
Abstract
This study aimed to compare children’s performance on two mnemonic functions that engage the lateral prefrontal cortex. Brain imaging studies in adults have shown that the mid-ventrolateral prefrontal cortex is specifically involved in active controlled retrieval, and the mid-dorsolateral prefrontal cortex is specifically involved in monitoring mnemonic information (Petrides, 2005). Eighty-two children aged from 6 years, 8 months to 8 years, 7 months were tested. They showed equivalent success rates in active retrieval and monitoring with color and shape information. However, children were slower in monitoring than in active retrieval in color trials. The results demonstrate that the specialized contributions of the lateral prefrontal cortex emerge conjointly during childhood giving children multiple tools to exert an active control within memory.
Working memory is a framework of processes that involve the temporary storage and manipulation of information for complex cognitive abilities (Baddeley, Allen, & Hitch, 2011). These processes involve the concerted activity of a network of cortical regions, including the parietal cortex and the prefrontal cortex (Pessoa, Guttierrez, Bandettini, & Ungerleider, 2002). The most lateral part of the prefrontal cortex is recognized as essential for the executive processes in memory by exerting a top-down modulation on specific posterior association cortical areas (O’Reilly, 2010; Petrides, 1995, 1996, 2005; Tomita, Ohbayashi, Nakahara, Hasegawa, & Miyashita, 1999; Wilson, Gaffan, Browning, & Baxter, 2010). The lateral prefrontal cortex is not homogeneous and can be subdivided into a mid-ventrolateral prefrontal cortex (MVLFC) (areas 45 and 47/12) and a mid-dorsolateral prefrontal cortex (MDLFC) (dorsal areas 46, 9/46, 9) based on cytoarchitectonic characteristics and connectivity patterns (Petrides & Pandya, 1994). Anatomical, lesion, and brain imaging studies have shown that each of these regions makes a specific functional contribution to cognition and memory (Petrides, 2005).
According to the Petrides model (1995, 1996), the specific role of the MVLFC is to control first-order executive processes such as active selection, comparison, and judgment of stimuli held in short- and long-term memory. Many imaging studies have supported this model, showing that the MVLFC plays a crucial role in guiding the selection and retrieval of linguistic knowledge (Thompson-Schill, D’Esposito, Aguirre, & Farah, 1997; Wagner, Paré-Blagoev, Clark, & Poldrack, 2001). The MVLFC is specifically involved in memory tasks that require retrieval under challenging conditions (Badre, Poldrack, Pare-Blagoev, Insler, & Wagner, 2005; Haxby et al., 1996; Jonides et al., 1993). It has been demonstrated that the common challenge in these tasks is the fact that stimuli in memory are combined in multiple and equiprobable ways. Under challenging stimulus conditions, retrieval of specific information in memory cannot be guided by associative links, as in automatic retrieval, for example when the color yellow is automatically triggered by the word “banana.” Instead, it requires controlled processing to disambiguate stored content (Petrides, 2002, 2005), defined as active controlled retrieval (Cadoret & Petrides, 2007). This active retrieval will be asked, for example, if the location of a car has to be retrieved while it was randomly parked in a location among three possible locations in a vast parking lot. Functional MRI studies have repeatedly demonstrated that when adult participants perform active controlled retrieval, the MVLFC (and not other prefrontal regions) is specifically engaged to exert active control over visual and tactile memory information (Cadoret & Petrides, 2007; Cadoret, Pike, & Petrides, 2001; Kostopoulos, Albanese, & Petrides, 2007; Kostopoulos & Petrides, 2003). Moreover, a single-cell recording experiment in monkeys revealed that the firing rate of neurons in the MVLFC changed in relation to retrieval. Some of these neurons coded specifically for whether the decision to retrieve information from memory was the instructed one, and were called “controlled retrieval decision neurons” (Cadoret & Petrides, 2007).
In contrast, the MDLFC is an area that specifically monitors information held on-line, that is, it “keep[s] track of the relative status of multiple information in working memory, such as which one from an expected set has occurred” on the basis of task requirements or subject’s current place (Champod & Petrides, 2010; Chen et al., 2004; Diamond, Prevor, Callender, & Druin, 1997; Lezak, Howieson, Loring, Hannay, & Fischer, 2004; Petrides & Milner, 1982; Petrides, 1995). For example, if the absence of a person from a group of four has to be reported from a past event (who was absent?), monitoring will help to check in memory the presence or absence of each group member. The functional specificity of the MDLFC was first determined with lesion studies in monkeys. If the lesions were restricted to the MDLFC (i.e., dorsal area 46 and area 9), monkeys showed impairment on tasks such as self-ordered and externally-ordered tasks, which required monitoring the occurrence of stimuli from an expected set (Petrides, 1995). This specificity has been confirmed with positron emission tomography (PET) studies. When subjects performed self-ordered or externally-ordered tasks in which they had to monitor in working memory visual or verbal stimuli, specific activation patterns in the MDLFC were observed (Petrides, Alivisatos, Evans, & Meyer, 1993a; Petrides, Alivisatos, Meyer, & Evans, 1993b).
During development, maturation of the lateral prefrontal cortex is protracted to late adolescence or early adulthood (Chugani, Phelps, & Mazziotta, 1987; Uylings, Delalle, Petanjek, & Koendering, 2002; Yakovlev & Lecours, 1967), resulting in a prolonged development of working memory (Olson & Luciana, 2008). Some studies suggest that the MVLFC and the MDLFC mature at different rates, the MDLFC being slower (Gogtay et al., 2004; Ofen et al, 2007). For example, the MVLFC reaches maximum thickness at approximately 9.7 years, whereas the MDLFC peaks 1 year later, at approximately 10.5 years (Shaw et al., 2008). Using brain imaging, Crone, Wendelken, Donohue, van Leijenhorst, and Bunge (2006) examined activations associated with the storage and manipulation of visual information in working memory and observed in three age groups (8–12 years, 13–17 years, and adults) that the MVLFC was activated in relation to storage in working memory, whereas activation in the MDLFC was found only in 13–17-year-olds and adults when manipulation in working memory was required. When subjects’ performance was correlated with their activation profiles, children scored lower on manipulation than both adolescents and adults. Thus, the MDLFC was recruited later in development than the MVLFC, suggesting a slower maturation rate. A difference in maturation between the dorsal and the ventral parts of the lateral prefrontal cortex should also influence the emergence of the specialized functional contributions of these regions that are active retrieval and monitoring functions.
Previous studies with children (Dionne & Cadoret, 2013; Luciana & Nelson, 1998) have shown that performance on active controlled retrieval and on monitoring improves with age. For instance, a significant improvement in active controlled retrieval was observed in children aged from 6 to 8 years, where the older group performed similar to adults on shape and color retrieval (Dionne & Cadoret, 2013). On the other hand, it was demonstrated that children monitor an increasing number of items with age: from four locations at age 8 years to eight locations at age 15 years (Luciana & Nelson, 1998). However, these data were collected on separate samples using tasks that were not matched in terms of stimuli and number of trials to reduce the confounding effects of stimulus difficulty or cognitive fatigue. Therefore, to understand how the developing lateral prefrontal cortex contributes to memory development and how the specific memory functions of the MVLFC and the MDLFC are expressed during middle childhood, the objective of this study was to compare active controlled retrieval and monitoring performance in children. The comparison was focused on the period between 6 and 8 years old because previous data showed that active retrieval development was more important during this period (Dionne & Cadoret, 2013). Based on these results, the first hypothesis was that older children would perform better than younger children in active retrieval. The same visual stimuli made of specific colors and shapes were used in both functions. Color and shape were selected because they were easy to dissociate in active retrieval for young children. As shown previously (Dionne & Cadoret, 2013) it was predicted that performance on color would be higher than on shape active retrieval. Assuming that the MVLFC matures first, a third hypothesis was that children would show better active controlled retrieval performance than monitoring performance, with a higher increase with age in active controlled retrieval.
Method
Participants
Participants were 87 children aged from 6 years, 8 months to 8 years, 7 months (41 boys; 46 girls) recruited from a regular public primary school in a suburban area of Montreal (Canada). The sample was largely Caucasian and middle class to upper middle class based on socioeconomic school ranking. Of the children, 43 were in the first grade and 44 were in the second grade. Inclusion criteria included being in the desired age range, having normal or corrected-to-normal vision and hearing, and having no current or past history of neurological illness, psychological illness, mental deficiency, or learning difficulties. The study was approved by the ethics committee of the Université du Québec à Montréal and all children and their parents gave their informed consent for participation. Parents were invited to complete a short questionnaire about children’s age, laterality, and neurological status. Children received a small present for their participation.
Procedure
Children were tested individually at school in a separate and quiet room in accordance with their teachers. The experimental protocol included an active controlled retrieval task and a monitoring task presented to children as computer games. Children were asked to place their right thumb on the green button (YES) and their left thumb on the red button (NO) of the response pad and to keep them there until the end of each trial block. The experimenter verbally described each task to the children first, and the children then performed six trials with verbal guidelines and success/error feedback from the computer as a tutorial. Children were thus progressively trained toward real task conditions. Phase duration was progressively reduced until verbal support was no longer provided. If children failed on more than half the trials, they were asked to repeat the tutorial once.
Tasks
Each trial in the active controlled retrieval task was initiated by the display of a fixation point at the center of the computer screen (Figure 1a). The encoding phase began 1,000 ms later when one of four shapes in one of four colors was presented at the center of the screen for 1,000 ms. The encoded stimulus therefore had two aspects: shape and color. Importantly, combinations were presented with random but equal probability across trials to ensure that children could not establish a stable relationship between shapes and colors. The encoding phase was followed by a delay (1,000 ms), during which the screen was blank. A cue then appeared (for 500 ms), instructing children to retrieve a specific aspect of the information presented during the encoding phase (shape or color). A grey circle instructed them to retrieve the shape and a circle containing patches of color instructed them to retrieve the color. Therefore, retrieval was mainly visual. Children were not allowed to speak even if some subjects could demonstrate some verbal labelling. As soon as the cue appeared subjects were required to select by visual imagery the cued characteristic and keep it in mind. After a further delay (2,000 ms) in which the screen was blank, the test phase followed. A single stimulus was presented, again consisting of two aspects: a shape and a color. Children had to decide whether an aspect (shape or color) of the stimulus corresponded to the aspect (e.g. shape) of the previously encoded stimulus that the cue had instructed the child to retrieve. If the relevant aspect of the test stimulus (e.g. shape) matched the aspect of the encoded stimulus, the child had to press the YES key as quickly as possible (within 900 ms). If the relevant aspect of the test stimulus did not match, the child had to press the NO key. Thus, the correct response in the test phase depended entirely on the child’s ability to retrieve from memory the specific aspect of the stimulus as defined by the retrieval cue. Children performed 32 trials: 16 shape-retrieval trials and 16 color-retrieval trials, randomly intermixed. In half of the trials, the test stimulus was either identical in both characteristics (shape and color) to the encoded stimulus or different in both aspects. For instance, if shape A was presented in color X during the encoding phase, shape A in color X could be presented in the test phase (identical in both characteristics), or shape B in color Y might be presented (different in both characteristics). In the remaining half of the test trials, only one of the stimulus dimensions of the encoded event was the same as that of the encoded stimulus. In the example above, shape A might appear in color Y or shape B might appear in color X.
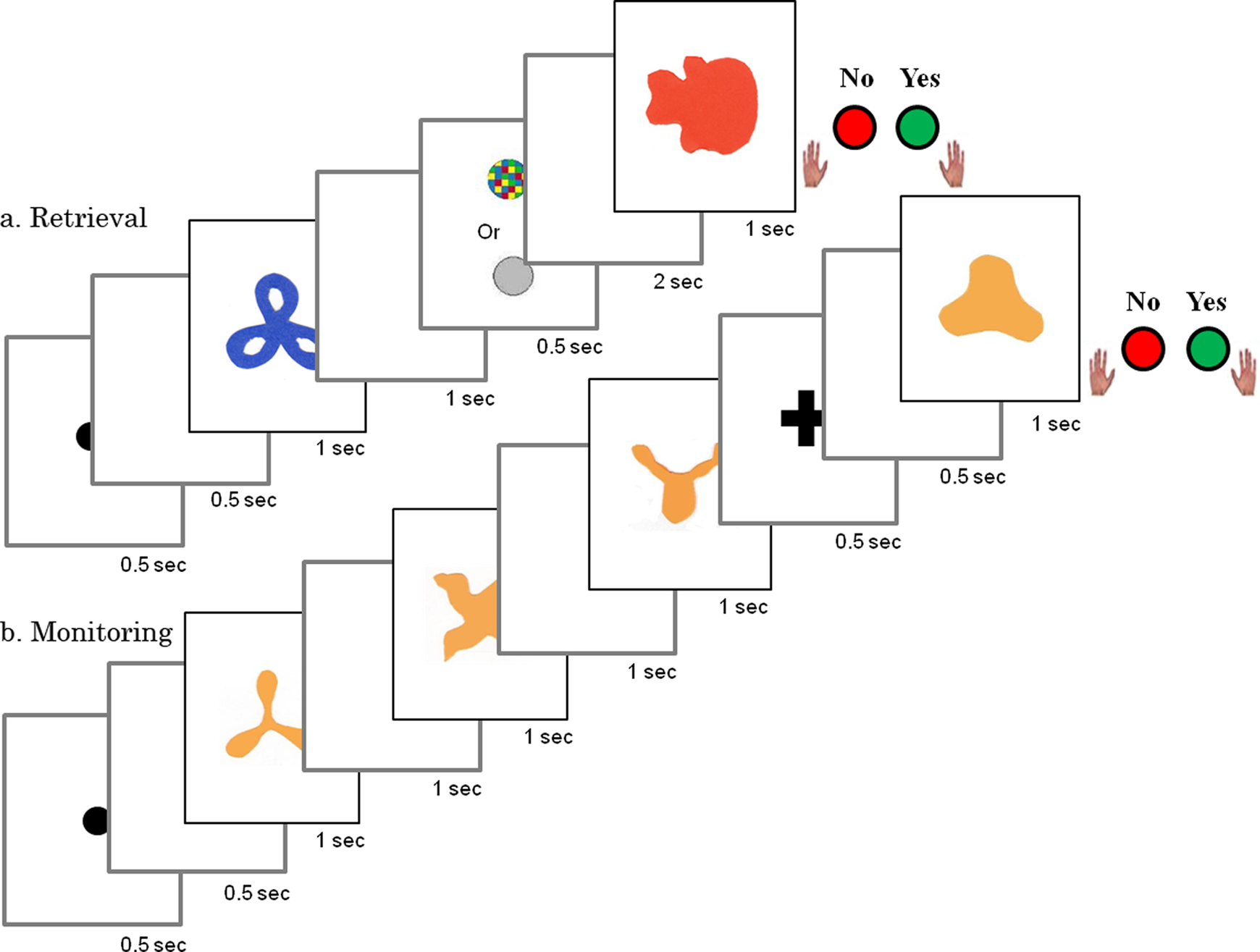
Schematic diagram of the active controlled retrieval and monitoring task phases.
Each trial in the monitoring task was initiated by the display of a fixation point at the center of the computer screen (Figure 1b). Three images from a bank of four were then presented sequentially for 1,000 ms each with a 1,000 ms delay between images during which the screen was blank. After the third image, a cross appeared at the center of the screen indicating that the test phase would follow. In the test phase, an image appeared and children had to decide whether or not it was the missing image from the bank of four. If it was, children had to press the YES button as quickly as possible, and if not, the NO button. Participants could decide that the probe was the missing image using deduction. If the status of each image was correctly monitored during the task, they could figure out after the third image what the missing picture was, and isolate it for the test phase. Children performed two series of 16 trials each. The first series contained images of four abstract shapes in the same color, and the second contained images of four different color chips. Figure 1b presents an example of a shape monitoring trial with a YES response. Before each series, children were invited to encode the four shapes or the four color chips, and the test began when they were comfortable to perform the task.
Tasks were presented in counterbalanced order. Participants were randomly assigned to one of four procedures: 1) Retrieval, Shape monitoring, Color monitoring; 2) Retrieval, Color monitoring, Shape monitoring; 3) Color monitoring, Shape monitoring, Retrieval; or 4) Shape monitoring, Color monitoring, Retrieval.
Apparatus
The experimental set-up comprised a computer screen placed in front of the children to present the stimuli and a response pad with two press buttons to record their responses. The green button was for a YES response and the red button for a NO response. Visual stimuli were a bank of 16 abstract shapes adapted from computer-generated Fourier descriptors (Tomita et al., 1999) and 16 color chips. Colors were distinctive primary colors. For example, in the bank, there were different types of blue (dark, light, etc.), but two types of blue were never presented in the same block of four colors to minimize the perceptual ambiguity in the task.
Blocks, each containing four shapes and four colors were randomly selected from the bank for the training and experimental series for both the retrieval and monitoring tasks.
Data analysis
Children’s performance was analyzed for accuracy (success rate, Sr) and speed (reaction time, Rt). Sr was calculated by summing the correct responses for each trial series expressed as the percent of correct responses on all trials. Rt was the mean Rt for correct responses. Five of the 87 children were identified as outliers 1 and were excluded from further analysis. An analysis of variance (ANOVA) revealed no significant main effect (p > .05) of sex or hand dominance for Sr and Rt on both tasks. An ANOVA was also conducted to test for whether children who performed more than one tutorial had an advantage over those who performed only one tutorial. Seventeen of the 82 children needed more than one tutorial for at least one test condition (active controlled retrieval, shape monitoring, or color monitoring), having failed on more than half the trials in the first tutorial. When performance on both tasks (active controlled retrieval and monitoring) was combined, these children scored an average of 48.5 correct responses out of 64, or 3.6 fewer than children who performed only one tutorial for each task. This significant difference (F(1, 80) = 5.36, p < .05) suggests that the second tutorial did not provide an advantage for memory performance.
To examine the relation between active retrieval and monitoring performances, Pearson correlations were calculated for Sr and Rt. Correlations were also calculated to test the relation between children’s age and performance.
Based on previous data showing the effect of visual characteristics on active retrieval (Dionne & Cadoret, 2013), the effect of age and visual characteristic was analyzed. Children were equally distributed into three age categories: younger from 80 months (6 years, 8 months) to 87 months (7 years, 3 months) (n = 27); intermediate from 88 months (7 years, 4 months) to 95 months (7 years, 11 months) (n = 28) and older from 96 months (8 years) to 103 months (8 years, 7 months) (n = 27). A repeated measures ANOVA with age categories (younger, intermediate, older) as factor and characteristic as within subjects factor with two levels (color and shape) was conducted on Sr and Rt for both functions. If the analysis of variance indicated significant effects, the Tukey HSD test was used to further explore significant interactions.
Then, a separate analysis was conducted for color and shape trials, because there was a significant effect of characteristics for each function (see below). A repeated measures ANOVA with age categories (younger, intermediate, older) as factor and function as within-subjects factor with two levels (active retrieval and monitoring) was conducted on Sr and Rt. If the analysis of variance indicated significant effects, the Tukey HSD test was used to further explore significant interactions.
Results
Active retrieval versus monitoring
Significant correlations were found between active controlled retrieval and monitoring for both Sr (r = 0.40, p < .001) and Rt (r = 0.51, p < .001). Significant correlations were found between age and active controlled retrieval and monitoring for both Sr (r = 0.32, p < .001), (r = 0.31, p < .001) and Rt (r = 0.23, p < .05), (r = 0.36, p < .001) respectively.
Success rate
For monitoring, the repeated measures ANOVA showed a main effect of characteristics, F(1, 79) = 32.45, p < .001, η2 p = .29, a main effect of age categories, F(2, 79) = 5.90, p < .01, η2 p = .13, but no interaction between characteristics and age categories (p = .94) on Sr. Further analysis with the Tukey HSD test showed that for all age categories combined, the success rate on shape monitoring trials was lower than on color monitoring trials (p < .001) (Figure 2A). Furthermore, the mean Sr in the older group for both shape and color monitoring was significantly higher than the mean Sr in the younger group (p < .05).
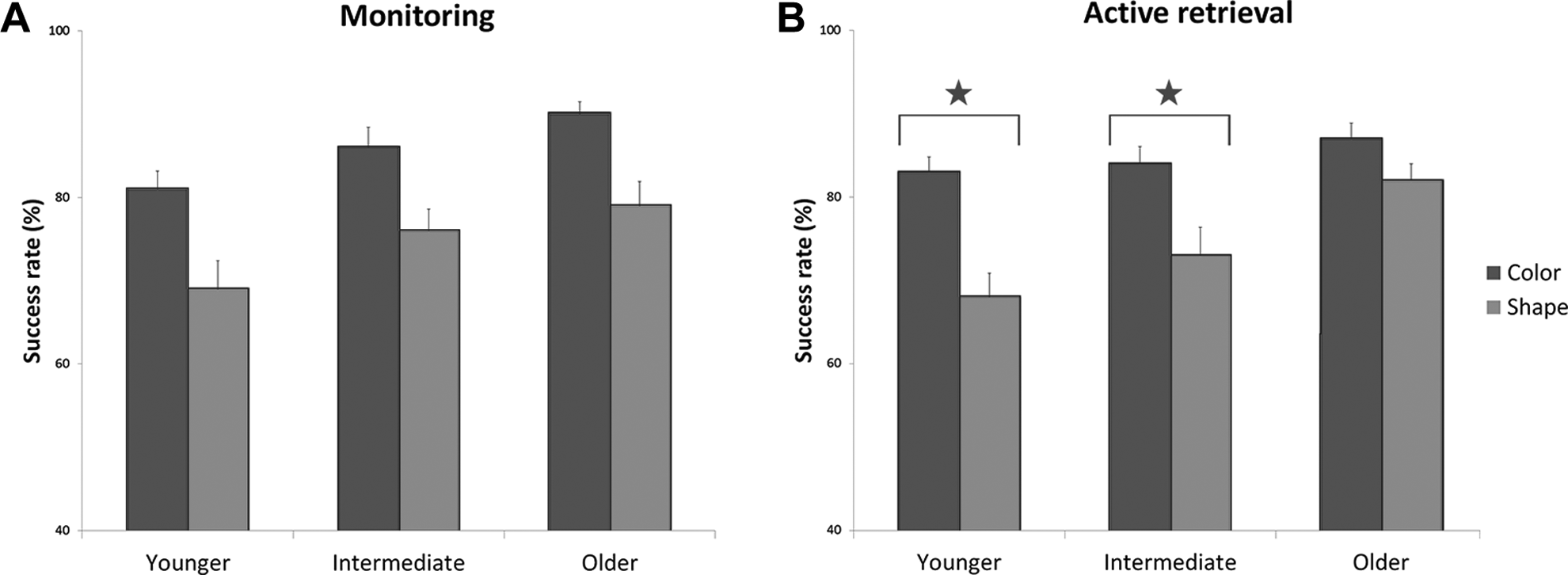
The mean success rate (+SE) for monitoring (A) and active controlled retrieval (B) for color and shape in the 3 age groups. Stars indicate significant differences at p < .05.
For active retrieval, the repeated measures ANOVA showed a main effect of characteristics, F(1, 79) = 38.22, p < .001, η2 p = .32, a main effect of age categories, F(2, 79) = 5.90, p < .01, η2 p = .13, and an interaction effect between characteristics and age categories, F(2, 79) = 3.21, p < .001, η2 p = .07, on Sr. Further analysis with the Tukey HSD test showed that the success rate on shape active retrieval trials was lower than on color active retrieval trials in the younger group (p < .001) and in the intermediate group (p < .01) but not in the older group (p = .58) (Figure 2B).
Considering the effect of characteristics on both monitoring and active retrieval, success rate on both tasks was compared separately for color trials and for shape trials. For shape trials, the repeated measures ANOVA showed a main effect of age categories on Sr, F(2, 79) = 6.58 p < .01, η2 p = .14, but no effect of functions (p = .89). For color trials, the repeated measures ANOVA showed a main effect of age categories, F(2, 79) = 4.66 p < .05, η2 p = .10, but no effect of functions (p = .59).
Reaction time
For monitoring, the repeated measures ANOVA showed a main effect of age categories, F(2, 79) = 5.80, p < .01, η2 p = .13, no effect of characteristics (p = .43) and no interaction effect between characteristics and age categories (p = .28) on Rt. Further analysis with the Tukey HSD test showed that the mean Rt in the older group (875 ms, SD = 199) was significantly lower than the mean Rt in the younger group (1006 ms, SD = 254) and in the intermediate group (1053 ms, SD = 225) (p < .05).
For active retrieval, the repeated measures ANOVA showed a main effect of characteristics, (1, 79) = 27.73, p < .001, η2 p = .26, an effect of age categories that was near significance (p = .054) and no interaction between characteristics and age categories (p = .91). Further analysis with the Tukey HSD test showed that the reaction time on shape active retrieval trials was higher than on color active retrieval trials (see Figure 3).
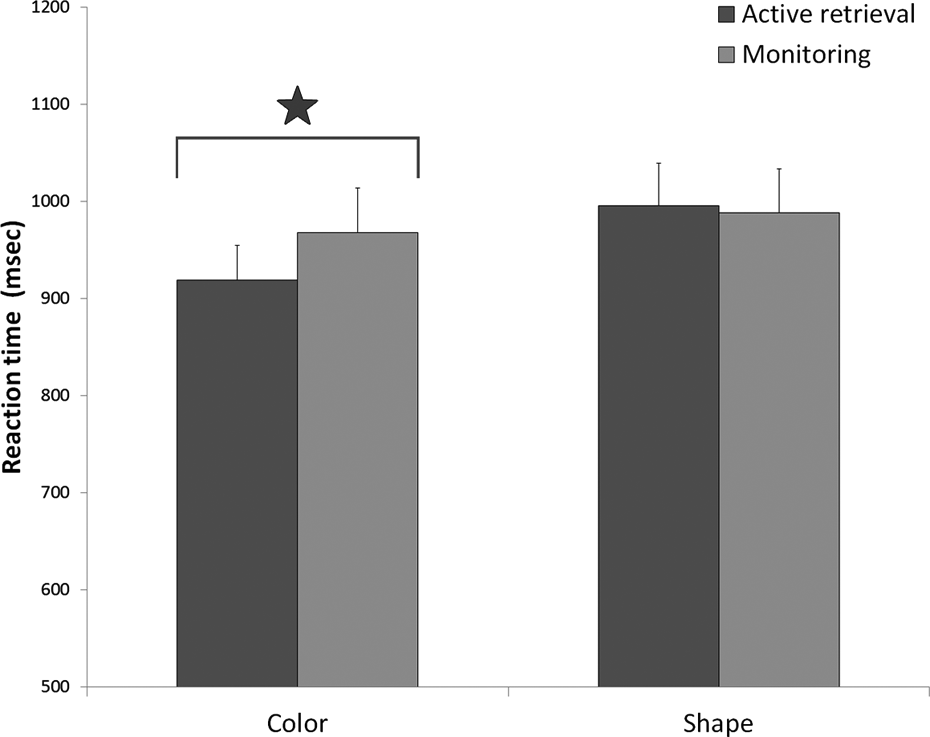
The mean reaction time (+SE) for active controlled retrieval and monitoring for color and shape. The star indicates a significant difference at p < .05.
Reaction times on active retrieval and on monitoring were compared separately for color and for shape trials. For shape trials, the repeated measures ANOVA showed a main effect of age categories, F(2, 79) = 3.18, p < .05, η2 p = .14, but no effect of functions (p = .81) on Rt. However, for color trials the analysis of variance showed a main effect of age categories, F(2, 79) = 7.53, p < .001, η2 p = .16, and a main effect of functions, F(1, 79) = 4.16 p < .05, η2 p = .05, but no interaction effect on Rt. As shown in Figure 3, the mean reaction time for color monitoring was higher than the mean reaction time for color active retrieval but there was no difference between active retrieval and monitoring reaction times for shape.
In sum the results show that children’s performance on active retrieval and monitoring were positively related. Children who performed better on active controlled retrieval also performed better on monitoring, and inversely, children who had more difficulty on one function also had more difficulty on the other. Success rates on both functions were equivalent and were higher when color had to be processed than shape. Reaction times on both functions were also equivalent for shape trials. However children were faster to report their decision when they had to retrieve the color than when they had to monitor the colors. Performance on both functions improved with age categories; older children were more accurate and faster than younger children. The difference between color and shape trials remained constant in monitoring but diminished with age categories in active retrieval.
Discussion
This study confirms that active retrieval and monitoring functions improve during middle childhood. It was initially predicted that children would score higher on active controlled retrieval than on monitoring, but the results demonstrated similar success rates in both functions. The only difference was that children were significantly slower to report their decision in monitoring than in active retrieval in color trials. Although small, this difference is interesting to note. Reaction time was measured when the test image appeared and at that time the demands were equivalent in both functions. Children had to perceive the visual display first, to decide whether the test image match or not the stored information and then to report their decision by pressing the left or the right key. The only difference between both functions was the processes engaged to select the stored information for the matching operation. As argued by Petrides (1995), active controlled retrieval and monitoring are distinct executive processes that must be distinguished from simple attention to a stimulus held in memory. They differ from mnemonic situations in which a stimulus in memory is attended to and the other stimuli remain in the background, and are not at the center of current awareness. Active controlled retrieval, which engages the MVLFC and not the MDLFC (Cadoret et al., 2001), implies that, on instruction, specific aspects of a representation are isolated from memory in high interfering situations. Ambiguity in memory representation can arise when stimuli are interrelated in multiple and more-or-less equiprobable ways, and are consequently weakly linked (Cadoret & Petrides, 2007). To isolate information, attention must be allocated to specific aspects in working memory while other stimuli that give rise to ambiguity must be suppressed. Monitoring within working memory through the MDLFC and not the MVLFC (Chen et al., 2004; Petrides, 1995) implies attention to a currently considered stimulus together with active consideration of several other stimuli whose current status is essential for making the decision. In the present study, the difference in reaction time for color trials suggests that monitoring the status of four colors in working memory was slightly more demanding than isolating one color from a multicomponent stored image. In contrast, isolating one shape in memory in a high interfering context was equivalent in terms of difficulty than keeping track of the status of four distinct stored shapes.
The expression of both executive memory processes was influenced by the visual characteristic that had to be processed, children scoring higher when color had to be retrieved and monitored than shape. As suggested by Prevor and Diamond (2005) children’s preference for shape or color appears to be highly dependent on the type of shape (e.g., geometric shapes, three-dimensional objects, everyday objects) and task demands (e.g., naming, matching, classifying). In the present study, color may have been easier to retrieve and monitor because the contrast between the different colors was more evident than the variations between shapes, and the colors were consequently less liable to be confused. Alternatively, the colors may have been more familiar than the abstract shapes, or they were easier to verbally label for further processing.
When performance was analyzed through age categories, it is interesting to note that active retrieval was the only function where the color shape difference diminished with age. Children’s success rate was lower for shape active retrieval than for color in the younger and intermediate groups but not in the older group. In contrast, the difference between color and shape remained constant in the three groups for monitoring. As shown previously (Arterberry, Milburn, Loza, & Willert, 2001; Dionne & Cadoret, 2013; Farrar & Goodman, 1992) children aged from 6 to 8 years old become more effective to disambiguate stored content to retrieve specific information in memory. Older children, in contrast to younger ones, demonstrated the same accuracy in shape retrieval than in color retrieval. However this improvement did not appear in monitoring, although older children were faster and more accurate than younger ones.
It was initially proposed that children would score higher on active controlled retrieval than on monitoring because this type of retrieval specifically engages the MVLFC, which is thought to mature first. However, no evidence of this difference was found, except a subtle difference in reaction time for color. Behavioral data do not provide direct evidence of brain functions but they can be considered as windows of brain maturation (Conklin, Luciana, Hooper, & Yarger, 2007; Luciana & Nelson, 1998, 2002; Welsh, Pennington, & Grossier, 1991). Furthermore, active controlled retrieval and monitoring processes have well established neuronal correlates in adults and these correlates have been defined in brain imaging studies with the same experimental tasks and with the same memory loading conditions as those used in the present study. It has been proposed that the MVLFC and the MDLFC play a critical role in controlling the mnemonic executive processes through their functional interaction with sensory-specific and multimodal posterior association cortical areas (Petrides, 1994). For example the decision that underlies active memory retrieval might be supported by a top-down modulation process exerted by the mid-ventrolateral prefrontal cortex on specific posterior association cortical areas in which memory representations are thought to be stored (Miyashita, 2004a, 2004b; O’Reilly, 2010). A similar performance in active retrieval and monitoring suggests that the developing neuronal mechanisms that support these functions have an equivalent efficiency during middle childhood. This provides additional support to the claim that the basic neural architecture of top-down control is present early in child development (Luna, 2009). Nevertheless, given the protracted development of the prefrontal cortex until late adolescence, this does not exclude that some differences can occur later in development. It has been shown that under conditions of high load and distracting information, the recruitment of prefrontal regions is increased (Bunge & Wright, 2007; Luna, 2009; Sander, Lindenberger, & Werkle-Bergner, 2012). More specifically, behavioral studies have shown that children monitor an increasing number of locations with age (Luciana & Nelson, 1998). In adults, Champod and Petrides (2010) showed a linear increase in activity in the MDLFC during the monitoring of a linearly increasing number of words in memory (from 3 to 5). It is possible that with maturation the MDLFC shows higher activation, allowing an increasing number of items to be monitored. To examine and compare the specific contributions of the developing lateral prefrontal regions in memory, it would be interesting to see if such a load effect can be observed in active retrieval associated with a linear increase in activity in the MVLFC.
Conclusion
These results showed that children have a variety of effective mechanisms to access the content of their memory. If the storage component of working memory has been largely examined in children, manipulation processes received much less attention and require more studies because of their essential role in cognitive development. The monitoring task was not an easy task, especially with shapes but the results showed for the first time that keeping track of the status of four complex visual elements in memory was as difficult as isolating a visual characteristic from a multicomponent stored image. While research in adults has shown that the lateral prefrontal areas have specific functional contributions in cognition (Petrides, 1996), these areas are often considered as a whole in developmental studies. A main contribution of this research was to address this issue of specificity of prefrontal functions in a developmental perspective.
Footnotes
Acknowledgments
We thank the children for participating in this study and the school professionals for their help.
Funding
This research was supported by funding from the University of Quebec in Montreal.