et al. 2007; Nelissen and Meijers 2011), creative activities and arts (Griskevicius et al. 2006), prosocial behaviors (Iredale, Van Vugt, and Dunbar 2008), loss aversion and smoking (Li et al. 2012; Chiou, Wu, and Cheng 2015), and preference for sexualized advertisements (Kim and Kim 2016; Sengupta and Dahl 2008).
Importantly, there is also evidence to suggest that mating motives can foster more risk-taking propensity among men than among women (Baker and Maner 2009; Knutson et al. 2008). For instance, Griskevicius et al. (2006) have shown that mating motives can influence men to display nonconforming behavior, which conveys their ability and willingness to take risks. Such motives can also induce impatience and impulsiveness among men, which are the focal traits associated with risk-taking propensities (Carducci and Wong 1998). In one experiment, after seeing attractive women’s faces, men chose smaller instantaneous rewards rather than delayed larger rewards (Wilson and Daly 2004). Similarly, other studies have found that mating motives selectively erase loss aversion—and therefore risk avoidance—among men, but not among women (Kenrick et al. 2009; Li et al. 2012).
Previous literature, therefore, suggests that mate acquisition motives can significantly alter people’s behaviors, resulting in gender-specific strategies with regards to risk taking. Baker and Maner (2009) explain that these gendered effects occur for two reasons. First, risk taking conveys mating-favorable characteristics, such as social dominance, confidence and ambition, which are desired by women rather than men when seeking a romantic partner (Buss 1989; Li et al. 2002). Accordingly, when men engage in salient positive differentiation, they can gain more strategic benefits by conspicuously displaying such mating-favorable, risk-taking behaviors (Zahavi 1975). Second, M. Wilson and Daly (1985) note that men have adaptively experienced greater competition from the same-sex rivals than women over access to mating opportunities, which puts more pressure on them to engage in relatively high risk–high payoff strategies when seeking mates. Thus, these studies provide evidence that activating mating motives can produce more pronounced gendered differences by making men more risk taking than women.
It is important to note, however, that the greater propensity for risk taking in men is not a fixed propensity and varies greatly according to different situational factors (Daly and Wilson 2001). For instance, men could be more or less inclined to risk taking according to their emotional state or social contexts—regardless of whether or not their mating motives are activated (Baker and Maner 2009). Accordingly, there could be important boundary conditions and specific contexts where the gendered effects associated with mating acquisition are particularly salient. In the following section, we argue that the ultimate purposes underpinning tourism activities (Crouch 2013) allow mating motives to become particularly important in the context of risk-taking tourism behaviors.
Risk-Taking Tourism Behavior
Risk-taking tourism behavior has been a focus of many studies in the tourism literature (Lepp and Gibson 2008; Uriely and Belhassen 2006; Pizam et al. 2004; Berdychevsky and Gibson 2015). Many of these studies have considered whether risk-taking tourism behaviors could be associated with certain personality traits (Plog 1973; Pizam et al. 2004; Lepp and Gibson 2008). Pizam et al. (2004) conducted a cross-cultural analysis across 11 different countries to explore how the combined psychological characteristics of risk-taking and sensation seeking (RSS) influence tourism-related decision-making and behaviors. Their findings show that scoring high on RSS characteristics is associated positively with engaging in adventurous tourism (e.g., Fuchs 2013; Plog 1973; Pizam, Reichel, and Uriely 2001; Walle 1997). Moreover, Pizam et al. (2004) found that there was a greater proportion of high-RSS respondents among men than among women. In another study, Reichel Fuchs, and Uriely (2007) have also found that men are less risk-averse compared to women. In a similar vein, Lepp and Gibson (2008) note that sensation seeking—a personality trait associated with a need for novelty and simulation—is correlated positively with traveling to regions of the world that are rated as riskier. However, while these authors concur with Pizam et al. (2004) that men tend to score higher on sensation seeking than do women, Lepp and Gibson (2008) found no evidence that gender has a direct impact on the actual risk-taking tourism behaviors. These results are consistent with Carr (2002), who notes that there may be very few gendered differences in terms of the actual leisure activities of tourists.
Extant theory regarding risk-taking tourism behaviors, therefore, offers mixed answers to questions about how and when gendered differences exert significant influence on travel choices and behaviors (Reichel, Fuchs, and Uriely 2007; Fuchs 2013). Although scholars tend to agree that men and women score differently in terms of the personality traits, such as sensation seeking, that are associated with adventurous tourism activities and risk taking, there is conflicting evidence in respect of the manifestations of such effects in the tourists’ actual behaviors and choices. The possibility that there could be other important factors that moderate this relationship has been mentioned (e.g., Carr 2002) but underanalyzed. Given that risk-taking and adventurous experiences are important aspects of traveling—and that the associated decision-making processes could be different for men and women—it is important that our theorizing can establish causally whether there are gendered differences in risk-taking tourism, and if so, when they occur, and how to account for such differences theoretically.
To inform our understanding of these important issues, we propose to adopt an evolution-based theorizing approach, and conduct our research on mate acquisition motives. In this endeavor, we draw on Crouch’s (2013) view that human propensity to travel could be underpinned by the fundamental purposes that have evolved from sociobiological origins. One of these purposes has been discussed by Fennell (2006), who draws attention to the social nature of reciprocal altruism (Trivers 1971), and discusses its relevance for building trust and cooperation in tourism through repeated interactions. That is, traveling can be considered as a means to expand social networks that offer greater opportunities for collaboration, which is necessary to produce gains in chances for collective survival and reproduction (Trivers 1971). Another purpose of traveling could also be signaling an individual’s mastery in exploring new territories, which is relevant to the salient positive differentiation used to succeed in acquisition of a mate (Griskevicius et al. 2007). This suggests that tourism could be an important area of human activity where evolutionarily fundamental motives, more broadly, and acquisition of a mate, in particular, can exert a particular importance in decision-making and behaviors.
Against this backdrop we have established that when mate acquisition motives are active, the propensity for risk taking tends to be more salient for men’s (but not women’s) salient positive differentiation (Baker and Maner 2009). In the context of travel research, this suggests that by activating mate acquisition motives, we could expect to observe more pronounced and predictable gendered differences with regard to risk-taking propensities. Specifically, when mating motives are salient, men can gain more strategic benefits for accomplishing their mate acquisition goals by participating in risk-taking tourism activities. This is because such activities convey social dominance, confidence, and ambition, and these are adaptively valued more by men in their attempts to gain status and reputation (Baker and Maner 2009). Accordingly, men are more likely to engage in risk-taking tourism behaviors despite the associated costs (Zahavi 1975). On the other hand, for women, even if mating motives are salient, risk-taking activities provide very few strategic benefits (Buss 1989; Li et al. 2002). Thus, women have less incentive to engage in costly behaviors involving risks, because such behaviors are less strategically useful for the purposes of attracting desirable mates (Goldberg 1995). Taken together, these propositions inform our main prediction with regard to the role of gender in risk-taking tourism, which can be summarized as follows:
General prediction: Mate acquisition motives elicit pronounced gendered differences with regard to risk-taking tourism behaviors, such that the effect of activating mating motives on the propensity for risk-taking tourism will be greater for men than for women.
The main purpose of our study is to test the general prediction outlined above. In this endeavor, we conducted four experimental studies. In study 1A, we show that activating mate-seeking motives has a greater impact among men than among women on the choice of high-risk activities associated with adventure tourism. In study 1B, we also show that the impact of mating motives is different from the responses to a romantic scenario concept activation. In study 2, we extend our results by showing consistent effects of mating acquisition on the gendered differences in the context of choosing between adventurous and nonadventurous travel destinations. We also establish how activating mate-seeking motives differs from the activation of another fundamental motive—kin care (Glocker et al. 2009). In study 3, we establish how mating acquisition motives produce gender-specific evaluations of and purchase intentions toward advertising appeals for adventurous travel.
All participants for our studies were recruited from Amazon Mechanical Turk online panel (Casler, Bickel, and Hackett 2013; Buhrmester, Kwang, and Gosling 2011), which has been used extensively in previous tourism studies (e.g., Choi et al. 2017; Kim, Kim, and Kim 2018). We detail our participants’ demographic information in Table 1. Furthermore, consistent with other studies that explored mating acquisition motives, we limited our participants to heterosexual adults aged between 18 and 40 years (Griskevicius et al. 2007; Kim and Kim 2016). In order to determine an adequate sample size, we conducted the power analysis using G*Power 3 software (Faul et al. 2007). Based on our experimental settings (i.e., power [1 – β = .80] with medium effect size [f = .30] at the significance level α = .05, 2-factorial design), the minimum required sample size was 90. Therefore, we recruited more than 90 participants for each study that activated mating motives.
Profile of Participants.
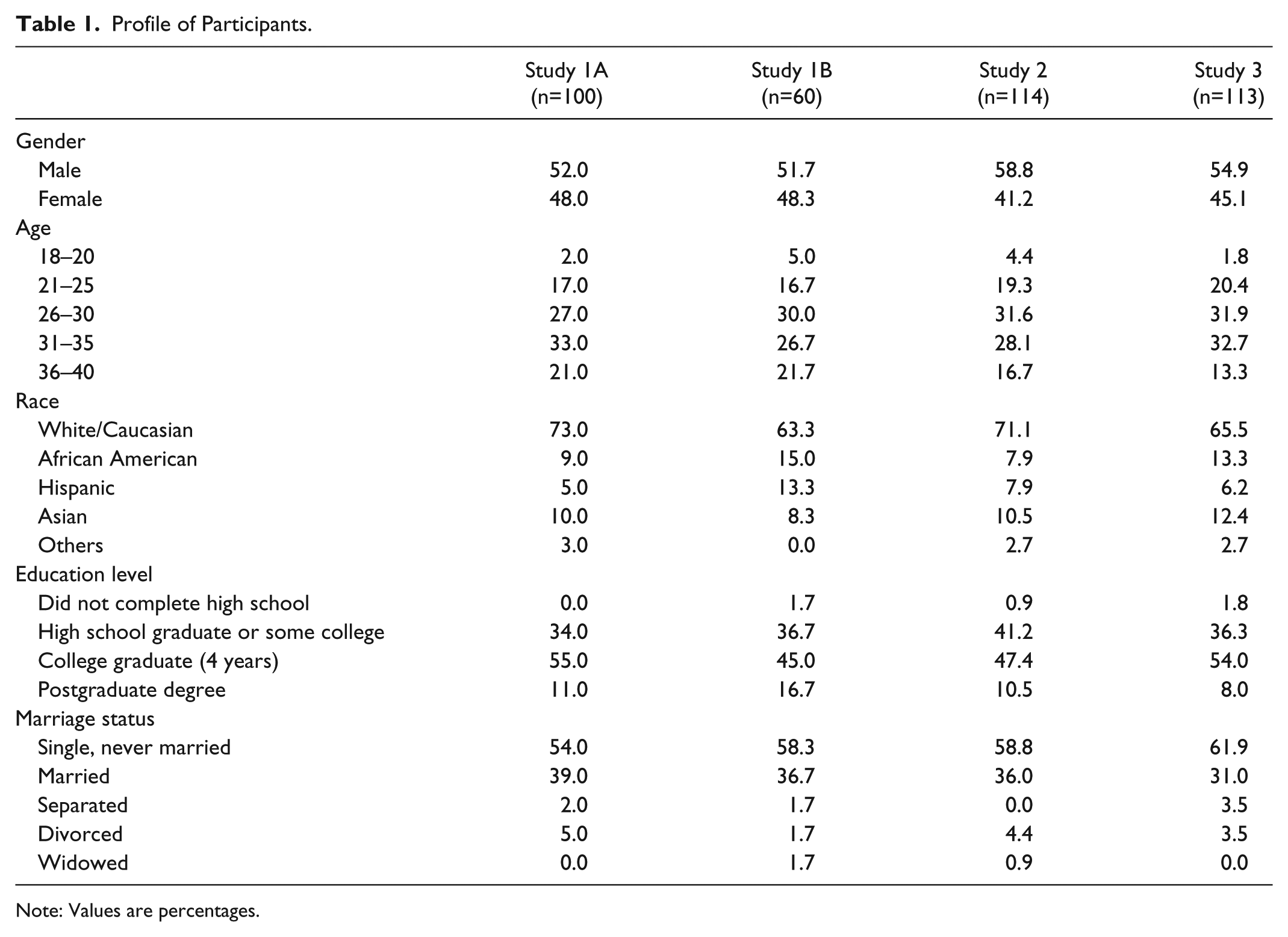
Note: Values are percentages.
Study 1A
Travel marketers and tourism retailers require a deeper understanding of how people evaluate and choose tourism activities when they travel (Moscardo et al. 1996; Sung, Morrison, and O’leary 2000; Swanson and Horridge 2004). Against this backdrop, we begin to test our general prediction regarding the role of mate-seeking motives in eliciting gendered differences in risk-taking tourism choices, by illustrating empirically how the activation of mating motives produces greater preference for high-risk activities associated with adventure tourism among men than among women.
Sample and Procedure
A total of 100 (48 women, Mage = 30.78, age range = 20–40) participants were randomly assigned to a 2 (mating motives: activation vs. control) × 2 (gender: men vs. women) between-subjects experimental design.
At the beginning of the experiment, participants were briefed that they would be completing multiple tasks. First, they were asked to report their gender. Based on this information, we manipulated mating acquisition motives (active vs. control), consistent with the techniques that were used in previous evolution-based studies (Griskevicius, Cialdini, and Kenrick 2006; Griskevicius et al. 2007; Kim and Kim 2016). In a control condition, participants were shown images of three different streets and were asked to choose the street that they perceived to be the most desirable to walk on. In a mating activation condition, participants were asked to imagine that they did not currently have a romantic partner. They were then exposed to the images of three attractive individuals of the opposite sex and were asked to choose one whom they would consider to be the most desirable as a potential partner. Next, participants were asked to evaluate the overall attractiveness of their chosen option (i.e., the attractiveness of their chosen potential partner in the activation condition, or of their chosen street in the control condition) on a 7-point scale (1 = not at all attractive, 7 = definitely attractive). Finally, they completed a short writing task on the activities that they imagined participating in (within their given scenario).
Following the mating activation exercise, participants were asked to imagine that they were planning a trip to New Zealand. They were shown 10 different activities and were asked to choose 4 that they would like to do during the trip. Four of these activities (bungee jumping, skydiving, rafting, and rock climbing) involved relatively higher risk than the other 6 (winery tour, harbor cruise, fishing, visiting galleries, dolphin-watching, and garden tour). Accordingly, each participant’s choice could include up to four high-risk activities, which we used to derive a continuous dependent variable (range: 0–4) for our main analysis. Following this task, participants evaluated all 10 activities on a 6-point scale (1 = not at all risky, 6 = extremely risky) (Park and Reisinger 2012), and their mood on a 7-point scale (1 = very bad, 7 = very good). Finally, participants were thanked and debriefed.
Results
Preliminary analysis revealed that the manipulation of high-risk activities was successful. The four activities that we chose to represent high-risk options associated with adventure tourism (M = 4.81, SD = 0.98) yielded a significantly higher perception of risk than the remaining six activities (M = 1.83, SD = 0.92); t(99) = 18.80, p < .001. Moreover, we found that rafting—which yielded the lowest risk perception among the four high-risk options—conveyed a significantly higher perception of risk than fishing—which yielded the highest-risk perception among the remaining low-risk options (Mrafting = 4.24, SD = 1.27; Mfishing = 2.28, SD = 1.30); t(99) = 11.65, p < .001.
For manipulation checks of the scenario-based mating acquisition motives, we conducted a 2 (mating motives: activation vs. control) × 2 (gender: men vs. women) analysis of variance (ANOVA) on the attractiveness of the chosen target (i.e., a partner or a street), consistent with other studies that employed similar priming techniques (Griskevicius et al. 2007; Kim and Kim 2016). Only the main effect of mating motives activation was found significant, F(1, 96) = 11.26, p < .01. The evaluation of potential partners in an activation condition (M= 6.52, SD = 0.62) yielded higher perceived attractiveness than the evaluation of streets in a control condition (M = 6.06, SD = 0.62). However, the analysis that included the target’s attractiveness—as a covariate—did not significantly alter our main findings. In addition, the main and interaction effects of mating motives activation and gender were not significant for mood evaluation (all p’s > .32). Importantly, the attractiveness of the chosen partner target between male and female participants yielded no significant difference, F(1, 46) = 0.05, p = .82; Mmen = 5.64, SD = 0.59 vs. Mwomen = 6.50, SD = 0.66. Thus, our manipulation of mating acquisition did not seem to produce external bias and we proceeded to investigate the main findings of our study.
A 2 (mating motives: activation vs. control) × 2 (gender: men vs. women) ANOVA on the preference for high-risk activities yielded a significant main effect of gender; F(1, 96) = 5.81, p < .05. Men displayed more preference for high-risk tourism activities (M = 1.08, SD = 1.08) than did women (M = 0.65, SD = 0.81). More importantly, however, we found a significant two-way interaction; F(1, 96) = 4.00, p < .05. Planned comparisons revealed that men selected more high-risk activities when mating motives were activated (M = 1.42, SD = 1.25) compared to the control condition (M = 0.79, SD = 0.83); F(1, 96) = 5.79, p < .05. Conversely, women showed consistent low interest in high-risk activities across both the mating activation (M = 0.58, SD = 0.88) and control conditions (M = 0.71, SD = 0.75); F(1, 96) = 0.21, p > .10. These results are depicted in Figure 1.
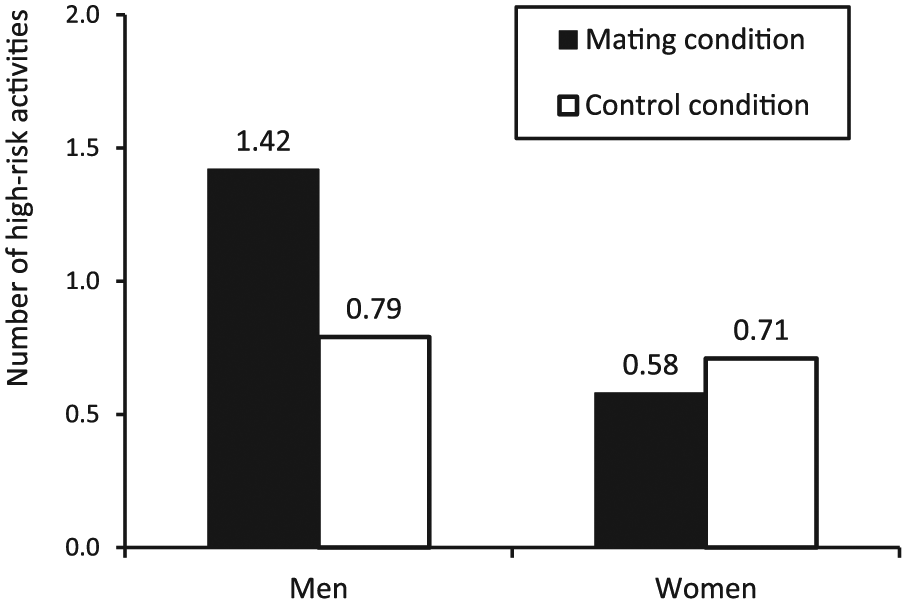
Results of study 1.
In addition, we also found that the relative preference for high-risk activities among men under the mating activation (vs. control) condition was consistent across all four high-risk activities as follows: bungee jumping (Mmating = 29.2% vs. Mcontrol = 10.7%), skydiving (Mmating = 33.3% vs. Mcontrol = 14.3%), rafting (Mmating = 45.8% vs. Mcontrol = 28.6%), and rock climbing (Mmating = 33.3% vs. Mcontrol = 25.0%). This provides evidence that the results of study 1A were not driven by any one particular high-risk activity associated with adventure tourism. Further, we also included marital status as an additional factor, but this has not changed our results.
Summary
The results of study 1A provide initial support for our main prediction that mating motives can elicit pronounced and predictable gendered differences with regards to risk-taking tourism. Consistent with several previous studies (e.g., Meng and Uysal 2008), we found that men, in general, preferred high-risk activities associated with adventure tourism more than women did. More importantly, however, we show how the activation of mating motives makes this effect even more explicit, having a greater effect on men than on women. That is, when mating motives were activated, the preference for high-risk activities among men increased significantly, whereas it had no effect on women.
Study 1B
One may question whether the findings of study 1A result merely from people thinking that they are embarking on a romantic trip (concept activation), rather than because they are motivated to engage in salient positive differentiation because their mate-seeking motives have been activated (goal activation). Put differently, in our previous study, it is possible that participants were assimilated to the romantic concept rather than primed to activate the mating-seeking motives. Since the concept and goal activations are theoretically different (see Förster, Liberman, and Friedman 2007 for details), we test this possibility in the follow-up study. Specifically, in order to exclude the alternative explanation, we conducted a study that manipulates the romantic concept activation—the scenario was of traveling with a romantic partner.
Sample and Procedure
A total of 60 (29 women, Mage = 30.07, age range = 19–40) participants were assigned randomly to a 2 (romantic concept: activation vs. control) × 2 (gender: men vs. women) between-subjects experimental design.
At the beginning of this experiment, participants were asked to imagine that they were planning a trip to New Zealand. In a control condition, they were also asked to imagine that they were single and would be traveling by themselves. In the romantic concept activation, they were asked to imagine that they had a romantic partner and would be embarking on a trip together. Next, they were asked to choose 4 of 10 activities that they would like to take part in during the trip, using the same procedures as in study 1A. Following this task, participants evaluated whether they perceived the trip to be romantic using a 7-point scale (1 = not at all romantic, 7 = very romantic). Finally, participants were thanked and debriefed.
Results
A 2 (romantic concept: activation vs. control) × 2 (gender: men vs. women) ANOVA yielded a marginally significant main effect of the romantic concept activation, F(1, 56) = 3.56, p = .064, whereby participants chose less high-risk activities when they imagined traveling with a romantic partner in the activation condition (M = 0.65, SD = 0.69), compared to when they imagined traveling by themselves in the control condition (M = 0.96, SD = 1.00). The main effect of gender was not significant, although the mean score for choosing high-risk activities among men (M = 0.84, SD = 0.86) was slightly higher than that for women (M = 0.72, SD = .84). Importantly, the interaction effect between gender and the romantic concept activation was not significant, F(1, 56) = 0.23, p = .63. This means that contrary to the results obtained from the mating motives activation in study 1A, the romantic concept activation in study 1B yielded a nonsignificant difference in men’s and women’s preference for high-risk activities.
Summary
In study 1B, we rule out an alternative explanation that the results of study 1A can be elicited merely by travelers’ considering a trip to be romantic in nature (i.e., concept activation). In fact, we observed that activating the romantic concept may produce opposite effects, such that tourists with partners could be more risk-averse than those without.
Study 2
In study 2, we extend the results of studies 1A and 1B in two ways. First, we explore the influence of mating motives on behavioral intentions when choosing a travel destination (Lam and Hsu 2006; Um and Crompton 1990; Woodside and Lysonski 1989). Traveling to riskier destinations may provide travelers with an additional reward of social acceptance and prestige (Lepp and Gibson 2008). This is in line with Bourdieu’s (1984) theory of capital, which postulates that people acquire economic, social, and cultural capital within their competition for positions of distinction and social status. That is, traveling to a high-risk destination could be perceived as a way to accumulate social capital. Therefore, we would expect that the activation of mating motives should elicit greater preference for traveling to riskier destinations among men than among women.
Second, this study juxtaposes the effect of mating acquisition motives against another type of fundamental motive—the kin care system (Griskevicius and Kenrick 2013). Kin care is defined as “the full-time care, nurturing and protection of children by relatives, members of their tribes or clans, godparents, stepparents, or other adults who have a kinship bond with a child” (Bell and Garner 1996, 11). This fundamental motive is evident within Hamilton’s (1964) inclusive fitness theory of altruism, which posits that natural selection would favor behaviors that benefit either the individuals, themselves, or closely related kin. Thus, caring for a child could be seen as adaptive given the genetic relatedness between a parent and a child. Contrary to the influence of mating acquisition, kin care motives do not encourage men to partake in risk-taking behaviors, because such costly behaviors offer little strategic benefits for protecting and supporting others (i.e., for satisfying kin care motives). Accordingly, we could expect that men would display more desire to visit riskier destinations when their motives serve a mate acquisition function rather than a kin care function.
Sample and Procedure
A total of 114 (47 women; Mage = 29.5, age range = 18–39) participants were randomly assigned to a 2 (fundamental motive: mating vs. kin care) × 2 (gender: men vs. women) between-subjects experimental design.
At the beginning of the experiment, participants were briefed that they would be completing multiple tasks. In the mating motives scenario, participants were primed using the same technique as in study 1A. In the kin care motives scenario, participants followed similar instructions. However, rather than being exposed to the images of attractive individuals of the opposite sex, they were shown images of three babies, and were asked to choose one whom they perceived to be the most desirable to take care of. Next, participants were asked to evaluate the overall attractiveness of their chosen option (i.e., the attractiveness of their potential partner in the mating motives condition, or of their chosen baby to take care of in the kin care motives condition) on a 7-point scale (1 = not at all attractive, 7 = definitely attractive). Finally, they completed a short writing task consistent with other studies.
Following the fundamental motives activation exercise, in order to conceal the true purpose of our experiment, participants were asked to complete five different choice tasks involving consumption decisions. Three of these tasks were unrelated for the purposes of our study, while the remaining two included the choice between travel destinations that varied in terms of their relative perception of risk (Keinan and Kivetz 2011). Participants were asked to imagine that they were planning a one-week trip, by themselves, and they had a choice between two destinations. In the first task, they had to choose between (a) Maui, Hawaii: “Relax at a luxurious resort. Enjoy perfect weather and beautiful beaches” [less adventurous destination] and (b) Costa Rica: “Explore the authentic culture, rainforests, volcanoes, and wildlife adventures” [more adventurous destination]. Participants were asked to indicate their relative preference on a 7-point scale (1 = I will definitely choose Hawaii; 7 = I will definitely choose Costa Rica). The second task involved a choice between: (a) Fort Lauderdale hotel, Florida [less adventurous destination] and (b) Ice hotel in Quebec [more adventurous destination] using the same 7-point scale. Since responses for the two tasks were significantly correlated (p < .05), we used the mean score of these two measures in our main analysis.
Following these choice tasks, participants were asked to evaluate the perceived adventurous appeal for all four destinations on a 5-point scale (1 = not at all adventurous, 5 = extremely adventurous) and indicate their mood on a 7-point scale (1 = very bad, 7 = very good). Finally, participants were thanked and debriefed.
Results
Preliminary analysis revealed that the manipulation of adventurous destinations was successful. Specifically, the two destinations that we chose to represent riskier destinations—Costa Rica and Ice hotel in Quebec (M = 4.24, SD = .75)—were perceived as being significantly more adventurous than the other two destinations—Hawaii and Fort Lauderdale hotel in Florida (M = 2.61, SD = 1.10); t (113) = 12.25, p < .001.
For manipulation checks of the scenario-based fundamental motives activation, we conducted a 2 (fundamental motives: mating vs. kin care) × 2 (gender: men vs. women) ANOVA on the attractiveness of scenarios’ targets. The analysis yielded a significant two-way interaction effect; F(1, 105) = 7.37, p < .01. The evaluation of mating targets’ attractiveness was not different for women (M= 6.26, SD = .92) and men (M = 6.31, SD = .71); F(1, 105) = .03, p > .10. However, the evaluation of kin care targets’ attractiveness was higher for women (M = 6.32, SD = .99) compared to men (M = 5.23, SD = 1.42); F(1, 105) = 13.86, p < .001. Our results are consistent with previous evolution-based studies, which suggest that women tend to display stronger kin care motivations than men (Griskevicius and Kenrick 2013). For the purposes of our main analysis, however, when we included the target’s attractiveness, as a covariate, it did not significantly alter our results. In addition, the main and interaction effects of fundamental motives activation and gender were not significant for mood evaluation (all p’s > .20). Thus, our manipulation of fundamental motives did not seem to produce external bias and we proceeded to conduct the main analysis of our study.
A 2 (fundamental motives: mating vs. kin care) × 2 (gender: men vs. women) ANOVA on the preference for riskier destinations did not yield any significant main effects. More importantly, however, the two-way interaction between fundamental motives and gender was significant, F(1, 109) = 4.44, p < .05. Planned comparisons revealed that men displayed more preference for risker adventurous destinations when mating motives were activated (M = 4.38, SD = 1.96) compared to when kin care motives were activated (M = 3.39, SD = 1.60); F(1, 105) = 4.46, p < .05. Conversely, women showed consistent interest in adventurous destinations across both the mating (M = 3.50, SD = 1.78) and kin care activation conditions (M = 4.05, SD = 2.23); F(1, 105) = .95, p > .10. These results are depicted in Figure 2.
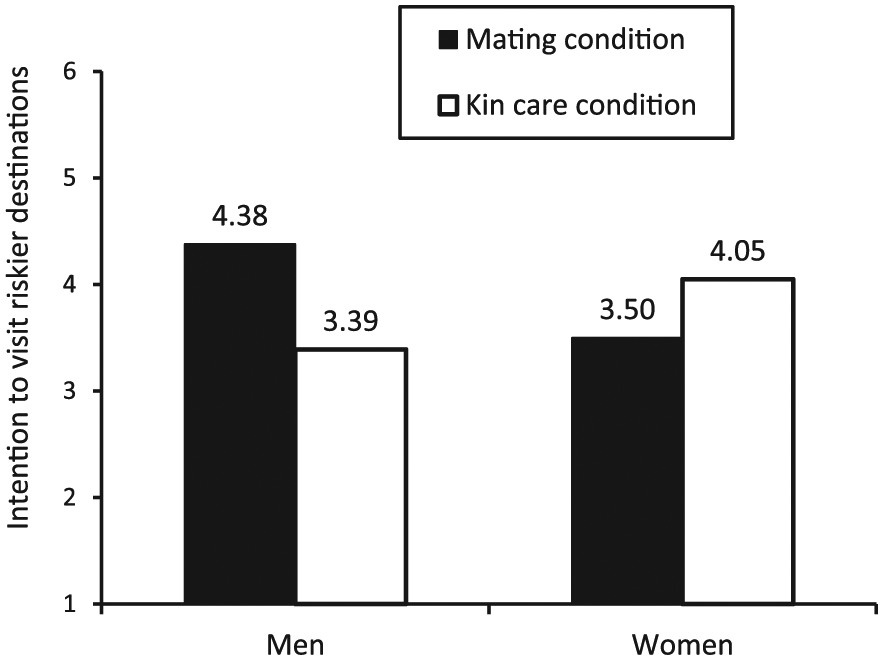
Results of study 2.
Summary
This study corroborates the results of studies 1A and 1B by extending our general prediction onto the behavioral intentions of choosing a travel destination, and by eliciting the unique effect of mating motives compared with the effect of another fundamental motive—the kin care system.
Study 3
In this final study, we extend our findings by considering the role of mating motives in predicting the effectiveness of travel advertising (Messmer and Johnson 1993). Here we show how the activation of mating motives results in divergent gender-specific purchase intentions in response to travel advertising appeals. In this endeavor, we developed two advertising messages for the same travel product, which maintained general consistency and varied only in terms of their relative perception of adventurousness (Lepp and Gibson 2008; Mihalič 2000). Thus, study 3 explores the effect of mating motives on the travelers’ behavioral intentions in response to the same travel product. Accordingly, this study addresses the limitation of our previous studies that employed choice-based tasks.
Sample and Procedure
A total of 113 (51 women, Mage = 29.7, age range = 18–39) participants were randomly assigned to a 2 (mating motives: activation vs. control) × 2 (advertising appeal: adventurous vs. nonadventurous) × 2 (gender: men vs. women) between-subjects experimental design.
First, we manipulated mating motives using the same technique as in study 1A. Participations in the control condition chose a street that they perceived to be the most desirable to walk on, whereas participants in the activation condition chose an image of a person that they would consider to be the most desirable as a romantic partner. Next, participants were asked to evaluate the overall attractiveness of their chosen option and complete a short writing task on the activities that they imagined participating in, within their given scenario.
Following the mating activation exercise, as a cover story for the experiment, participants were told that a travel agency “Great Mountain Tours” (a fictitious name) was planning to run print advertisements for travel packages that were priced at $1,300, and that the participants were going to see a mock-up advertisement being prepared to give an impression of how the final print would look. Each participant was shown only one of the two mock ads. The ads maintained visual consistency, but included one of two blocks of the ad copy that discussed the adventurousness of the travel package. The adventurous appeal ad copy said, “Capture an extreme adventure at a whole new altitude with Great Mountain Tours! You live only once, but you can feel alive every moment . . .” The nonadventurous appeal ad copy said, “Experience tranquility at a whole new altitude with Great Mountain Tours! Be at one with Nature and be at one with yourself. . .” On the following screen, participants were asked to indicate their attitude toward the tour package on three 7-point scale items (1 = very bad / very unfavorable / very negative, 7 = very good / very favorable / very positive, Cronbach’s α = .95), and their purchase intention on a 7-point scale (1 = very unlikely to purchase this package, 7 = very likely to purchase this package). Next, they were asked to evaluate their mood on a 7-point scale (1 = very bad, 7 = very good). Finally, participants were thanked and debriefed.
Results
In order to confirm the manipulation of adventurous advertising appeals, we conducted a separate pretest with 27 participants (15 women, average age = 31.3). Each participant was randomly assigned to view only one ad concept, and evaluated the risk appeal of an advertising message on two 7-point scale items (1 = conveys very low risk / not at all adventurous, 7 = conveys very high risk / very adventurous, Cronbach’s α = .71). The analysis revealed that the manipulation of adventurous appeals was successful. Participants rated the adventurous appeal ad copy (M = 5.22, SD = 1.20) to convey significantly higher risk than the nonadventurous appeal ad copy (M = 4.81, SD = 1.53; t(26) = 2.58, p < .05).
A 2 (mating motives: activation vs. control) × 2 (advertising appeal: adventurous vs. nonadventurous) × 2 (gender: men vs. women) three-way ANOVA on attitude toward the package did not yield any significant main or interaction effects (all p’s > .10). We, therefore, proceeded to conduct a three-way analyis of covariance on purchase intention that included attitude as a covariate. The results yielded a significant three-way interaction effect; F(1, 104) = 5.51, p < .05. To further investigate the underlying mechanism, we analyzed purchase intentions for each gender by conducting two separate 2 (mating motives: activation vs. control) × 2 (advertising appeal: adventurous vs. nonadventurous) ANOVAs—one with male participants and another one with women. The results showed no significant main or interaction effects between mating motives and advertising appeal among women (all p’s > .10). That is, women displayed insignificantly different purchase intention for adventurous and nonadventurous ads across both mating activation and control conditions; Mmating & adventurous = 4.10, SD = 2.33, Mmating & nonadventurous = 4.38, SD = 2.10, F(1, 47) = .65, p > .10; Mcontrol & adventurous = 3.40, SD = 2.23, Mcontrol & nonadventurous = 3.43, SD = 2.41, F(1, 47) = 2.08, p > .10.
For men, however, we found a marginally significant two-way interaction, F(1, 56) = 3.68, p = .06. Planned comparisons revealed that when mating motives were activated, men conveyed significantly higher purchase intention for the adventurous ad (Mmating & adventurous = 3.88, SD = 1.93) compared to the nonadventurous ad (Mmating & nonadventurous = 2.87, SD =2.13); F (1, 56) = 8.05, p < .01. On the other hand, in the control condition, there was no significant difference (Mcontrol & adventurous = 3.57, SD = 2.83, Mcontrol & nonadventurous = 3.94, SD =2.32); F(1, 56) = .01, p > .10. Thus, our three-way and two-way analyses offer additional support for our general prediction that mating motives elicit higher risk-taking propensities in tourism behaviors among men compared to women. We illustrate these results in Figure 3.
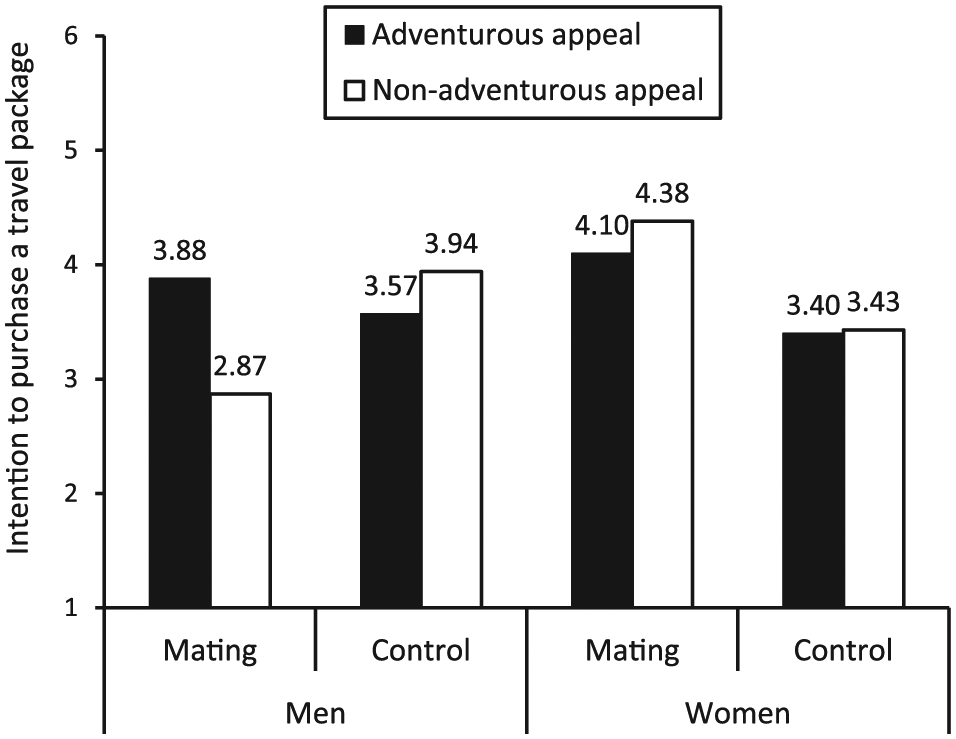
Results of study 3.
Summary
Within the set of four studies that we present in this article, study 3 had two main purposes. First, we corroborated the results of our previous studies in the context where travelers evaluate the same travel activity rather than choose between activities (studies 1A, 1B, and 2). Second, we extend the general prediction regarding the role of mating motives activation into the domain of travel advertising effectiveness (Messmer and Johnson 1993).
In addition, study 3 shows that, while mating motives had a significant impact on the gendered differences in behavioral purchase intentions associated with adventurous appeals, they did not produce an equivalent effect in terms of more general attitudes toward risk-taking travel. These findings resonate with calls to explore whether “males and females perceive and judge risks and consequences differently or . . . they instead respond differently to the same judgments (Crouch 2013). At least in the context of our study, we provide support for the latter argument—that is, men and women seem to exhibit different behavioral responses to the same judgment of risk associated with adventurous tourism.
General Discussion
Risk-taking and adventurous behaviors are important aspects of travel research and practice (Carr 2002; Lepp and Gibson 2008; Uriely and Belhassen 2006). However, there have been inconclusive results regarding when and how women and men differ in terms of their relative risk-taking travel preferences (Lepp and Gibson 2008; Fuchs 2013). Our study sheds a novel perspective on this gap in the literature by adopting an evolution-based theorizing approach to traveling (Fennell 2006; Crouch 2013). In particular, we establish that the fundamental motives of mate acquisition play an important role in eliciting more predictable gendered differences with regard to risk-taking tourism behaviors, whereby their activation selectively increases the propensity for risk-taking tourism among men rather than women. Across the results of four experimental studies, we show that this effect is consistent across different domains of travel decision-making, including the choice of high-risk travel activities (study 1A and 1B), adventurous destination choices (study 2), and the evaluation of adventurous appeals in travel advertising (study 3).
Our study contributes to tourism research in several ways. First, Crouch (2013) has drawn attention to the potential of evolutionary psychology as a means to advance tourism research, offering a useful conceptual framework to study evolution-based explanations of travel and tourism behavior. Within this framework, travel and tourism behavior consists of evolutionary drivers, enablers, and expressions. Drivers are evolutionary forces that favor travel as a means of promoting the reproductive and survival success. Evolutionary enablers are travel consumption behaviors and choices that provide people with the means to respond to the drivers. Finally, expressions are the consequential manifestations of travel behaviors that result from the drivers and enablers. Informed by this framework, we advance Crouch’s (2013) theoretical account by establishing empirically a particular causal relationship between the evolutionary drivers (mate acquisition motives), enablers (risk taking), and tourism expressions (gender-specific preferences for high-risk activities, adventurous destination choices, and advertising of adventurous travel).
Moreover, previous studies have largely associated risk-taking propensities among travelers with particular personality traits, such as sensation-seeking (Lepp and Gibson 2008; Pizam et al. 2004) and thrill and adventure seeking (Pizam, Reichel, and Uriely 2001). Resulting from this perspective is an implicit assumption that the risk-taking propensity is a relatively stable characteristic of travelers—since personality traits are also relatively stable constructs. Contrary to this view, and informed by the notion that people’s desire for travel could be underpinned by the fundamental motives that have evolved from sociobiological origins—such as signaling the individual’s mastery in exploring new territories (Crouch 2013)—we provide an alternative explanation underpinning risk-taking tourism behaviors. We establish that choosing high-risk travel activities, traveling to adventurous destinations, and even purchasing travel packages that are accompanied by adventurous advertising appeals, are the evolutionary expressions of achieving salient positive differentiation (Griskevicius et al. 2007). Accordingly, we argue that—unlike the tendencies that result from personality traits—the risk-taking propensities in traveling as evolutionary expressions are not stagnant. Instead, such propensities can be activated strategically to satisfy particular evolutionary challenges—mate acquisition motives.
Subsequently, our findings advance research on the gendered differences in tourism behaviors by complementing extant theories that have largely focused on proximate explanations (e.g., Carr 2002; Pizam et al. 2004). For instance, Pizam et al. (2004) notes that scoring high on RSS characteristics is more evident among men than among women, offering useful insights into the gendered differences in risk-taking tourism. However, our study extends this proximate relationship with an ultimate explanation (Scott-Phillips, Dickins, and West 2011), whereby we highlight that these observable gendered differences in psychological systems can be more holistically explained—and accordingly, predicted—when we take into account the activation of mate seeking. That is, consistent with “costly signaling theory” (Zahavi 1975), partaking in risk-taking activities can be conceived as an opportunity to display the individual’s ability to incur costs, which increases his or her prestige. The results of our study provide empirical evidence that such “costly signaling” behaviors in tourism are more evident among men than women, when their mate-seeking motives are activated.
The study also offers several recommendations for tourism management and practice. First, an obvious implication derived from our findings is that demographic characteristics (e.g., age, gender) and travel characteristics (e.g., single travelers, with partners) should be conjointly taken into consideration. In relation to this, important insights can be derived by contrasting the divergent results that we obtained from study 1A and study 1B. We found that merely eliciting romantic associations does not selectively increase the desire for risk taking during a trip. It is only when the mate-seeking motives are activated that men tend to display more preference for high-risk travel. Thus, men travelers are not always more risk taking compared to women. Therefore, travel agents should consider gender in the context of other traveler’s characteristics—such as age, whether they travel alone, and whether they display a tendency to be conspicuous (e.g., appear fashionable), when recommending adventurous tourism destinations.
In the same vein, the results of our study offer implications for travel advertising and brand positioning. Travel marketers can use different advertising appeals and positioning strategies based on the relevant characteristics of their target consumers. For instance, our results are able to explain why marketing campaigns and slogans that convey adventurous and risky messages, such as Las Vegas as the “Sin City,” work successfully for travel destinations that target predominantly young men consumers. Another implication could be related to the use of sexual appeals in promoting adventurous tourism destinations. Specifically, since the use of sexualized advertisements for adventurous destinations is likely to activate mate-seeking motives and convey higher risk, such advertising would be more effective in targeting men than women.
Although our study offers a novel contribution, future research should address several limitations. First, it is important to acknowledge that our aim is by no means to suggest that evolutionary psychology, more broadly, and mating motives, more specifically, should be regarded as the only perspective to investigate risk-taking tourism. Rather, we illustrate how this evolution-based theorizing approach enables tourism researchers to develop novel insights about various issues pertaining to modern travel behaviors that were absent heretofore. In this endeavor, our study shows how this sociobiological approach allows us to reconcile previous inconsistencies within the extant theory regarding whether and when men and women vary in terms of their risk-taking travel behaviors.
Second, given that the primary objective of our study was to establish a causal relationship between the activation of mate-seeking motives and gendered differences in travel behaviors, we used the established scenario-based method that is suitable for experimental studies to manipulate mating motives (Griskevicius, Cialdini, and Kenrick 2006; Griskevicius et al. 2007; Kim and Kim 2016). However, there are some limitations. For instance, it could be argued that our control condition, which asked participants to choose a street that they perceived to be the most desirable to walk on, could activate the romantic concept. Therefore, future research could explore other ways to activate mate-seeking motives, which will extend the external validity of our results in more naturalistic settings. Further research using longitudinal design, field observations, and secondary data including the actual travel behaviors could also be useful in this endeavor. In addition, our studies were limited to adult participants younger than 40 years, which limits the generalizability of our findings to other age groups.
Third, our study conceptualized mating activation as an independent variable and gender as a moderating variable. Alternatively, it is possible to categorize gender as an independent variable and mating activation as a moderator. Future research can explore whether this alternative conceptualization could offer additional insights regarding risk-taking propensities in tourism behaviors. Finally, there are other forms of travel, such as sustainable travel, luxury travel, or status-related travel that could be investigated from the evolution-based perspective. For example, previous research suggests that women are more likely to achieve salient positive differentiation by emphasizing generosity and engaging in prosocial behaviors (Griskevicius et al. 2007). Thus, it is possible that when mating motives are activated, women may show more preference for sustainable tourism compared to men. Another area of future research could be also to explore the impact of mating acquisition motives on the issues pertaining to dark and deviant tourism (e.g., Oppermann 1999) or Hedonism II (Sheller 2004), which were beyond the scope of our article.
In conclusion, we hope that this study, with its emphasis on fundamental motives, gendered differences, and risk-taking propensities in tourism, will offer new insights for the researchers and practitioners who are interested in adopting an evolution-based perspective in travel research.
Footnotes
Author Note
Both authors contributed equally to this work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.