
Abstract
We used signal detection theory to evaluate spatial recognition memory utilizing a behavioral test hypothesized to tax pattern separation. Correlations with standardized neuropsychological tests also were examined. Healthy young (n = 40) and older (n = 30) adults completed a spatial recognition memory test involving high- and low-similarity conditions. Using d’ as the dependent variable, we found that older adults were significantly impaired relative to young adults on the high- and low-similarity conditions (ps < .05). Both groups performed significantly better in the low-similarity condition compared to the high-similarity condition (p < .05), with young adults exhibiting greater improvement relative to older adults. We also found that young adults may rely on spatial attention abilities when performing our test, while older adults might rely on memory and executive function abilities. These findings indicate that young and older adults may utilize different cognitive abilities when performing certain spatial memory tests.
Age-related changes in spatial memory have been well documented in both humans (reviewed by Iachini, Iavarone, Senese, Ruotolo, & Ruggiero, 2009) and animals (reviewed by Sharma, Rakoczy, & Brown-Borg, 2010). Spatial memory impairment also has been well documented in older adults diagnosed with Alzheimer’s disease (AD; Kessels, Boekhorst, & Postma, 2005; Sahakian et al., 1988) and mild cognitive impairment (MCI; Alescio-Lautier et al., 2007; deIpolyi, Rankin, Mucke, Miller, & Gorno-Tempini, 2007; Hort et al., 2007), which is considered to be a transition stage between healthy aging and AD (Petersen, 2004). Deficits in spatial memory have been suggested to serve as an early indicator of AD (Buccione et al., 2007; deIpolyi et al., 2007; Hort et al., 2007). Spatial memory impairments in older adults may result from changes in a variety of brain regions including the hippocampus, temporal lobes, and the frontal-parietal network (Iachini et al., 2009). In particular, age-related spatial memory decline has been suggested to result from age-related changes in the hippocampus across species (reviewed by Barnes, 1988). Recently, studies have begun to examine how age-related changes in particular subregions of the hippocampus may affect specific mnemonic processes, such as pattern separation in older adults.
Pattern separation has received considerable attention in the learning and memory literature as a neural mechanism to reduce interference, similarity, or ambiguity among memory representations, thereby increasing the likelihood of accurate encoding and subsequent retrieval. Pattern separation is described by computational models as the orthogonalization of similar inputs into nonoverlapping, distinction representations (O’Reilly & McClelland, 1994; Treves & Rolls, 1994). There is considerable evidence that the dentate gyrus (DG) and CA3 hippocampal subregions play a critical role in pattern separation (for reviews, see Gilbert & Brushfield, 2009; Kesner & Rolls, 2015; Rolls, 2013; Schmidt, Marrone, & Markus, 2012; Yassa & Stark, 2011).
Age-related structural and functional changes have been well documented in the human hippocampus (J. S. Allen, Bruss, Brown, & Damasio, 2005; Driscoll & Sutherland, 2005; Good et al., 2001; Raz et al., 2005; Small, Tsai, DeLaPaz, Mayeux, & Stern, 2002; Walhovd et al., 2010). In particular, the DG subregion may be highly susceptible to these age-related changes (Small et al., 2002). Doxey and Kirwan (2014) used functional magnetic resonance imaging (fMRI) and diffusion tensor imaging (DTI) to measure functional and structural correlates of behavioral pattern separation in the hippocampus and medial temporal lobe. The size of left hemisphere DG/CA3 regions was found to be the strongest predictor of performance, aside from age, on a behavioral task hypothesized to require pattern separation. However, resting functional connection strengths and diffusion in white matter tracts did not significantly predict task performance (Doxey & Kirwan, 2014). Age-related changes in the perforant pathway input to the DG have been identified using high-resolution fMRI and ultrahigh-resolution DTI (Yassa, Mattfeld, Stark, & Stark, 2011). It was found that decreased pattern separation activity in the DG/CA3 regions of older adults was associated with structural changes in the perforant pathway. These changes were hypothesized to weaken the processing of new information and strengthen the processing of stored information (Yassa, Mattfeld, et al., 2011), which may result in less efficient pattern separation. A recent study used ultrahigh in-plane resolution DTI to examine the specificity of perforant path integrity, as it relates to mnemonic discrimination (possible measure of pattern separation) relative to other cognitive constructs and other white matter tracks across the adult lifespan (Bennett & Stark, 2016). The authors found that integrity of the perforant path and other white matter tracts in the medial temporal lobe declined with increased age. Mnemonic discrimination was found to relate to integrity of the perforant path and other tracts within the temporal lobe, indicating that mnemonic discrimination may be mediated by a broader network within the medial temporal lobe.
There is considerable evidence indicating that older adults are impaired relative to young adults on behavioral tasks hypothesized to tax pattern separation for visual objects (Doxey & Kirwan, 2014; Pidgeon & Morcom, 2014; Reagh et al., 2016; Stark, Yassa, Lacy, & Stark, 2013; Toner, Pirogovsky, Kirwan, & Gilbert, 2009; Yassa, Lacy, et al., 2011; Yassa, Mattfeld, et al., 2011), spatial locations (Holden, Hoebel, Loftis, & Gilbert, 2012; Reagh et al., 2014, 2016; Stark, Yassa, & Stark, 2010), temporal order (Roberts, Ly, Murray, & Yassa, 2014; Rotblatt et al., 2015; Tolentino, Pirogovsky, Luu, Toner, & Gilbert, 2012), verbal stimuli (Ly, Murray, & Yassa, 2013), and emotional information (Leal & Yassa, 2014). The majority of studies examining age-related changes in pattern separation have used a variant of a continuous recognition memory test for visual objects originally developed by Kirwan and Stark (2007). However, far less is known about the effects of age on tests hypothesized to assess spatial pattern separation. Given that spatial memory deficits are well documented in older adults (Iachini et al., 2009) and may be an early marker of AD (Buccione et al., 2007), additional research is needed to assess how age-related changes in pattern separation may affect spatial memory in old age. We are only aware of four studies that have examined differences between young and older adults on behavioral tasks hypothesized to assess spatial pattern separation (Holden et al., 2012; Reagh et al., 2014, 2016; Stark et al., 2010). All four studies have used different tests to measure behavioral pattern separation, but the findings have shown consistently that at least a subset of older adults show impairments relative to young adults on the tests. In addition, similar studies have been conducted to examine what is referred to as transposition distance effects in aging and have reported that older adults demonstrate larger effects than young adults (P. A. Allen et al., 1998). This effect was attributed to elevated levels of internal noise in older adults relative to younger adults (P. A. Allen, 1990), which was later supported using a molar entropy model (P. A. Allen et al., 1998). However, none of these studies have examined relationships between performance on these tests hypothesized to assess pattern separation and a large battery of standardized neuropsychological tests that assess a variety of cognitive domains. Therefore, it is largely unknown how performance on these tests relates to constructs that are theoretically related, or unrelated, to the construct of pattern separation. Furthermore, additional research is needed to examine whether performance on these tests relates to the same or different constructs (e.g., attention, memory, executive function) in young and older adults. In this study, we examined relationships between performance on our test and a large battery of standardized neuropsychological tests assessing a variety of cognitive domains.
A previous study (Holden et al., 2012) used a delayed-match-to-sample test involving the manipulations of the distance between two spatial locations to examine behavioral pattern separation, with mean percent correct serving as the primary dependent variable. However, the original task did not provide a measure of recognition memory. Therefore, it was difficult to assess potential age-related deficits in spatial pattern separation as separate from a general deficit in spatial memory. In this study, we modified the original task into a recognition memory test format so that signal detection theory (d’) could be used to analyze the behavioral data. A d’ score reflects the absolute difference in standard deviation units between a participant’s hit rate and false-positive rate (Macmillan & Creelman, 1991). Therefore, one advantage of using signal detection theory is that it provides a single summary statistic to characterize overall accuracy (Murdock, 1982), rather than one measure for old items and a second measure for new items. Three recent studies have used signal detection theory to evaluate performance on visual memory tests hypothesized to measure behavioral pattern separation (Loiotile & Courtney, 2015; Reagh et al., 2016; Stark, Stevenson, Wu, Rutledge, & Stark, 2015). On these tasks, participants are asked to discriminate between previously encoded stimuli, novel stimuli, and stimuli that are highly similar to those previously encoded (lures). Using signal detection theory, these recent studies indicate that d’ (or a related measure such as da), a measure of a participant’s ability to distinguish between previously encoded “old” stimuli and novel stimuli, might be useful in characterizing performance on behavioral pattern separation tasks (Loiotile & Courtney, 2015; Stark et al., 2015). Reagh et al. (2016) also used signal detection theory in a recent study comparing object and spatial mnemonic discrimination. Therefore, we used d’ as our dependent measure to examine age-related differences in performance on our spatial recognition memory test, hypothesized to tax behavioral pattern separation.
Method
Participants
Study participants (n = 70) included 30 healthy older adults over 60 years of age (M = 71.07 years, SD = 7.53) and 40 younger adults 18 to 25 years of age (M = 19.50, SD = 1.70). A one-way analysis of variance (ANOVA) test revealed that the older adults (M = 15.37, SD = 2.06) attained significantly more years of education, F(1, 68) = 33.79 p < .01, than the young adults (M = 13.13, SD = 1.14). In addition, raw scores on the Wide Range Achievement Test—Fourth Edition (WRAT-4; Wilkinson & Robertson, 2006) were significantly higher for older adults (M = 64.07, SD = 5.07) than young adults (M = 61.50, SD = 4.35), F(1, 68) = 5.18 p < .05. The WRAT-4 is a measure of word reading ability and a proxy measure of educational level. A χ2 analysis did not reveal a significant sex difference, χ2 (1, N = 70) = .81, p = .37, between the young (58% female) and older (47% female) adult groups. The young adults were recruited from a pool of undergraduate students at San Diego State University and the older adults were community-dwelling individuals recruited from the San Diego community. All procedures were approved by the Institutional Review Board at San Diego State University, and all participants provided informed consent prior to participation in this study. All participants underwent a near and far visual screening test before participation and all except one had corrected vision that fell between 20/20 and 20/40.
Neuropsychological and Mood Measures
Participants completed a battery of standardized neuropsychological tests that included measures for the following constructs: (a) Memory—Hopkins Verbal Learning Test-Revised (HVLT-R; Brandt & Benedict, 2001) and Brief Visuospatial Memory Test-Revised (BVMT-R; Benedict, 1997); (b) Executive Functioning—Verbal Fluency and Trail Making tests from the Delis Kaplan Executive Function System (D-KEFS; Delis, Kaplan, & Kramer, 2001); (c) Attention—Digit Span and Spatial Span subtests of the Wechsler Memory Scale-Third Edition (WMS-III; Wechsler, 1997); (d) Visuospatial Processing—Benton Judgment of Line Orientation Test (Benton, Sivan, Hamsher, Varney, & Spreen, 1994); and (e) Language—Word Reading subtest of the WRAT-4 (Wilkinson & Robertson, 2006) and Boston Naming Test (Kaplan, Goodglass, & Weintraub, 1983). Raw scores were used in all correlation analyses.
To assess global cognitive functioning, older adult participants were administered the Dementia Rating Scale-2 (DRS-2; Jurica, Leitten, & Mattis, 2001). The average DRS-2 score for older adults was 139.17 (SD = 3.94). To screen for depression, older adults completed the Geriatric Depression Screening Scale (GDS; Yesavage, Brind, Rose, & Lum, 1983) and young adults completed the Beck Depression Inventory-II (BDI-II; Beck, Steer, Ball, & Ranieri, 1996). All young adults had a BDI-II score lower than 20 (minimal to mild depression) and all older adults had a GDS score of 5 or lower (normal) with the exception of one participant whose score was 7 (suggestive of depression). The average GDS score for older adults was 1.70 (SD = 1.86), and the average BDI score for young adults was 6.25 (SD = 5.12).
Spatial Recognition Memory Task
Participants were administered a spatial recognition memory task that was based on a delayed-match-to-sample test developed to assess the effect of similarity or interference on spatial memory and potentially spatial pattern separation (Holden et al., 2012). During the new task, participants were seated approximately 40 cm in front of a computer monitor that had a 15-cm black border affixed around the perimeter of the screen (see Figure 1) to minimize the possibility of using a specific visual cue on the monitor (e.g., button, light) to remember the spatial location of stimuli. The monitor was placed against a large plain wall to further minimize the use of an environmental room cue (e.g., a light switch) to identify the location of stimulus on the screen. As shown in Figure 1, each trial consisted of a sample phase followed by a choice phase. During the sample phase, participants viewed a gray circle measuring 1.7 cm in diameter that appeared on the computer screen for 5 s. The circle appeared in one of 18 possible locations within a fixed nonvisible horizontal line across the middle of the screen. There was a 10-s delay between the sample phase and choice phase during which participants were required to look away from the screen and read a designated string of random letters to prevent fixation of the eyes on the location of the original circle from the sample phase. After the 10-s delay, a tone was sounded to signal the beginning of the choice phase. During the choice phase, a new gray circle appeared, either in the same location (“same” trial) or in a different location (“different” trial) relative to the sample phase. On “different” trials, the choice phase circle was placed in a location that differed from the sample phase location by one of four possible spatial separations (0.5, 1.0, 1.5, or 2.0 cm), to one side or the other of the sample phase circle location. Participants were to say “same” if they thought the choice phase circle was in the same location as the sample phase circle or “different” if they thought the choice phase circle was in a different location than the sample phase circle. Participants were allowed 5 s to make a response. After 5 s, the words “Next Trial” appeared on the computer screen to indicate that a new trial was about to begin.
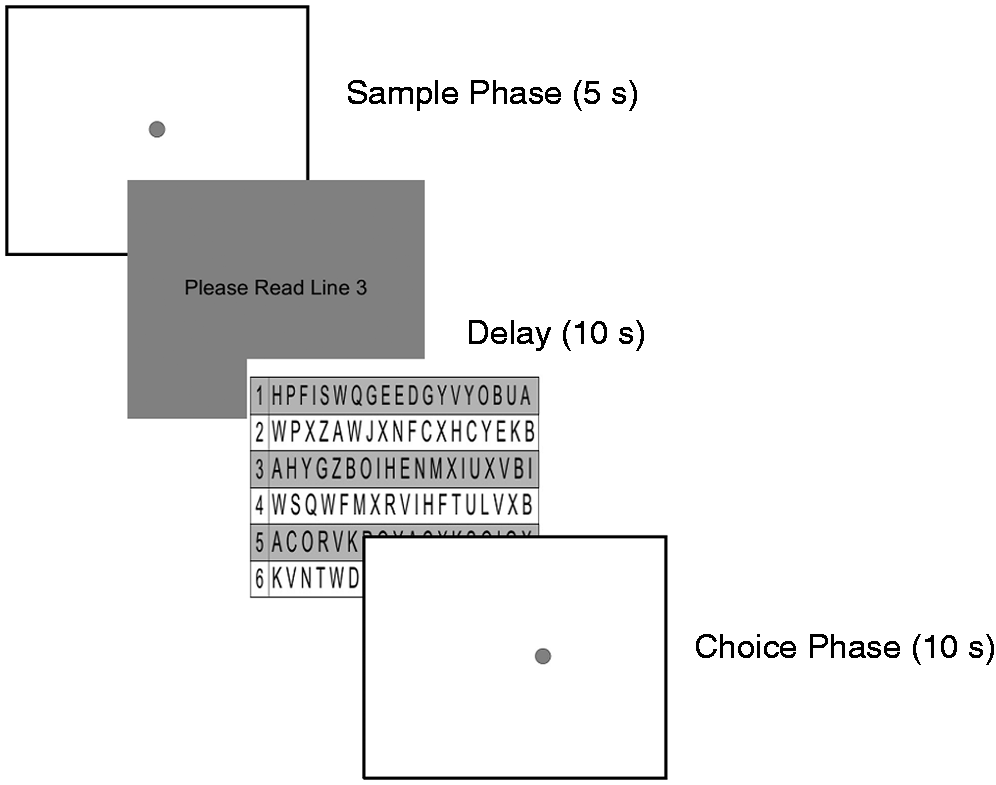
A schematic of the testing procedure showing a sample phase stimulus, the delay instructions, and a choice phase stimulus (B).
The task consisted of a total of 72 trials, including 24 recognition (“same”) trials and 48 discrimination (“different”) trials. The 48 discrimination trials consisted of 12 trials for each of the four spatial separations. Each group of 12 trials was balanced across the entire width of the screen to ensure that there was not an unintentional bias toward one particular area on the screen. To minimize fatigue effects, the 72 total trials were split into two sets of 36 trials. The two sets were identical in design. The order of the two sets of 36 trials was pseudorandomized, with the restriction that there were no more than three consecutive trials of the following: (a) spatial separation or (b) choice circle on a particular side. In addition, the pseudorandomization imposed a restriction that there would be no more than four consecutive same or different trials. Each set of 36 trials took approximately 12 to 15 minutes to complete, with the entire test battery lasting approximately 2.5 hours.
The recognition memory data were analyzed using standard signal detection theory procedures to measure sensitivity (Macmillan & Creelman, 1991). The raw number of hits and false-positive errors were transformed into conditionalized rates. Since measures of performance using signal detection theory are undefined for hit rates equal to 1 or false-positive rates equal to zero, a linear transformation was applied to the calculation of hit rates and false-positive error rates as suggested by Upton (1978) for log-linear models using the following equations: Hit Rate (HR) = [(number of hits + .05)/(number of targets + 1)] and False-Positive Error Rate (FR) = [(number of false-positive errors + .05)/(number of distractors + 1)]. The z-transformation was used to convert the hit rates and false-positive error rates into a z-score. The sensitivity measure of signal detection theory (d’) was then calculated using the formula d’ = [z(HR) − z(FR)], which provides an indication of the strength of the signal relative to the noise. A d’ score reflects the absolute difference in standard deviation units between a participant’s hit rate and false-positive rate (Macmillan & Creelman, 1991). A second parameter c was also calculated, which provides a measure of response bias to indicate if a participant is particularly liberal or conservative in providing “yes” responses.
Results
Using d′ as the dependent variable, a 2 × 2 mixed model ANOVA was used to analyze the data with age (older adults, young adults) as a between-group factor and interference (high, low) as a within-group factor. Smaller spatial separations (0.5 and 1.0 cm) on “different” trials were hypothesized to result in greater similarity or interference than larger separations (1.5 and 2.0 cm) and hence place greater demands on pattern separation. Therefore, performance on trials involving spatial separations of 0.5 or 1.0 cm was averaged to form the high-interference condition and performance on trials involving spatial separations of 1.5 or 2.0 cm was averaged to form the low-interference condition. Reducing the number of interference conditions from four to two increased the number of trials per condition, which is important when using signal detection theory and produces a more robust measure of performance.
The analysis revealed a statistically significant main effect of age-group on test performance, F(1, 68) = 11.91, p = .001, η2 = 0.15, with young adults outperforming older adults overall. There was also a significant main effect of interference on performance, F(1, 68) = 27.19, p < .001, η2 = 0.27, with higher performance in the low-interference condition compared to the high-interference condition. In addition, there was a significant Age-Group × interference interaction, F(1, 68) = 4.07, p < .05, η2 = 0.04. As shown in Figure 2, a Newman-Keuls post hoc comparison test of the age-group × interference interaction revealed that older adults were significantly impaired relative to young adults on both the high- and low-interference conditions (ps < .05), and these effects were associated with medium to large effect sizes (high interference: d = .64, low interference: d = 0.91). Both groups performed significantly better on low-interference trials relative to high-interference trials (ps < .05), but young adults exhibited greater improvement relative to older adults (26% larger effect size). The increases in performance from the high- to low-interference conditions in both groups were associated with medium effect sizes (older adults: d = 0.56; younger adults: d = 0.76). Single sample t tests revealed that scores on c (a measure of response bias in signal detection theory) did not differ from zero in older adults on high-interference t(29) = −1.24 p = .23 or low-interference t(29) = .75 p = .46 trials. Given that prior studies have reported that older adults may have a more liberal response bias (P. A. Allen, 1990), our findings suggest that the older adults in this study did not show a liberal or conservative response bias.

Mean (±SE) d′ scores for young and older adults on the high- and low-interference conditions on the spatial recognition memory test.
Pearson’s r correlations were conducted to examine relationships between performance on the standardized neuropsychological measures and the high- and low-interference conditions of the spatial recognition memory task in young and older adults. All correlation coefficients and associated p values are presented in Table 1. The standardized test battery included measures from the following domains: verbal and visual memory, attention, executive functioning, visuospatial perception, language, reading, and motor speed.
Pearson r Correlations for Young (n = 40) and Older (n = 30) Adults Between Performance on High- and Low-Interference Conditions of the Spatial Recognition Memory Test andNeuropsychological Tests Measuring Memory, Attention, Executive Function, Visuospatial Perception, and Language.
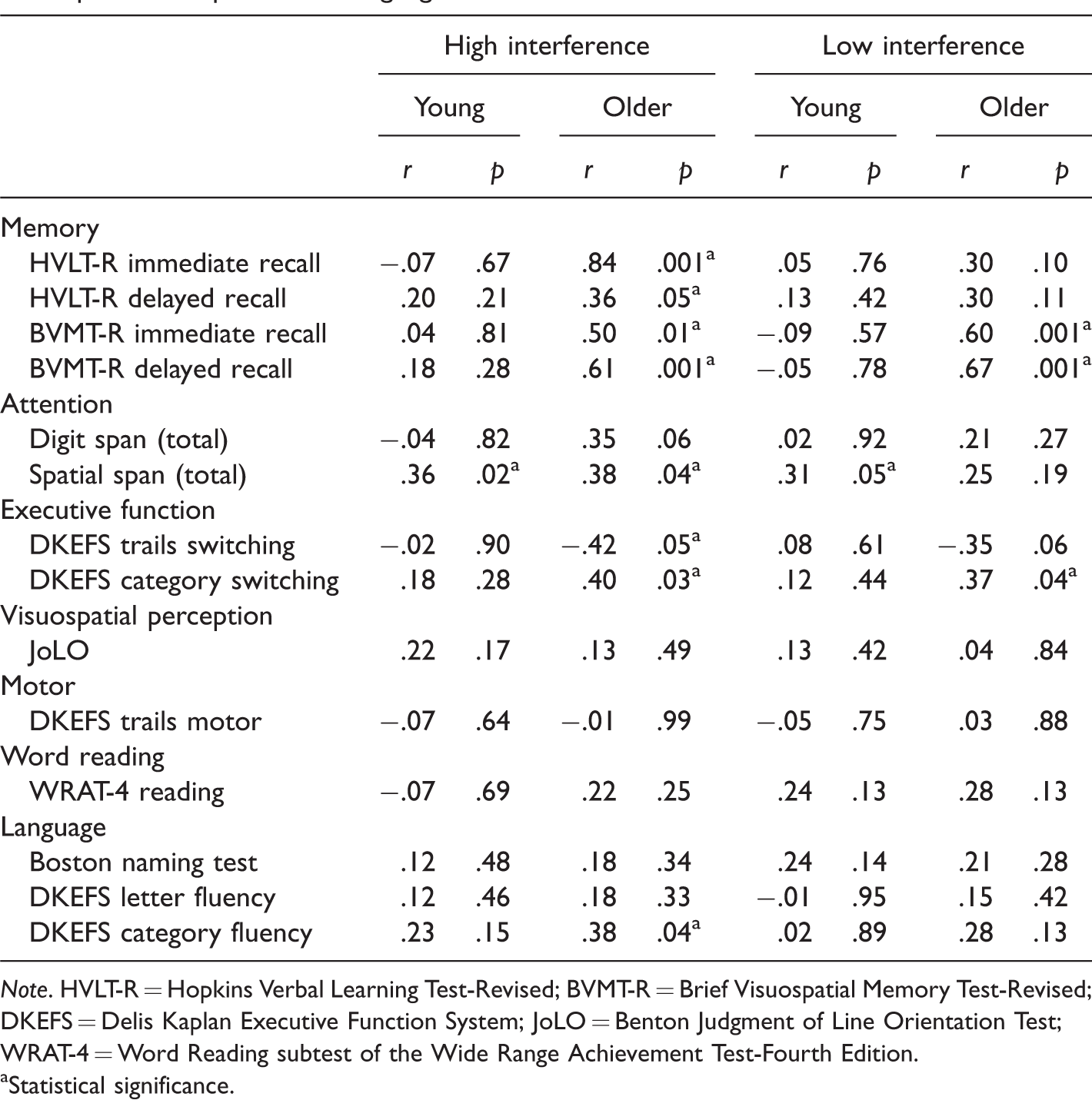
Note. HVLT-R = Hopkins Verbal Learning Test-Revised; BVMT-R = Brief Visuospatial Memory Test-Revised; DKEFS = Delis Kaplan Executive Function System; JoLO = Benton Judgment of Line Orientation Test; WRAT-4 = Word Reading subtest of the Wide Range Achievement Test-Fourth Edition.
aStatistical significance.
In older adults, scores on the DRS-2 did not correlate significantly with performance on the low- (r = .20, p = .28) or high-interference (r = .22, p = .25) conditions of the spatial recognition memory test. GDS scores in older adults did not correlate significantly with performance on the low- (r = −.02, p = .91) or high-interference (r = −.18, p = .34) conditions of the spatial recognition memory test. Similarly, BDI-II scores in young adults did not correlate significantly with performance on the low- (r = −.2.0, p = .22) or high-interference (r = −.14, p = .40) conditions of the spatial recognition memory test.
One-way ANOVA tests were conducted to assess age-group differences on all standardized neuropsychological tests and the spatial recognition memory test. The data from young and older adults along with the results of the statistical analyses and associated effect size estimates are reported in Table 2.
Mean (SD) d′ Scores on the Spatial Recognition Memory Test and Raw Scores on Standardized Neuropsychological Tests of Young (n = 40) and Older (n = 30) Adults, Along With the Results of One-Way Analysis Of Variance Tests Comparing Age-Group Differences and Associated Effects Size Estimates.
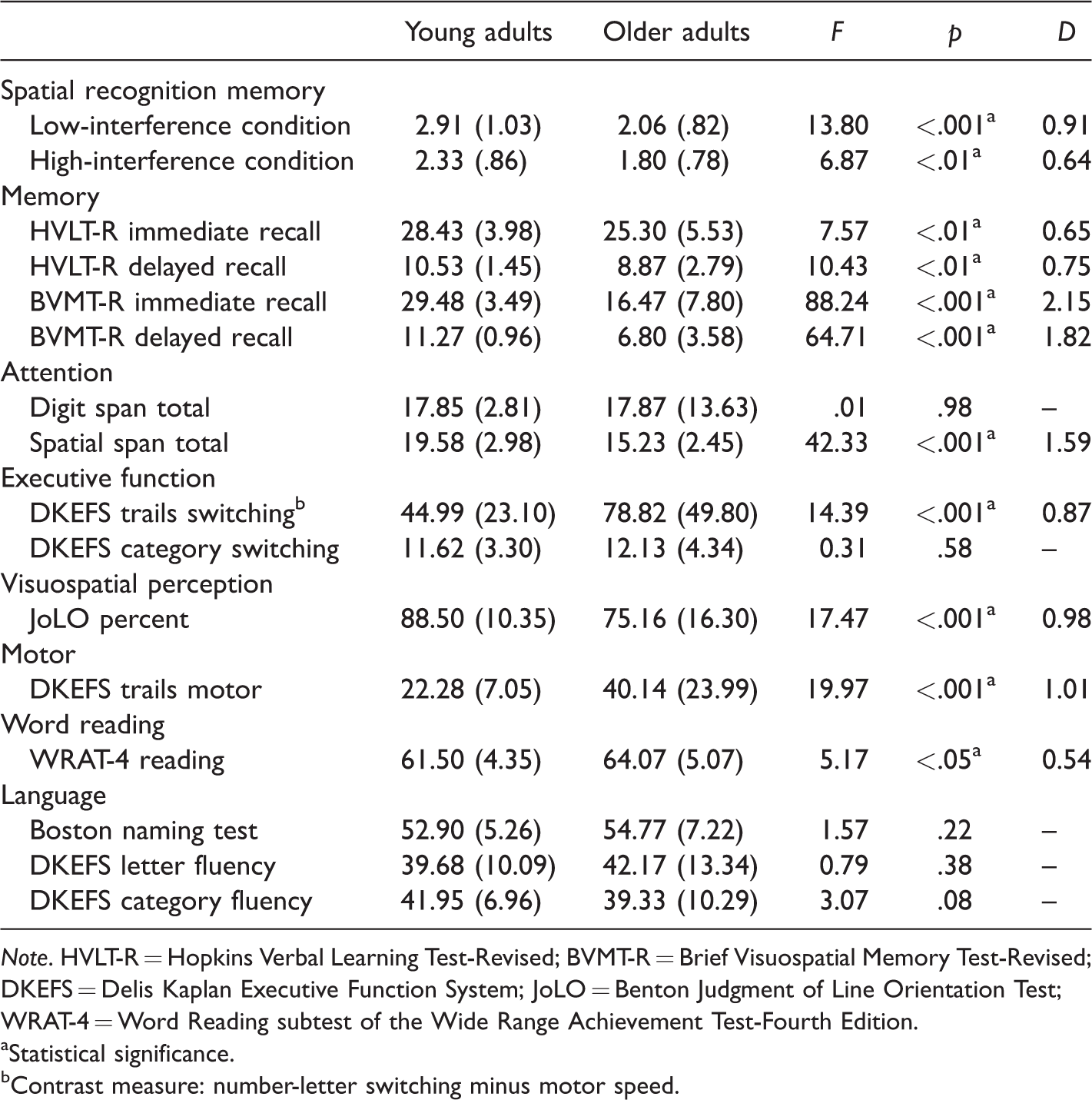
Note. HVLT-R = Hopkins Verbal Learning Test-Revised; BVMT-R = Brief Visuospatial Memory Test-Revised; DKEFS = Delis Kaplan Executive Function System; JoLO = Benton Judgment of Line Orientation Test; WRAT-4 = Word Reading subtest of the Wide Range Achievement Test-Fourth Edition.
aStatistical significance.
bContrast measure: number-letter switching minus motor speed.
Discussion
One aim of this study was to use signal detection theory to assess age-related differences on a spatial recognition memory test hypothesized to assess spatial pattern separation. It was hypothesized that trials involving smaller spatial separations (i.e., 0.5 cm and 1.0 cm separation trials) resulted in increased similarity or interference, and hence higher demand for pattern separation, relative to trials involving larger spatial separations (i.e., 1.5 cm and 2.0 cm separation trials). We found that both young and older adults performed significantly better on trials with lower levels of interference compared to trials with higher levels of interference. However, young adults exhibited greater improvement in performance on low-interference trials compared to high-interference trials than did older adults (26% larger effect size). The findings are similar to those reported in a recent study published by Reagh et al. (2016) using a different spatial memory test and a dependent variable related to d’, referred to as da. Therefore, the data add to a small, but growing, number of studies providing evidence that spatial pattern separation may be less efficient in older adults relative young adults (Holden & Gilbert, 2012; Holden et al., 2012; Reagh et al., 2014, 2016; Stark et al., 2010). Although a number of studies have assessed age-related differences on visual object tests hypothesized to tax pattern separation, there is a need to develop behavioral tests to assess pattern separation in other domains. Given the connection between spatial memory impairment and AD (Buccione et al., 2007; deIpolyi et al., 2007; Hort et al., 2007), tests such as the present one may be used in conjunction with standardized neuropsychological measures to help differentiate neurocognitive changes associated with normal aging, MCI, and AD. A recent study from our laboratory found that a subset of healthy, nondemented older adults genotyped as apolipoprotein E-ε4 (APOE-ε4) carriers were significantly impaired relative to APOE-ε4 noncarriers and young adults on the delayed-match-to-sample version of the present test (Sheppard et al., 2015). Given that the APOE-ε4 is a genetic risk factor for AD, these findings highlight the potential utility of the present test in assessing cognitive deficits in individuals at risk for AD and potentially in a prodromal stage of the disease.
Our current findings, using a recognition memory test, are highly consistent with a previous study (Holden et al., 2012), which utilized a similar delayed-match-to-sample test. In both studies, it was found that young and older adults improved as interference, and the hypothesized demand for pattern separation, decreased. However, both studies found that young adults outperformed older adults. We feel it is important to note that our effects sizes for age-group differences using the new spatial recognition memory test in this study were 52.38% larger on high-interference trials (d = .64) compared to the previous Holden et al. (2012) study (d = .42). We modified the original test (Holden et al., 2012) into a recognition memory format so that we could assess general spatial recognition memory on same trials relative to recognition memory on the trials involving varying degrees of interference. The original task did not provide a measure of recognition memory. Therefore, it was difficult to assess potential age-related deficits in spatial pattern separation as separate from a general deficit in spatial memory. In addition, the recognition memory format enabled us to use signal detection theory to analyze the data, an approach that has been used in recent studies to assess age-related differences on the mnemonic similarity tests, which are hypothesized to assess pattern separation for visual object (Stark et al., 2015) or spatial (Reagh et al., 2016) information. A recent study reported that mnemonic discrimination in older adults may be more impaired for visual object information than for spatial information (Reagh et al., 2016). Therefore, future studies are needed to compare the sensitivity of the present spatial recognition memory test to other behavioral tests hypothesized to assess pattern separation for nonspatial information.
A second aim of this study was to examine relationships between performance on our spatial recognition memory test and standardized neuropsychological tests that assess various domains of cognition. In young adults, we found that performance on the spatial recognition memory test was significantly correlated with performance on a spatial attention test (spatial span total score) but not an auditory attention test (digit span total score) in both the high- and low-interference conditions. In older adults, performance on our test was also found to correlate with the spatial attention measure (spatial span test), but only in the high-interference condition. Performance in the high-interference condition was also found to correlate significantly with standardized measures of verbal and visuospatial memory for both immediate and delayed recall. In addition, performance was significantly correlated with standardized measures of executive functioning (switching condition of the trail making test and the fluency test). Similar results were found for the low-interference condition in older adults, with the exception that verbal memory (HVLT-R) and spatial attention (spatial span) did not correlate with performance on this condition of the spatial recognition memory test. These findings suggest that performance on our test is specifically related to the construct of spatial attention in young adults. However, in older adults, performance on our test also is related to the constructs of visuospatial memory and executive functioning. Although young adults may rely on spatial attention abilities when performing our test, older adults might be relying on more memory and executive abilities, particularly when interference is increased.
Performance on our test was not found to be related to performance on tests assessing cognitive constructs not theoretically related to the construct of spatial recognition memory, including visuospatial perception, motor functioning, word reading, language, or auditory attention. These findings illustrate that any age-related deficits on our test were not solely due to impairments in visuospatial perception, motor functioning, or general attention in older adults. Furthermore, scores on a measure of global cognitive functioning (DRS-2) in older adults did not correlate with performance on the high- or low-interference conditions of the spatial recognition memory test, indicating that age-related deficits on our test were not likely due to global cognitive impairment in the older adults. Similarly, scores on standardized measures of depression did not correlate with performance on the spatial recognition memory test in young or older adults. All participants were screened for depression and none of the participants endorsed more than mild depressive symptoms (based on BDI-II and GDS scores). This is important given that recent studies have reported a negative relationship between depression scores and scores on tests hypothesized to assess pattern separation (Shelton & Kirwan, 2013). All participants completed a near and far visual screening test, and all except one older adult had corrected vision that fell between 20/20 and 20/40. This minimized the possibility that decreased visual acuity contributed to age-related differences on the spatial recognition memory test. Furthermore, performance on the Benton Line Orientation Test, a measure of visuospatial perception, was not found to be correlated with performance on the spatial recognition memory test in either group. The older adults scored significantly higher on the WRAT-4, which is a proxy measure for educational level. Therefore, age-related deficits on the spatial recognition memory test in older adults are not likely due to lower education levels in this group. In addition, the young and older adult groups did not differ in sex, which is another important demographic variable. Taken together, these data provide evidence that demographic variables, visual acuity, depressive symptoms, and global cognitive function were assessed and controlled for in this study.
On our spatial recognition memory test, young adults were found to outperform older adults in both the low- and high-interference conditions, and the differences were associated with large (.91) and moderate (.64) effect sizes, respectively. The effects sizes for age-related differences on the spatial recognition memory test were comparable to (high-interference condition) or larger than (low-interference condition) the effect sizes for the HVLT-R, which is a test of verbal memory. However, the effects sizes for the spatial recognition memory test were lower than the effect sizes for age-related differences on the BVMT-R, a test of visuospatial memory, and the Spatial Span test, a measure of spatial attention. The effects sizes for the low-interference condition of the spatial memory test were comparable to the effect sizes for the DKEFS Trail Making Test switching condition, a measure of executive functioning, and the Benton Judgment of Line Orientation test, a test of visuospatial perception. However, the effect size for the high-interference condition was lower than the effect sizes associated with age-related differences on these measures. In summary, the present results provide evidence that our spatial recognition memory test is comparable to a number of different standardized neuropsychological tests in terms of sensitivity to age-related differences between young and older adults. However, it is important to emphasize that our test assesses an aspect of memory function that is not directly measured by existing standardized tests, which is the effect of interference on spatial memory.
Our results from this study demonstrate that spatial pattern separation, as assessed by a spatial recognition memory test, may be less efficient in older adults compared to young adults. In addition, we found that young adults may rely on spatial attention abilities when performing our test, while older adults may rely more on memory and executive abilities, particularly when interference is increased. These findings indicate that young and older adults may utilize different cognitive abilities when performing our test and potentially other behavioral tests thought to assess pattern separation. The current findings also provide preliminary evidence for convergent validity for our spatial recognition memory test. Although the present findings add to our understanding of age-related changes in spatial recognition memory and pattern separation, studies involving larger samples are needed to characterize performance on this test across the adult lifespan and examine the psychometric properties of the test. Future studies also should examine the incremental value of the present, and other, pattern separation tests in evaluating age-related cognitive impairment in comparison with existing standardized neuropsychological tests.
Footnotes
Acknowledgments
The authors thank Carina Hartley and Kyle Scroggins for their assistance with data collection. The authors also thank all of the participants for their contributions to this study.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Paul E. Gilbert was supported by National Institute on Aging/National Institutes of Health grants R01AG034202 and P30AG059299. Shannon Y. DeJesus was supported by a National Institute of General Medical Sciences/National Institutes of Health grant 5R25GM058906.