
Abstract
Despite progress in attractiveness research, we have yet to identify many fitness-relevant cues in the human phenotype or humans’ psychology for responding to them. Here, we test hypotheses about psychological systems that may have evolved to process distinct cues in the female lumbar region. The Fetal Load Hypothesis proposes a male preference for a morphological cue: lumbar curvature. The Lordosis Detection Hypothesis posits context-dependent male attraction to a movement: lordosis behavior. In two studies (Study 1 N: 102, Study 2 N: 231), we presented men with animated female characters that varied in their lumbar curvature and back arching (i.e., lordosis behavior). Irrespective of mating context, men’s attraction increased as lumbar curvature approached the hypothesized optimum. By contrast, men experienced greater attraction to lordosis behavior in short-term than long-term mating contexts. These findings support both the Lordosis Detection and Fetal Load Hypotheses. Discussion focuses on the meaning of human lordosis and the importance of dynamic stimuli in attractiveness research.
Natural selection should have shaped information-processing systems in sexually reproducing organisms to attend to fitness-relevant phenotypic cues in potential mates, and to regulate mating attraction in response to these cues. Abundant evidence across a wide variety of non-human taxa supports this hypothesis (e.g., Birkhead & Fletcher, 1995; Norris, 1993; Petrie, 1994; Weiss et al., 2011). In humans, this overarching hypothesis has resulted in the discovery of a variety of evolved standards of attractiveness: fitness-relevant phenotypic characteristics that humans systematically find attractive. For example, in women, a small waist coupled with a larger gluteofemoral region is a cue to nulligravidity that is linked to higher stores of docosahexaenoic acid, a limiting factor in infant brain development (see Lassek & Gaulin, 2019). In adaptive alignment with this, men are most attracted to a waist-to-hip ratio associated with nubility, nulliparity, and high levels of this key fatty acid (Lassek & Gaulin, 2019). Research also suggests that an individual’s facial bilateral symmetry (Thornhill & Gangestad, 1999), clear skin (Fink & Penton-Voak, 2002 ), and straight and white teeth (Buss, 2006) are indicators of positive reproductive outcomes of mating with that individual. Evidence suggests that humans have mate preferences for all these features (see Buss, 2006; Fink & Penton-Voak, 2002; Thornhill & Gangestad, 1999; see also Lewis et al., in press, for review).
Despite the empirical contributions that an evolutionary approach has made toward our understanding of human standards of attractiveness, the complete suite of fitness-relevant cues present in the human phenotype remains largely unmapped, as does humans’ psychology for responding to these cues. Our current understanding of these cues and our responses to them is largely limited to the face (e.g., Buss, 2006; Fink & Penton-Voak, 2002 ; Thornhill & Gangestad, 1999); women’s waist and thighs (i.e., waist-to-hip ratio, see Lassek & Gaulin, 2019), men’s muscularity (Frederick & Haselton, 2007), and composite or overall bodily indices that do not refer to a specific component of the morphological phenotype (e.g., symmetry, see Gangestad & Thornhill, 1998; Parsons, 1990). Furthermore, evolutionary research on attractiveness has been almost completely limited to static morphological cues. This nearly exclusive focus on static morphology neglects the fact that movements and other dynamic behaviors can convey important fitness-related information. Even more importantly, some of this information may be communicated exclusively through behavior. Selection should therefore have shaped psychological adaptations to attend to these behaviors.
Here, we (a) focused on a largely unmapped domain of the human phenotype—the female lumbar region, (b) identified both behavioral and morphological cues hypothesized to predict fitness-relevant outcomes, and (c) tested a priori hypotheses about the psychological systems that may have evolved to process these cues.
Lordosis Behavior
A combination of phylogenetic considerations and theoretical reasons suggests that women’s behavioral phenotype in their lumbar region may contain at least one reproduction-related cue. Lordosis behavior 1 (Kuehn & Beach, 1963)—a contraction of the spinal muscles that results in an arching of the lower back—is a cue to receptivity among females of diverse mammalian taxa (Ågmo & Ellingsen, 2003; Beach, 1976; Flanagan-Cato, 2011; Gordon et al., 1977; Nance & Myatt, 1987; Owen et al., 2016; Pfaff & Sakuma, 1979; Pfaus et al., 2003; Rodriguez-Sierra et al., 1975). Historically, lordosis behavior has been regarded as a “reflex” (e.g., Pfaff & Sakuma, 1979) governed by hormones, pheromones, and the vomeronasal organ (Haga et al., 2010). In humans, the vomeronasal organ is vestigial (Zhang & Webb, 2003); hormones, while important, do not directly dictate sexual behavior; and there is a great deal of conscious regulation of mating behavior (Wunsch, 2017). Due to these differences between humans and non-primate mammalian species in which lordosis behavior has been observed, some research has suggested that lordosis may have been lost in the evolution of human sexual behavior (see Wunsch, 2017, for review).
However, there are both theoretical and empirical reasons to believe that lordosis behavior might exist in humans in a modified form that is not an obligate reflex, but rather a facultative behavior that can be selectively deployed. If lordosis was originally a cue to female sexual receptivity among our mammalian ancestors, then selection should have favored male psychological adaptations to experience mating attraction in response to this behavior. Such male responses would have created the selective conditions for the evolution of female adaptations to selectively engage in lordosis behavior to modulate male mating interest. Behavioral patterns observed in non-primate mammals provide evidence consistent with this proposed sequential evolution: (a) lordosis behavior among females as an indicator of sexual receptivity → (b) male adaptations to experience mating attraction in response to this behavior → (c) facultative female adaptations to modulate male mating interest by selectively engaging in lordosis. Indeed, in some non-primate mammals, lordosis behavior both is a cue to receptivity (i.e., a response to a male’s attempt to initiate mating behavior) and can be used facultatively as a signal of proceptivity (i.e., a behavior initiated by a female to evoke male interest, Beach et al., 1976). For example, in some rodents, when females are attracted to a male who has not approached them, they reposition themselves in closer proximity to the male and then engage in lordosis behavior (Beach et al., 1976). Lordosis behavior has also been observed as a proceptive signal in primates (e.g., macaques, Hanby, 1976). The existence of lordosis behavior as a proceptive signal among these primates is particularly important, because, like humans, they exhibit a relative attenuation of the olfactory circuits and a reduced association between hormones and sexual activity (Wunsch, 2017). Collectively, these findings suggest that although lordosis might have initially evolved as a hormonally governed reflex, subsequent selection pressures may have shaped female adaptations to selectively use it as a proceptive signal. If so, this suggests the hypothesis that selection shaped male psychological mechanisms to attend to this behavior. We refer to this as the Lordosis Detection Hypothesis.
Lumbar Curvature
The lordosis detection hypothesis articulates one reason that selection should have favored male psychological adaptations to attend to women’s lumbar region: to detect the behavioral cue of lordosis. However, women’s lumbar region may also contain fitness-relevant morphological cues.
Wedging in women’s lumbar vertebrae is critical for solving the adaptive problem of a forward-shifting center-of-mass during pregnancy. If ancestral women lacked the vertebral wedging necessary to shift their gravid center-of-mass back over the hips, they would have experienced as much as an 800% increase in hip torque during pregnancy (Whitcome et al., 2007). This torque on the hips subjects a woman’s lower back musculature (e.g., erector spinae) to sustained contraction, which would have resulted in muscular fatigue, increased her susceptibility to debilitating back injury, and significantly impaired her ability to forage (see Whitcome et al., 2007). This would have risked malnutrition for the woman, her developing fetus, her other dependent offspring, and her mate. By contrast, ancestral women with sufficient vertebral wedging to shift the gravid center-of-mass back over the hips would have been significantly less likely to incur these fitness costs (Whitcome et al., 2007).
These fitness differences between women as a function of their vertebral wedging would have created selection pressures for the evolution of male psychological adaptations to attend to cues to this wedging. Men who preferentially mated with women capable of re-centering the gravid center-of-mass would have derived several fitness benefits. This would have included having a mate who was less susceptible to spinal injury during pregnancy, could forage more effectively and longer into pregnancy, and could sustain multiple pregnancies with reduced likelihood of injury. This means that a woman’s lumbar vertebral wedging would have affected her fitness, the fitness of her offspring, and the fitness of her mate. These direct and indirect effects on her mate’s fitness would have created selection pressures for the evolution of a male mate preference for, and attraction to, specific angles of lumbar curvature—clinically operationalized as the angle formed between the buttocks and the thoracic spine while in a neutral posture—because this is an externally observable, reliable cue to wedging in the lumbar vertebrae (e.g., see George et al., 2003).
Selection should have shaped a male preference for a “Goldilocks” or “sweet spot” degree of lumbar curvature. Both insufficient vertebral wedging and excessive vertebral wedging would have posed fitness problems. Whereas insufficient vertebral wedging (hypolordosis) would have been associated with the inability to shift the gravid center-of-mass back over the hips, excessive vertebral wedging (hyperlordosis) would have increased shearing forces on the spine, which can lead to debilitating injury such as herniated intervertebral disks (see Whitcome et al., 2007; White & Punjabi, 1990). Selection therefore should have favored a male preference for an angle of lumbar curvature that minimizes the net fitness threats posed by both hypolordosis and hyperlordosis. This intermediate angle would cue the ability to shift the gravid center-of-mass back over the hips without excessive spinal shearing, thereby avoiding the harmful outcomes associated with both hypo- and hyperlordosis. This leads to the hypothesis of a male preference for, and attraction to, an angle of lumbar curvature that is maximally distant from the countervailing fitness threats of hypolordosis and hyperlordosis, 2 which medical orthopedic literature indicates is approximately 45.5° (see Fernand & Fox, 1985). We refer to this hypothesis—that men have a mate preference for an angle of lumbar curvature of approximately 45.5°—as the Fetal Load Hypothesis (see Lewis et al., 2015; Lewis, Russell, et al., 2017).
Discriminating Between the Lordosis Detection and Fetal Load Hypotheses
The Lordosis Detection Hypothesis and Fetal Load Hypothesis might seem to be in competition with one another. Each articulates a rationale for why selection should have shaped psychological mechanisms in men’s minds to attend to women’s lumbar region. However, there is a crucial distinction: between behavioral cues and morphological cues. This distinction reveals that these two hypotheses are not in competition with one another but rather are complementary; they propose distinct, non-mutually exclusive features in male mating cognition.
The Lordosis Detection Hypothesis posits a mate preference for a dynamic, movement-based cue (back-arching behavior), whereas the Fetal Load Hypothesis proposes a preference for a morphological cue (a woman’s lumbar curvature while in a neutral posture). Both hypotheses may be correct. If the Lordosis Detection Hypothesis is correct, then we should expect men to be attuned to the movement of increased back arching. And if the Fetal Load Hypothesis is correct, then we should expect men to be attracted to specific angles of lumbar curvature when women are in a neutral posture. Together, the Lordosis Detection Hypothesis and Fetal Load Hypothesis suggest that men’s mating cognition might attend to both lordosis behavior and lumbar curvature.
Another way in which the hypotheses can be differentiated is by the emphasis they place on mating context. The Lordosis Detection Hypothesis proposes that mechanisms for responding to lordosis behavior should be sensitive to mating context: Because it is more pressing to identify cues to proceptivity in short-term mating contexts than long-term contexts, the Lordosis Detection Hypothesis suggests that men’s attraction to back-arching behavior should be stronger in short-term than long-term contexts. By contrast, the Fetal Load Hypothesis does not predict context-dependent effects. To the extent that a central evolutionary function of both uncommitted and committed mating was to produce offspring, we should expect men to be attracted to cues to a woman’s ability to successfully gestate in both short-term and long-term mating contexts.
Study 1: Testing the Fetal Load and Lordosis Detection Hypotheses
To test the Lordosis Detection Hypothesis and the Fetal Load Hypothesis, we used human modeling software to generate animated female characters that varied on two dimensions: their (a) neutral lumbar curvature and (b) degrees of back arching (i.e., lordosis behavior). This enabled us to concurrently test for independent effects of these two distinct cues, and thereby test the distinct predictions generated from the Fetal Load Hypothesis and Lordosis Detection Hypotheses.
Method
Ethics statement
This study was approved by the Charles University Human Research Ethics Committee (Approval number: 2018/008).
Participants
One hundred two heterosexual male participants (Mage = 28.36 years, SDage = 11.43 years, age range = 18–68 years) completed the study. Participants were from Australia (62%), the United States (16%), the United Kingdom (9%), Canada (4%), and India (2%), as well as New Zealand, South Africa, Zimbabwe, China, Malaysia, UAE, Mexico, and Switzerland (all 1%). Participants were recruited through the Social Psychology Network, the subject pool at Murdoch University, personal contact from the researchers, on-campus advertising at Murdoch University, and snowball sampling. Student participants recruited via the Murdoch University subject pool received partial course credit for their participation.
Materials and procedure
As part of a larger study on attraction and mating, participants viewed the digital animated characters and completed the measures described below via an online survey hosted on the Qualtrics survey software platform.
Stimuli
We generated 17 animated virtual characters using a professional software tool for creating models of human figures and rendering animated digital content. The animated characters were displayed to participants in side profile and varied on two dimensions: their angle of neutral lumbar curvature (LC) and the degree of lower back arching that they exhibited in the animation (arching) (Figure 1; study stimuli publicly available at https://osf.io/sfcv6/?view_only=edb1bba510df41d69cbed69a0b3fa964). So that the stimuli were representative of the naturally occurring distributions of these variables, the characters ranged in neutral lumbar curvature from 29° to 66° (see Fernand & Fox, 1985) and in arching of the lower back from 8° to 28° (see Whitcome et al., 2007) (see Table S1 in the Supplemental Material for full details).
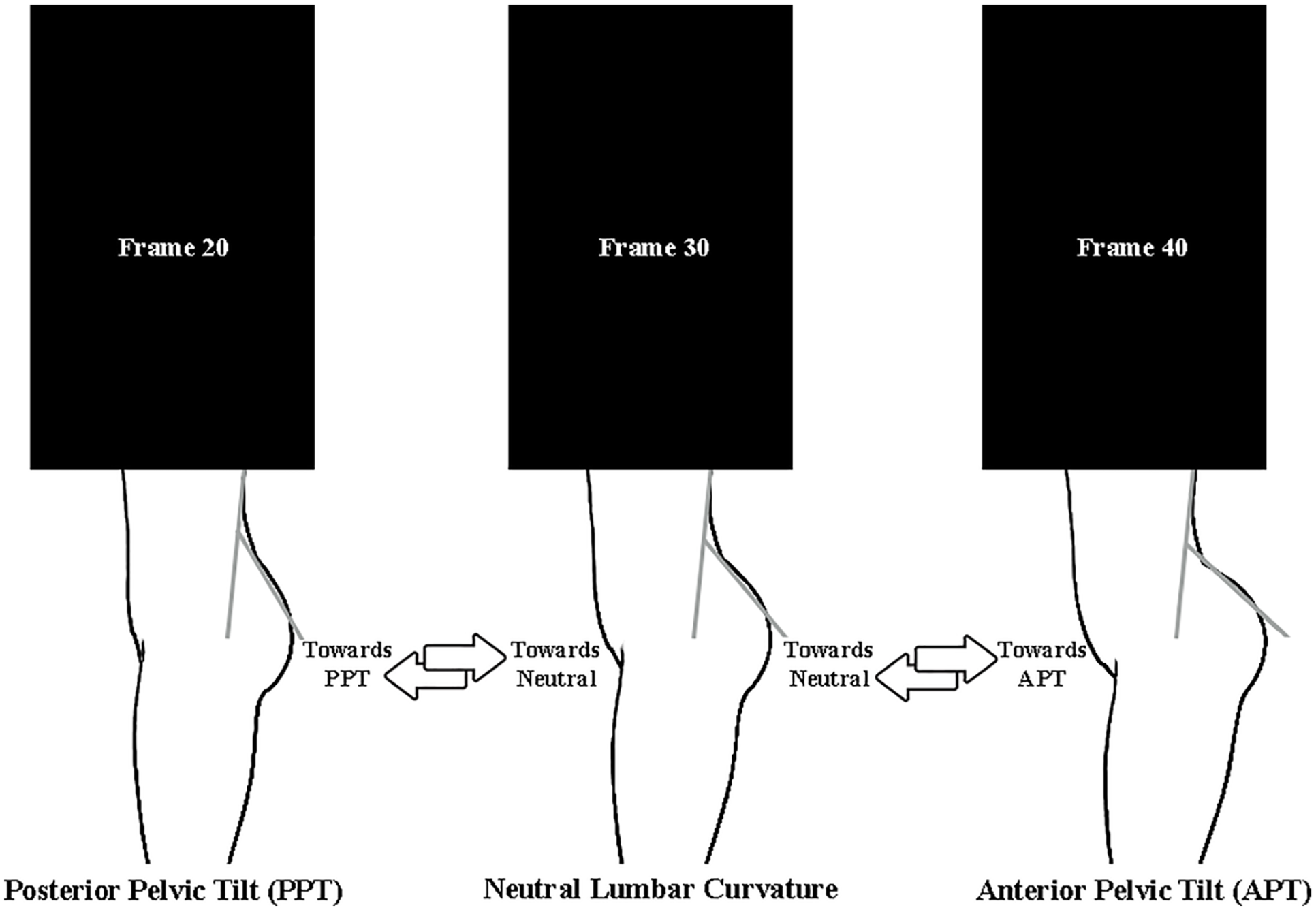
Static contour depiction of a dynamic study stimulus. Each stimulus was a video of an animated character moving between posterior and anterior pelvic tilt (i.e., engaging in back-arching behavior), passing through their neutral lumbar curvature. Some stimuli shared the exact same range of back-arching behavior (i.e., the range between posterior and anterior pelvic tilt) but differed in their neutral lumbar curvature; these stimuli enabled us to isolate the effect of lumbar curvature on perceived attractiveness. Other stimuli had identical angles of neutral lumbar curvature but differed in their back-arching behavior; these stimuli enabled us to isolate the effect of back-arching behavior on perceived attractiveness. (CC-By-Attribution 4.0 International).
Attractiveness ratings
Each participant viewed and rated all characters. The characters were presented one by one, in random order, with order randomized anew for each participant. The participant was asked to rate each depicted female character’s attractiveness (a) as a short-term mate and (b) as a long-term mate on a response scale ranging from 0 (extremely unattractive) to 10 (extremely attractive).
Results
Because each participant rated the attractiveness of all stimuli across both mating contexts, we fit study data to linear mixed-effect models that nested attractiveness ratings within each participant and incorporated a random intercept for each participant; this controlled for between-participant differences in baseline perceptions of attractiveness and enabled us to more powerfully isolate the effects of lumbar curvature and lordosis behavior on attractiveness. All analyses were conducted using the Statsmodels package (version 0.11.0; Seabold & Perktold, 2010) in Python (version 3.7.6; Van Rossum & Drake, 2009). Study data and analysis code are publicly available at https://osf.io/sfcv6/?view_only=edb1bba510df41d69cbed69a0b3fa964.
Distinct preferences for lumbar curvature and back arching?
To tease apart the independent effects of neutral lumbar curvature and dynamic back arching and thereby disentangle the Fetal Load Hypothesis and the Lordosis Detection Hypothesis, we fit all study data to a linear mixed-effect model in which lumbar curvature 3 and back arching were simultaneously entered as fixed effect predictors of attractiveness ratings. We also incorporated the interactions (a) between mating context and lumbar curvature and (b) between mating context and back arching to test whether any effects of lumbar curvature or back arching were dependent on mating context.
If the Fetal Load Hypothesis is correct, then men should be most attracted to an angle of lumbar curvature that reflects the proposed biomechanical optimum for successful gestation: men’s perceptions of women’s attractiveness should increase as women’s lumbar curvature approaches 45.5° (Prediction 1). Moreover, the Fetal Load Hypothesis suggests that this relationship between lumbar curvature and attractiveness should not be moderated by mating context (Prediction 2).
If the Lordosis Detection Hypothesis is correct, then men should be attracted to increased back arching (Prediction 3), but this effect should be more pronounced in short-term than in long-term mating contexts (Prediction 4).
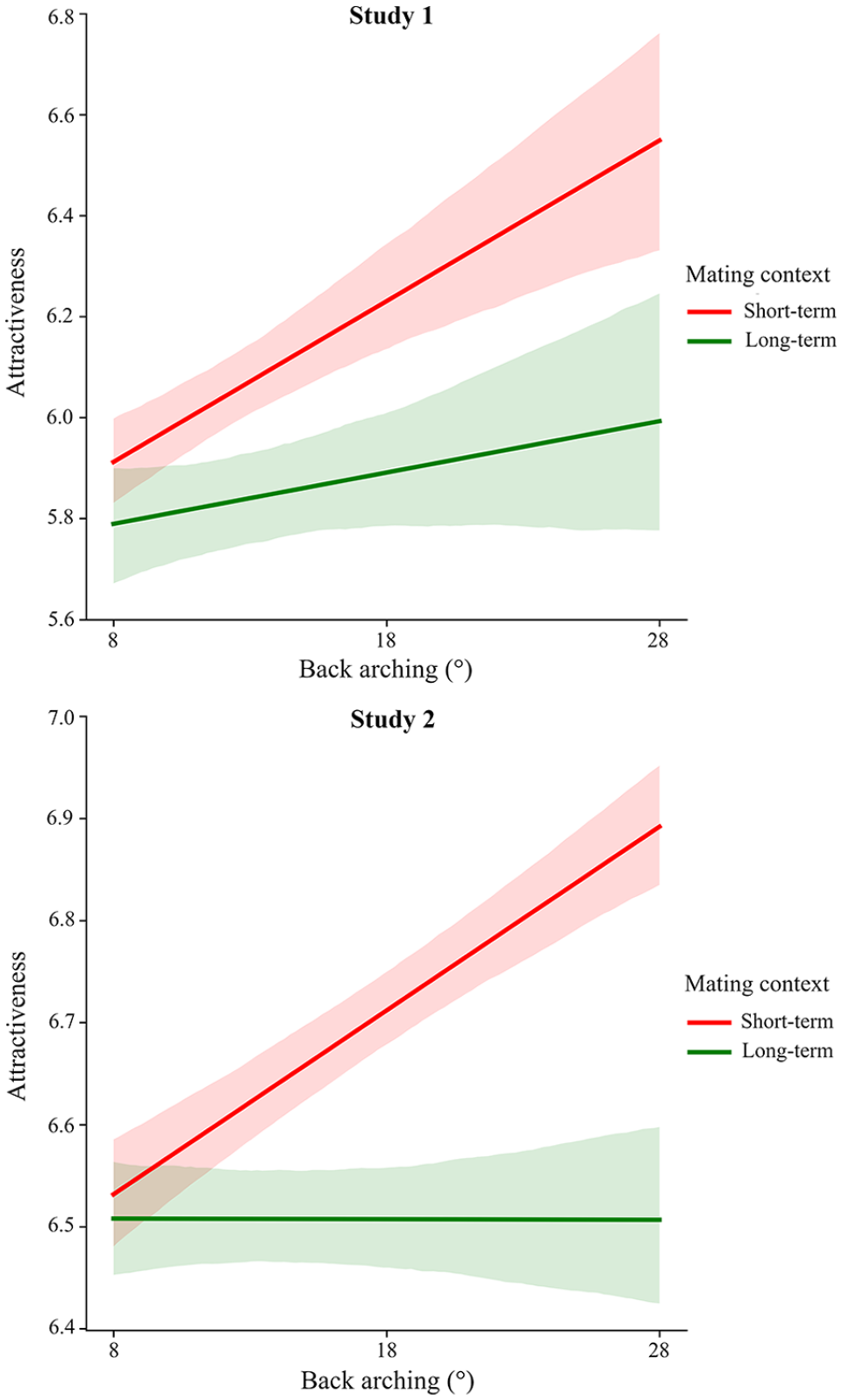
Results aligned precisely with all four predictions. As predicted by the Fetal Load Hypothesis, men’s attraction increased as the female characters’ lumbar curvature approached the proposed biomechanical optimum, b = −.051, SE = .007, p < .001, 95% confidence interval [CI] [−.065, −.037], β = −.14, and this effect did not vary across mating contexts: there was no interaction between lumbar curvature and mating context, b = −.011, SE = .010, p = .278, 95% CI [−.030, .009], β = −.02 (Figure 2, top panel).

The relationship between lumbar curvature and attractiveness.
By contrast—and in precise alignment with the Lordosis Detection Hypothesis—the effect of back arching on attractiveness was context-dependent; back arching interacted with mating context to predict perceptions of attractiveness, b = −.020, SE = .007, p = .006, 95% CI [−.035, −.006], β = −.04. We therefore conducted subsequent analyses to identify the distinct effects of back arching in short-term and long-term contexts. The Lordosis Detection Hypothesis suggested that back arching should have a positive effect in short-term contexts and that this effect should be stronger than in long-term contexts.
This is exactly what we observed. Back arching positively influenced perceptions of attractiveness in short-term mating contexts, b = .038, SE = .005, p < .001, 95% CI [.028, .049], β = .14. In long-term mating, back arching was also positively associated with attractiveness, b = .018, SE = .005, p < .001, 95% CI [.008, .028], β = .07, but, as indicated by the significant interaction between mating context and back arching, the effect of back arching was stronger in context of short-term mating (Figure 3, top panel).
The mating context-dependent relationship between lordosis behavior and attractiveness.
Study 2: Replication and Extension
Given the reproducibility crisis in psychology (Open Science Collaboration, 2015) and the fact that Study 1 is the first to report the above findings, we conducted a second study with an independent sample to replicate these findings. In addition, we incorporated individuals’ sociosexual orientation into study analyses. This enabled us to determine whether an orientation toward casual sexual relations moderated the observed effects.
Method
Ethics statement
The data for the replication study were collected under the same approved protocol as Study 1 and under protocol 21/09 approved by the Institutional Review Board of Charles University.
Participants
Two hundred thirty-one heterosexual male participants (Mage = 30.21 years, SDage = 9.89 years, age range = 18–72 years) completed the study. Participants were from the United States (45%), India (21%), Australia (17%), Czech Republic (7%), Brazil (2%), and the United Kingdom (2%), as well as Vietnam, Sweden, China, Georgia, Zimbabwe, Kenya, Philippines, Japan, Italy, Poland, and Thailand (each <1%). Participants were recruited through the Social Psychology Network, MTurk, Facebook, the subject pool at Murdoch University, and a database of individuals who agreed to be contacted to participate in research studies. Student participants recruited via the Murdoch University subject pool received partial course credit for their participation.
Materials and procedure
Stimuli
The same digital animated characters used in Study 1 were used in Study 2.
Attractiveness ratings
As in Study 1, each participant viewed all characters one by one, in random order, and rated each character’s attractiveness as a short-term mate and as a long-term mate on a scale from 0 (extremely unattractive) to 10 (extremely attractive).
Sociosexual orientation
We assessed participants’ orientation toward casual sexual relations using the Sociosexual Orientation Inventory–Revised (SOI-R, Penke & Asendorpf, 2008), which measures a person’s history of engaging in, attitudes about, and desire for uncommitted sex. The SOI-R consists of nine items such as “With how many different partners have you had sexual intercourse on one and only one occasion?,” “Sex without love is OK.,” and “How often do you experience sexual arousal when you are in contact with someone you are not in a committed romantic relationship with?” All items are scored on a nine-point scale and composited to form a single global SOI-R score. Higher scores correspond to a more unrestricted orientation (i.e., orientation toward casual sex).
Results
Replication analyses
First, to test the Study 1 findings for replication, we fit the Study 2 data to the exact same statistical models as those conducted in Study 1. Without exception, the four key findings replicated. In support of Prediction 1, men’s perceptions of the female characters’ attractiveness increased as the characters’ lumbar curvature approached the proposed biomechanical optimum, b = −.018, SE = .004, p < .001, 95% CI [−.026, −.009], β = −.05, and, in support of Prediction 2, this effect did not depend on mating context: There was no interaction between lumbar curvature and mating context in predicting perceptions of attractiveness, b = −.009, SE = .006, p = .153, 95% CI [−.022, .003], β = −.01 (Figure 2, bottom panel). These findings replicated the key results in support of the Fetal Load Hypothesis.
In contrast to the context-independent effects of lumbar curvature predicted by the Fetal Load Hypothesis, the Lordosis Detection Hypothesis predicts context-dependent effects: It predicts that back arching will positively influence perceptions of attractiveness more in short-term than long-term mating contexts. Consistent with this hypothesis, the effect of back arching on men’s perceptions of the female characters’ attractiveness differed between short-term and long-term mating contexts, as indicated by the significant interaction between mating context and back arching in predicting perceptions of attractiveness, b = −.017, SE = .005, p < .001, 95% CI [−.026, −.008], β = −.03. We therefore conducted subsequent analyses to examine the effect of back arching on perceptions of attractiveness in short-term and long-term contexts separately. In alignment with the Lordosis Detection Hypothesis, and as in Study 1, back arching positively influenced men’s perceptions of the female characters’ attractiveness as a short-term mate, b = .020, SE = .003, p < .001, 95% CI [.014, .026], β = .08. By contrast, back arching did not positively influence men’s perceptions of the female characters’ attractiveness as a long-term mate, b = .004, SE = .003, p = .246, 95% CI [−.003, .010], β = .01 (Figure 3, bottom panel).
Extension: Men’s sociosexual orientation
To test whether men’s sociosexual orientation (i.e., orientation toward uncommitted sexual relations) moderated the effects of lumbar curvature and back arching, we conducted additional analyses that incorporated men’s SOI-R scores. Specifically, we ran the same model as in Study 1 and the Study 2 replication analyses, but added two interactions: (a) between back arching, mating context, and men’s SOI-R scores; and (b) between lumbar curvature, men’s SOI-R scores, and mating context. The first interaction term enabled us to determine whether the context-dependent effect of back arching obtained among both sociosexually restricted and unrestricted men. The second interaction term enabled us to determine whether the effect of lumbar curvature varied as a function of men’s sociosexual orientation, in interaction with mating context.
We did not have a priori predictions about these interactions. This is because one can reason about the possible interactions between mating context and sociosexual orientation in several different ways. One body of literature emphasizes that psychological adaptations are generally species- or sex-typical (e.g., see Tooby & Cosmides, 1990). This “universal” perspective would seem to suggest that, given the appropriate contextual inputs, most men could engage in either committed or uncommitted mating (e.g., Lewis et al., 2015; see also Al-Shawaf et al., 2018). On this view, we might not expect the effects of lumbar curvature and back arching to vary as a function of men’s SOI-R scores; we might observe effects of context but not sociosexual orientation. A different perspective emphasizes sociosexual orientation as a stable individual difference variable (see Penke & Asendorpf, 2008; Simpson & Gangestad, 1991; see also Al-Shawaf, Lewis, Alley, & Buss, 2015; Al-Shawaf, Lewis, & Buss, 2015; Lewis, Al-Shawaf, et al., 2012). On this view, we might expect restricted and unrestricted men to respond differently to the cues of back arching and lumbar curvature. Currently, there are insufficient empirical data to adjudicate between these distinct views. 4 Consequently, we considered analyses involving sociosexual orientation to be exploratory.
We used a backward stepwise approach (removal criterion: p > .05) in which we began with a model incorporating the two, three-way interactions described above, along with all lower-order constituent terms. This resulted in a sequence of analyses that examined both three-way interactions, all two-way interactions, and the fixed effect (i.e., the “main effect”) of sociosexual orientation. All analysis code is publicly available at https://osf.io/sfcv6/?view_only=edb1bba510df41d69cbed69a0b3fa964. Neither of the three-way interactions were significant (both ps > .39). This indicated, for example, that the context-dependent effect of back arching did not depend on men’s sociosexual orientation. The fixed effect of lumbar curvature and the two-way interaction between back arching and mating context remained statistically significant in the final model; that is, the context-dependent effect of back arching and the context-independent effect of lumbar curvature were not moderated by men’s SOI-R scores and were robust to their inclusion in study analyses.
The only significant terms involving men’s sociosexual orientation that emerged in the final model were the fixed (i.e., “main”) effect of men’s SOI-R scores, b = .528, SE = .063, p < .001, 95% CI [.406, .651], β = .38—which indicated that unrestricted men perceived the female characters to be more attractive, on average, than did restricted men—and the interaction between men’s SOI-R scores and back arching, b = −.003, SE = .001, p = .023, 95% CI [−.006, .000], β = −.02. To examine the interaction between men’s SOI-R scores and back arching, we conducted simple slopes analyses. The interaction reflected that, averaged across mating contexts, the coefficient for back arching was larger for restricted men (M − 1SDSOI: b = .017, SE = .003, p < .001) than for unrestricted men (M + 1SDSOI: b = .006, SE = .003, p = .07). Ultimately, however, the effects observed in Study 1 and in the Study 2 replication analyses emerged among both restricted and unrestricted men: restricted and unrestricted men alike were attracted to back arching in short-term mating contexts (M + 1SDSOI: b = .013, SE = .004, p < .001; M − 1SDSOI: b = .027, SE = .004, p < .001) but not long-term mating contexts (M + 1SDSOI: b = .000, SE = .004, p = .912; M − 1SDSOI: b = .007, SE = .005, p = .136).
General Discussion
The current studies report and replicate several previously unknown findings. First, these studies are the first to demonstrate independent effects of the dynamic cue of back arching and the morphological cue of lumbar curvature. These findings support both the Fetal Load Hypothesis and the Lordosis Detection Hypothesis.
Second, these studies are the first to test whether mating context moderates the effects of lumbar curvature and back arching on perceptions of attractiveness. Consistent with the Fetal Load Hypothesis, we found that the effects of lumbar curvature were independent of mating context: men were attracted to the proposed biomechanically optimal angle of lumbar curvature regardless of mating context. By contrast, and consistent with the Lordosis Detection Hypothesis, the effects of back-arching behavior depended on mating context: Men were more attracted to back arching in short-term than long-term mating contexts. The two studies found nearly identical results, highlighting their replicability, which we discuss in greater detail below.
Mating Context and Sociosexual Orientation
Men’s sociosexual orientation did not moderate any of the key effects observed in Study 1 or the Study 2 replication analyses. For example, restricted and unrestricted men alike were attracted to back arching in short-term but not long-term mating contexts. This may be most consistent with the “universal” perspective described above: Men may be able to shift into either a short-term or long-term mating mindset in alignment with the present mating context, regardless of their sociosexual orientation. The fact that sociosexual orientation exerted relatively little influence when mating context was incorporated into study design raises several important questions for future research.
First, if any given individual is capable of engaging in either committed or uncommitted mating in alignment with the present mating context (e.g., see Lewis et al., 2015), then by what processes do individual differences in sociosexuality predict important mating-related outcomes? We suspect that a key to the answer lies in differences in the selection, evocation, and manipulation (Buss, 1987) of mating environments by restricted and unrestricted individuals. Unrestricted individuals may preferentially select social contexts that facilitate short-term mating. They may also evoke different responses from others in the social environment (e.g., see Plomin et al., 1977; Scarr & McCartney, 1983). For example, because sociosexuality is positively associated with physical attractiveness (e.g., Al-Shawaf, Lewis, & Buss, 2015) and people value physical attractiveness in a potential mate (e.g., see Buss, 1989)—especially when seeking a short-term mate (e.g., Li & Kenrick, 2006)—sociosexually unrestricted individuals, more so than restricted individuals, may evoke an environment in which others are interested in them as short-term partners. Finally, unrestricted individuals may be more likely than restricted individuals to manipulate the mating environment so that it opens up opportunities for uncommitted sexual relations (e.g., see Lewis, Easton, et al., 2012).
In the current research, we had all men evaluate the study stimuli in both short-term and long-term mating contexts. This design inherently controls for any processes by which restricted and unrestricted men might find themselves in different mating contexts. This design—namely, making the mating context explicit—may reduce variability in participants’ interpretation of context, which has several advantages. For example, if research instructs participants to consider their “ideal mate” without specifying the mating context (i.e., short-term vs. long-term), participants may think about their preferred context: Restricted individuals may consider their ideal long-term mate, whereas unrestricted individuals may consider their ideal short-term mate. If so, it would not be clear whether any differences between restricted and unrestricted participants’ preferences should be attributed to differences in sociosexual orientation or differences in mating context.
A related advantage of specifying mating context is that it enables clearer tests of the overarching hypothesis of universal psychological adaptations (e.g., Tooby & Cosmides, 1990). A core hypothesized feature of universal evolved psychological mechanisms is that they are sensitive to contextual inputs and change their outputs (e.g., mate preferences) as a function of context (DeKay & Buss, 1992; see also Al-Shawaf et al., 2019, 2021; Goetz et al., 2012; Lewis, 2015; Lewis, Al-Shawaf, et al., 2017; Lewis & Buss, 2022; Lewis et al., 2021; Lukaszewski et al., 2020). On this view, we should expect to observe variability in people’s preferences if they interpret ambiguous contextual information differently. Conversely, when mating context is not left open to interpretation (e.g., it is not so ambiguous that restricted and unrestricted participants can interpret it in line with their preferred mating context), we might expect to observe more similar psychological responses between restricted and unrestricted individuals. This appears to be consistent with what we observed in the current study.
The current finding—that sociosexual orientation exerted relatively little influence when mating context was explicitly incorporated into study design—together with the fact that sociosexual orientation does predict mating-related outcomes in the real world (e.g., see Simpson & Gangestad, 1991), suggests to us that intrinsic links between sociosexuality and social processes, such as the differential selection, evocation, and manipulation of mating environments by restricted and unrestricted individuals, may be key to understanding when individual differences in sociosexuality are (not) likely to predict important mating outcomes.
We also note that it may be prudent to treat the results involving sociosexual orientation more tentatively than the core results (i.e., the context-independent effect of lumbar curvature and the context-dependent effect of back arching). With respect to the effects of lumbar curvature and back arching, Study 1 effectively served as a preregistration for Study 2; in the Study 2 replication analyses, we ran the exact same statistical models as in Study 1, and these confirmatory analyses in Study 2 reproduced, without exception, all key findings involving lumbar curvature and back arching. However, because Study 2 was the first involving analyses that incorporated men’s sociosexual orientation, Study 1 did not serve the same preregistration function for these analyses, and we cannot directly address the reproducibility of the results involving men’s sociosexual orientation.
Under any condition, more research is needed in this area, as there is limited work concurrently investigating the effects of mating context and sociosexual orientation. Research on mate preferences has tended to focus on either the effects of mating context or sociosexual orientation, but not both, and not the interaction between them. Ultimately, more empirical data are needed to better resolve the relationship between mating context and sociosexual orientation, as well as their interaction.
Display of Gluteofemoral Fat Reserves: An Alternative Explanation?
In the current studies, we generated a priori predictions based on the Lordosis Detection Hypothesis and found evidence consistent with those predictions. The broad observation of male attraction to female back-arching behavior, however, is also consistent with another hypothesis: The Gluteofemoral Fat Display Hypothesis. Here, we describe this hypothesis—but also illustrate why it cannot account for study findings.
A possible alternative function of back-arching behavior could be to display gluteofemoral fat reserves; evidence suggests that men are attracted to such reserves because they are important to infant brain development (see Lassek & Gaulin, 2019). However, this hypothesis does not generate a key prediction about the context-dependent effects of back-arching behavior that (1) the Lordosis Detection Hypothesis does generate and (2) was supported in both the original study and the replication study. Specifically, a central prediction of the Lordosis Detection Hypothesis is that back arching will be more attractive in short-term mating contexts than in long-term mating contexts. The Gluteofemoral Fat Display Hypothesis does not generate this prediction. Rather, the Gluteofemoral Fat Display Hypothesis appears to either predict a context-independent effect—gluteofemoral fat reserves should be perceived as attractive in both short-term and long-term mates—or a context-dependent effect in the opposite direction: Because gluteofemoral fat reserves are a cue to parity and future reproductive potential—which are more important in the context of long-term mating—the Gluteofemoral Fat Display Hypothesis might predict that displaying these reserves should increase women’s attractiveness more in the context of long-term mating. Either way, the Gluteofemoral Fat Display Hypothesis does not generate the prediction that back arching should increase attraction more in short-term than long-term contexts.
This means that tests of the context-dependent effect of lordosis behavior are a clear adjudicator between the Lordosis Detection Hypothesis and the Gluteofemoral Fat Display Hypothesis. In both the original study and the replication study, we found that back-arching behavior increased women’s attractiveness more in the context of short-term than long-term mating. These findings are precisely those expected under the Lordosis Detection Hypothesis, and, at a minimum, cannot be readily accounted for by the Gluteofemoral Fat Display Hypothesis (and may directly contradict it).
The Operationalization of the “Optimum” Lumbar Curvature
We operationalized the Fetal Load Hypothesis by predicting that men will be most attracted to a lumbar curvature angle of 45.5°. This operationalization is likely to be imperfect (e.g., it rests on the assumption that the fitness costs of hypolordosis and hyperlordosis are approximately equal), but it has several key advantages relative to alternatives.
First, we arrived at this operationalization a priori rather than post hoc. After we generated the Fetal Load Hypothesis, we operationalized the optimum by using a two-step approach of first identifying, in the medical orthopedic literature, the values associated with the medical problems of hypolordosis and hyperlordosis, and then computing the value that is maximally distant from those countervailing adaptive problems. This process highlights another advantage of this operationalization: It is based on clearly outlined adaptive problems—We operationalized the optimum lumbar curvature as the point maximally distant from the medical problems of hypolordosis and hyperlordosis.
An alternative way to operationalize the Fetal Load Hypothesis would be to simply predict a quadratic (i.e., inverted U) relationship between lumbar curvature and attractiveness. However, there are two problems with this. First, it would be easy—too easy—to find support for this general prediction: the observation of any quadratic relationship would support it. This would not be scientifically prudent, as it could lead to inferring support for the Fetal Load Hypothesis if the inflection point of the curve was at 39°, 57°, or any other value. That is too many degrees of inferential freedom and represents a much less rigorous test of the hypothesis than the approach we used of operationalizing the hypothesized optimum in advance and testing men’s preferences against this pre-specified value.
A second problem with this generic quadratic approach is that it has an element of circularity. The Fetal Load Hypothesis generates that prediction that the (a) values that men prefer will align with (b) a fitness optimum. To test that hypothesis, the values that men prefer cannot feed into the operationalization of the optimum; the optimum must be operationalized independently of any known preferred values. Otherwise, the logic becomes circular: If the hypothesis is that men’s preferred values will correspond to the optimum, and then the operationalization of the optimum is based on the values preferred by men, the argument is little more than a tautology.
In sum, despite whatever limitations the 45.5° operationalization might have, it has several key advantages. The quadratic approach is potentially circular in logic and offers too many researcher degrees of freedom with respect to inferring support for the hypothesis. By contrast, the 45.5° operationalization reflects an a priori derivation of optimum and is based on values identified in advance as being linked to specific adaptive problems.
Dynamic Stimuli
The current studies point to the importance of moving beyond static stimuli in evolutionary research on attractiveness. The exclusive use of static stimuli neglects the fact that selection could have shaped mate preferences for movements that convey important fitness-related information. Using dynamic stimuli enables researchers to (a) test for these movement-based preferences; (b) prevent the misattribution of preferences for dynamic cues as preferences for static cues, and vice versa; and (c) test the possibility that selection shaped preferences for both static and dynamic cues in the same feature or region of the phenotype. Without stimuli that are designed to test hypotheses about both classes of phenotypic cues, certain discoveries will remain inaccessible. In the studies reported here, we identified distinct predictions generated by the Fetal Load Hypothesis and the Lordosis Detection Hypothesis, designed stimuli to test these predictions, and discovered previously unknown features of mating cognition.
Limitations and Future Directions
Despite the contributions of the studies reported here, the current research represents only an initial foray into lordosis in humans. The current studies focused on psychological adaptations to attend to the back-arching movement, but do not directly demonstrate lordosis behavior. An important task for future work will be to more clearly establish the existence of human lordosis behavior.
For now, we note that there are convergent lines of evidence suggesting that lordosis behavior likely does exist in humans—or, at the very minimum, that the assumption that it does not exist should be carefully reconsidered. The first line of evidence comes from vernacular dance, a “form of nonverbal communication” (Pérez, 2016, p. 16). One such vernacular dance that has recently entered common vocabulary is twerking, which refers to a “a sexually provocative dance or dance move involving thrusting movements of the bottom and hips” (Oxford University Press, n.d.). Formally, the “thrusting movements” refer to the anterior-posterior tilting of the pelvis—a defining feature of lordosis behavior. Although “twerking” might be new as a household term, its etymology dates back more than 200 years and the behavior is far from new—it was documented across diverse cultures long before it became a social media phenomenon. Indeed, the lordotic movement “unite[s] a dizzying array” (Pérez, 2016, p. 22) of dances from myriad cultures across geographic regions, religious backgrounds, biological ecologies, linguistic groups, and cultural values, attitudes, morals, and rituals (see Figure S1 in the Supplemental Material for a cross-cultural sampling of dances from Africa, Asia, Europe, Oceania, and North and South America that incorporate the lordotic movement).
A second line of evidence that points toward the existence of human lordosis behavior comes from unpublished work by Senveli (2017). Senveli took two photographs of women—one while a female research assistant was in the room with the woman, and another after a professional male model entered. A comparison of the women’s photographs before and after the arrival of the male model indicated that, on average, women adjusted their posture to have a more acute angle between the thoracic spine and sacrum—that is, they engaged in back arching—when the model entered the room.
The evidence remains preliminary, but, taken together, men’s attraction to the lordotic movement observed in the current study, Senveli’s (2017) tentative experimental results, and the omnipresence of the lordotic movement in dances from distant and independent cultures (see Figure S1 in the Supplemental Material) suggest that, at a minimum, researchers should reconsider the assertion that lordosis behavior does not exist in humans.
We encourage future research to more firmly establish the existence of human lordosis behavior and to identify what cultural inputs during ontogeny influence the development of lordosis behavior and the meaning assigned to it, what cultural values influence perceptions and moral judgments of it, what norms regulate the contexts in which the lordotic movement is condoned or deemed taboo, and what shared and distinct meanings it has across cultures. The current manuscript has focused on one potential communicative function of human lordosis behavior, but a key goal of future research should be to identify other meanings, symbols, or signals (not necessarily related to proceptivity) that human lordosis behavior might be used to communicate.
Toward the goal of mapping the semantics of this behavior in different contexts and cultures, ethnographic accounts may be useful in demonstrating the existence of human lordosis behavior while simultaneously illuminating the diverse meanings that the behavior may have. For example, in some cultures, the lordotic movement may be embedded in specific forms of dance, and engaging in those dances may be a cultural ritual. Under such cultural conditions, the meaning of lordosis behavior could range from merely fulfilling one’s cultural duties without any romantic or sexual intent to actively signaling sexual interest. Brazil’s Candomblé religion illustrates this well. In Candomblé, the lordotic movement appears among the nonverbal courtship signals of the gods. The orixá (deity) Oyá de esteira, whose name refers to a bedroll that she carries with her for the purposes of opportunistic mating (Pérez, 2016), “performs the ‘plate-breaking’ dance: hands on hips, buttocks prominently extended” (Gleason, 1992, p. 290). In Candomblé, dance is a religious obligation (Pérez, 2016). Consequently, when a Candomblé practitioner performs the plate-breaking dance, which captures the “libidinal heat” of this “erotically charged” deity through the lordotic movement (Cartwright, 2013, p. 165), this movement may have multiple possible meanings and may induce culturally specific responses. This is just one example of the many ways in which the intent of the individual engaging in the lordotic movement as well as the perceived meaning of the behavior may vary as a function of cultural context and other situational variables.
An important avenue for future research will be to identify the distinct meanings of lordosis behavior across cultures, between individuals within a given culture, and within individuals across different situations. Although some women in some contexts might use the lordotic movement to signal mating interest, it would be logically unsound to infer that the lordotic movement always signals mating interest. Drawing conclusions about all instances from some instances is always unwarranted, and the inference “If a woman is engaging in lordosis behavior, it means that she is sexually interested” would be entirely unjustified. In other words, and for the avoidance of doubt, the notion that the lordotic movement might be used by some women in some contexts as a signal of proceptivity in no way suggests that a given woman exhibiting back-arching behavior in a particular circumstance is inviting a sexual advance. We caution against such strong and unjustified inferences; there are myriad reasons unrelated to mating why women might engage in the lordotic movement. The current studies report data consistent with the hypothesis that signaling proceptivity could be one of the reasons that a woman might engage in the lordotic movement, but future work is needed to further test this hypothesis, to more firmly establish the existence of human lordosis behavior, and to identify the many distinct meanings and functions this behavior could have in humans.
Conclusion
The current studies provide evidence of several previously undocumented features of men’s mating psychology: Men’s minds (a) track both the morphological cue of women’s lumbar curvature and the dynamic cue of lordosis behavior, and (b) regulate mating attraction in response to these cues differentially as a function of mating context—in precise adaptive alignment with the context-dependent relevance of the cues. These findings, all predicted a priori on the basis of evolutionary reasoning, underscore the value of an evolutionary approach for making new predictions and discoveries about human mating psychology.
We hope that the current work also contributes to important directions for future research. In particular, we hope that the current studies inspire renewed focus on an understudied class of cues in the human phenotype: dynamic cues. Little work has been dedicated to charting these important components of the human phenotype or the psychological systems that attend to them. We hope that the current study makes a modest contribution toward the goal of mapping these cues and the cognitive systems that may have evolved to process them.
We also hope that the current studies motivate research investigating the interactions between humans’ universal evolved psychological mechanisms and the specific cultural inputs to which people are exposed during their development and lifespan. Ultimately, such an integrated approach will be necessary for understanding the meaning and perception of human behavior, including similarities and differences in these meanings and perceptions across people, contexts, and cultures.
Supplemental Material
sj-docx-1-psp-10.1177_01461672221115218 – Supplemental material for Lordosis in Humans
Supplemental material, sj-docx-1-psp-10.1177_01461672221115218 for Lordosis in Humans by Ayten Yesim Semchenko, Zeynep Senveli, Mitchell R. L. Forrest, Jonathon Flores, Vojtěch Fiala, Laith Al-Shawaf, David M. Buss and David M. G. Lewis in Personality and Social Psychology Bulletin
Footnotes
Authors’ Note
A.Y.S. contributed to data collection and manuscript preparation, and prepared, analyzed, and visualized all data. Z.S. contributed to project conceptualization and manuscript drafting. J.F. contributed to study design, generated the study materials, and assisted with manuscript preparation. D.M.G.L. elaborated the hypotheses, designed the studies, and wrote the manuscript. L.A-S., M.R.L.F, and D.M.B. contributed to manuscript revising and editing. V.F. contributed to data collection and manuscript reviewing.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported in part by grant 1742218 from Charles University Grant Agency (GAUK) to A. Y. Semchenko.
Supplemental Material
Supplemental material is available online with this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.