
Abstract
In 1995, Nelson published a paper describing a model of memory development during the first years of life. The current article seeks to provide an update on the original work published 20 years ago. Specifically, we review our current knowledge on the relation between the emergence of explicit memory functions throughout development and the maturation of associated brain regions. It is now well established that the brain regions subserving explicit memory functions (i.e. the hippocampal formation) are far from mature at birth, and exhibit important and gradual structural changes during childhood and beyond. Accordingly, explicit memory functions develop progressively. While some functions are present shortly after birth (formerly proposed as pre-explicit memory), others exhibit protracted developmental profiles during the first years of life. We examine the link between the emergence of different memory functions and the maturation of specific hippocampal circuits.
Introduction
In 1995, the second author of this paper published an article in Developmental Psychology entitled “The ontogeny of human memory: A cognitive neuroscience perspective” (Nelson, 1995). The paper was an attempt to provide a theoretical framework for explaining the neural bases of early memory development. The paper was long on inference and somewhat shorter on actual data, for the very simple reason that most of what was known at the time about the neural circuitry that underpins memory development was based on animal models and, in some cases, adult lesion models; much less was known about human brain development. Moreover, the behavioral literature had reached a point in its scientific maturity that few new insights were to be had from methods that were 30 years old (mostly looking-time tools). Nevertheless, the author proposed the following: that some elements of declarative or explicit memory developed very early in life, made possible in part by the precocious development of the hippocampus; in this context the notion of “pre-explicit” memory was proposed. However, more sophisticated forms of explicit memory, such as recognition memory that involved long delays between familiarization and test, cross-modal memory, and possibly event memory, were delayed in their emergence, due in part to more protracted development in a variety of structures that reside in the medial temporal lobe (MTL), such as the dentate gyrus, and in some cases, projections between these areas and their targets in the prefrontal cortex (PFC). These forms of memory came on-line in the second half of the first year of life, but continued to develop for years afterwards.
In contrast to the development of explicit memory, much less was known at that time about implicit memory. This was attributed to two factors: that many forms of implicit memory depend on motor activity, which is underdeveloped in infancy and, with the exception of work by Rovee-Collier and her colleagues, was largely unexplored; and that the circuits and structures involved in implicit memory (e.g., striatum, cerebellum) were both distributed and again, under-studied. Sadly, this remains a relatively unexplored area of inquiry, at least in relation to the early years of life.
In the current paper we attempt to revisit some of the ideas laid out in Nelson’s 1995 paper regarding the ontogeny of explicit memory. Over the past 20 years, tremendous progress has been made in studies of early brain development, and it is this work that will receive the bulk of our attention in this paper. However, some new insights have been gained on the behavioral side, and these, too, will be discussed. The goal of this paper, then, is not only to revisit some of the issues first discussed in the “Ontogeny” paper, but also to review new findings and, in turn, upgrade the views first proposed by Nelson in 1995.
Adult models of memory function
Memory is not a unitary function; that is, there is not just one type of memory but several types. This is true at the behavioral level but, more importantly, such segregation can be observed at the neural level. Early evidence of this position can be traced back to the 1950s, when Brenda Milner and her colleagues presented the case of Henry Molaison, better known as “H. M.” (Scoville & Milner, 1957). H. M. received a bilateral resection of the MTL (including the hippocampal formation and the amygdala, although see Annese et al., 2014 for a detailed and recent exposition of precisely which regions of the MTL were actually resected) in order to treat epilepsy. Although the surgery was successful at eliminating his seizures, it also led to partial retrograde amnesia (i.e., the loss of memory that was acquired before the onset of amnesia) that included several years preceding the surgery, and nearly complete anterograde amnesia (i.e., an inability to form new memories following the surgery). In other words, he was able to remember his childhood, but was unable to remember his doctor a few minutes after she had visited him in the hospital. Interestingly, this profound amnesic syndrome was not accompanied by any sensory, motor, or intellectual impairments (Milner, Squire, & Kandel, 1998). Furthermore, the most striking discovery was that H. M. was able to perform a number of specific learning and memory tasks (Corkin, 1968; Milner, 1962; reported in Milner et al., 1998). For example, H. M. was able to learn new motor skills and exhibited successful performance in perceptual learning tasks (Squire, 1992a; Squire & Wixted, 2011).
H. M.’s condition 1 served to highlight the distinction between two general types of memory: declarative or explicit memory and nondeclarative or implicit memory (Figure 1; see also Baddeley, 1976; Cohen & Squire, 1980; Sherry & Schacter, 1987). The terms explicit and implicit memory will be used here. Implicit memory was intact in H. M., although there were serious disturbances in explicit memory. Explicit memory is defined as the memory for facts about the world (semantic memory) and autobiographical events (episodic memory) and depends on MTL structures. Implicit memory, on the other hand, includes habits, skills, priming, classical conditioning, and nonassociative learning, and is subserved by a variety of brain regions such as the striatum, the cerebellum, or regions of the neocortex (Figure 1). The former will be the focus of this review.
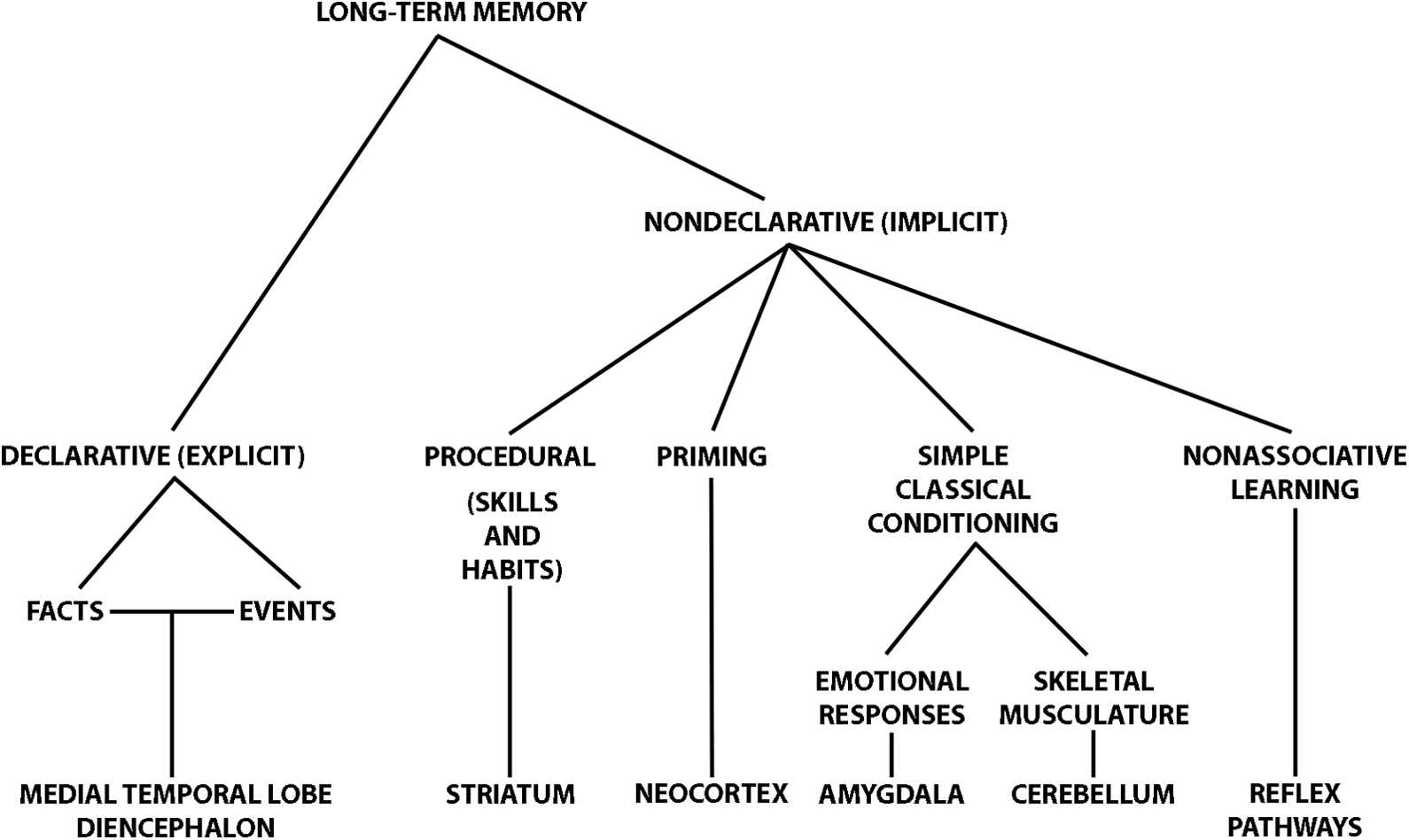
Taxonomy of the different memory systems and the brain regions subserving them. From Squire and Zola (1996), adapted from Squire and Knowlton (1995). Courtesy of The MIT Press.
It is now well established that the hippocampal formation is a functional entity fundamental for explicit memory processing. Indeed, a lesion circumscribed to this brain region leads to memory impairments in humans (Bright et al., 2006; Rempel-Clower, Zola, Squire, & Amaral, 1996), as well as in monkeys (Alvarez, Zola-Morgan, & Squire, 1995; Zola-Morgan & Squire, 1986). There is extensive literature about how, exactly, the hippocampal system might contribute to or subserve memory function. The hippocampal formation receives only highly processed and integrated information from unimodal and polymodal regions of the neocortex via the entorhinal cortex (Insausti, Amaral, & Cowan, 1987; Lavenex & Amaral, 2000; Suzuki & Amaral, 1994) and in turn further integrates and processes this rich set of information before sending even more highly integrated, complex information back to most of the regions of origin (Lavenex, Suzuki, & Amaral, 2002), where long-term memory is thought to be consolidated (Squire, Stark, & Clark, 2004; Wang & Morris, 2010; Winocur, Moscovitch, & Bontempi, 2010; for a recent review of the different theories on the topic see Winocur, Moscovitch, & Sekeres, 2013). However, the nature of this “integration” remains elusive. It has been proposed that the hippocampal system is involved in the “binding” of information from the neocortex (Eichenbaum, Schoenbaum, Young, & Bunsey, 1996; Marr, 1971; Squire, 1992b). Indeed, the organization of the hippocampal circuitry is particularly well suited to this purpose; the convergence of neocortical projections places the hippocampal formation in an ideal position to bind together information that is widely distributed in the brain (Squire & Zola-Morgan, 1991). However, this model considers the hippocampal formation as a single entity and does not define the specific role of distinct hippocampal regions in memory processing. The hippocampal formation is composed of distinct regions interconnected both serially and in parallel (Figure 2; Amaral & Lavenex, 2007): Neocortical information reaches the hippocampal formation mainly via the entorhinal cortex and is sent through the main hippocampal regions (first the dentate gyrus, then CA3, CA1, and finally the subiculum), before being sent back to the entorhinal cortex, thus closing a functional loop for information processing within the hippocampal formation. Furthermore, more recent studies of the neuroanatomical organization of the hippocampal formation have revealed a number of parallel pathways between different hippocampal regions (Figure 2; Amaral & Lavenex, 2007). Indeed, the entorhinal cortex projects in parallel to different hippocampal regions (CA3, CA2, CA1, and the subiculum, for example), which might therefore receive and process information independently from the region located upstream in the hippocampal loop for information processing described above.
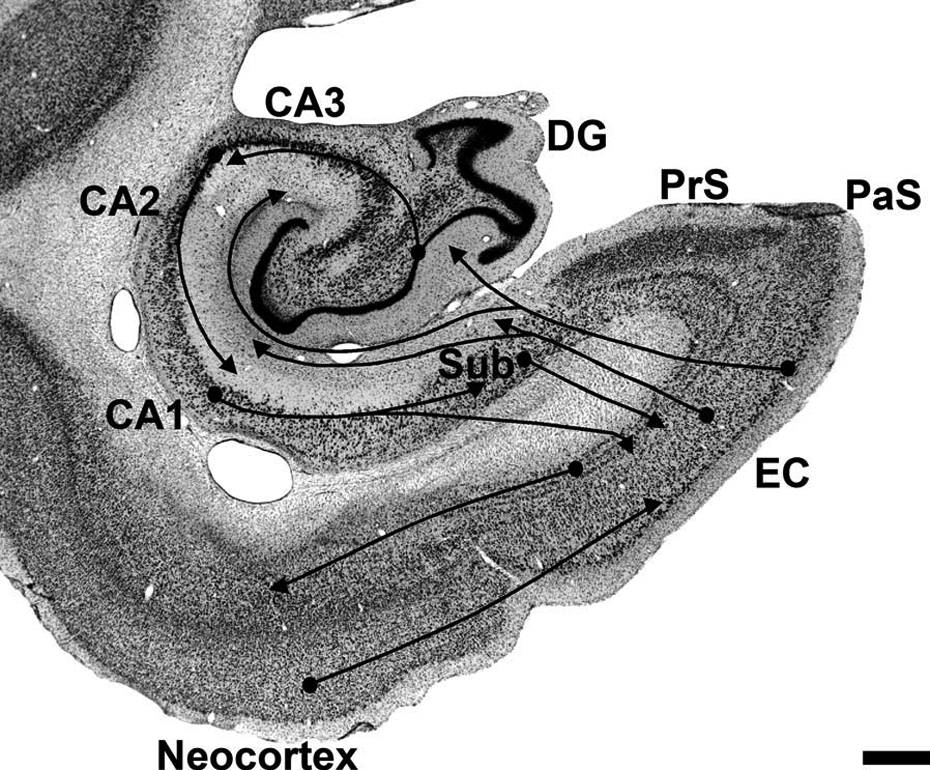
Schematic representation of the main serial and parallel pathways within the hippocampal formation. EC: entorhinal cortex; DG: dentate gyrus; CA3, CA2, CA1: fields of the hippocampus; Sub: subiculum; PrS: presubiculum; PaS: parasubiculum. Scale bar = 1 mm. From Jabès, Banta Lavenex, Amaral, and Lavenex (2011).
The hippocampal formation should therefore not be seen as a single entity, as it is made up of distinct regions and circuits that might subserve different types of explicit memory functions.
Differential maturation of the hippocampal formation circuitry
One striking advance in our understanding of memory organization has been provided by quantitative studies on the development of the hippocampal formation. It has long been thought that the hippocampal formation, with the notable exception of the dentate gyrus (which is known to be one of only two brain structure exhibiting neurogenesis postnatally), was quite mature at birth, subserving a wide range of memory functions (see Nelson, 1995). While this seems to be relatively true for some regions of the hippocampal formation, others appear to be far from mature at birth, exhibiting gradual and distinct developmental profiles postnatally (Favre, Banta Lavenex, & Lavenex, 2012; Jabès, Banta Lavenex, Amaral, & Lavenex, 2010, 2011; Lavenex, Banta Lavenex, & Amaral, 2007). However, we know little about how the differential maturation of the hippocampal regions is linked to the gradual emergence of explicit memory functions during development. In this section we review the quantitative neuroanatomical data collected in monkeys on the maturation of the hippocampal formation. In the next section we attempt to make links between the development of distinct hippocampal circuits and the emergence of specific memory functions.
The results of quantitative analyses of the structural development of the hippocampal formation in monkeys can be summarized in three points. First, they confirmed the protracted postnatal development of the dentate gyrus (neuron addition and maturation, volumetric expansion of individual layers) that extends beyond the first postnatal year (longer than previously thought; for a discussion see Jabès et al., 2010). Interestingly, the hippocampal regions and layers receiving strong direct or indirect projections from the dentate gyrus exhibit equally protracted developmental profiles (Jabès et al., 2011). Therefore, the late maturation of the dentate gyrus and the putative functional circuits downstream from it might subserve the late emergence of certain types of “hippocampus-dependent” memory functions. Second, the layers receiving direct entorhinal projections exhibit earlier maturation than the layers receiving indirect projections via the dentate gyrus and the rest of the hippocampal circuits (Jabès et al., 2011). This suggests that the differential and earlier maturation of entorhino-hippocampal circuits might subserve the earlier emergence of other types of “hippocampus-dependent” memory functions. Finally, some hippocampal regions (CA2, the subiculum, presubiculum, and parasubiculum) seem to be relatively mature at birth; therefore they develop earlier than their main source of excitatory inputs, that is, the dentate gyrus, CA3, and CA1 (Jabès et al., 2011). Interestingly, these four regions are characterized by their particularly significant connections with different subcortical regions (Amaral & Lavenex, 2007). These data suggest that hippocampal-subcortical circuits involving these four regions might mature the earliest, at birth or shortly after, and subserve the emergence of the earliest “hippocampus-dependent” memory functions.
In sum, it has been shown in the monkey that the hippocampal formation is far from mature at birth and exhibits important morphological changes beyond the first year of life (which roughly corresponds to 4 years of age in humans; Fortman, Hewett, & Bennett, 2001). Moreover, it has been suggested that the different circuits of this structure develop gradually from birth to adulthood, underlying the emergence of distinct memory processes (Jabès et al., 2011; Lavenex & Banta Lavenex, 2013).
Functions that depend on the hippocampal formation
Different human memory functions appear to emerge at different ages during the first years of life (Bauer, 2006; Newcombe, Lloyd, & Ratliff, 2007; Richmond & Nelson, 2007; for a recent review see Mullally & Maguire, 2014). However, the available information has long been insufficient to make links between the emergence of these different memory functions and the maturation of specific brain regions (i.e., the hippocampal formation and related brain structures; Lavenex et al., 2007; Nelson, 1998). In this section we review the functional changes occurring during this developmental period, and try to make links with what we know about brain maturation and specifically the development of distinct hippocampal circuits.
Recognition memory
One of the earliest-emerging memory functions is visual recognition memory, that is, the process by which an item is recognized as having been seen previously. This function has been widely assessed during infancy using the visual paired-comparison (VPC) paradigm (based on the natural preference of infants to look at a novel stimulus as compared to a familiar one; Fagan, 1977; Fantz, 1956) and seems to be functional shortly after birth (for earlier reviews see Nelson, 1995; Pascalis & de Haan, 2003). Monkeys 15 days of age have been shown to exhibit a novelty preference after a 10-second delay (Bachevalier, Brickson, & Hagger, 1993). In humans, novelty preferences after an interval of 2 minutes between familiarization and test have been observed only a few days after birth (Pascalis & de Schonen, 1994). Three-month-old children show stimulus retention after an interval of 24 hours (Pascalis, de Haan, Nelson, & de Schonen, 1998) and 6-month-olds after a 2-week delay (Fagan, 1973; but see also Pascalis & de Haan, 2003).
There are conflicting data on the role of the different MTL structures in recognition memory. For example, in adult individuals, it has been shown that VPC performance is severely impaired following lesions restricted to the hippocampal formation (Zola et al., 2000). In contrast, other studies have shown equivalent impairments in a VPC task after hippocampal or perirhinal lesions or even more severe deficits after perirhinal lesion (Nemanic, Alvarado, & Bachevalier, 2004; Pascalis, Hunkin, Holdstock, Isaac, & Mayes, 2004). The literature on the role of the hippocampal formation in recognition memory during development is quite sparse. Taylor, Mills, and Pang (2011) have shown right hippocampal activation during a face-recognition VPC task using magnetoencephalography (MEG) in children aged 6–19 years. Martinos et al. (2012) found a correlation between hippocampal volume and performance on a face-recognition VPC task in a population of 6-month to 5-year-old children who had experienced at least one episode of prolonged febrile seizures. To our knowledge, the only experimental study on the involvement of the hippocampal formation in recognition memory during the first months of life has been conducted on infant monkeys. This study showed that the performance of newborn and 6-month-old monkeys on a VPC task was not impaired at any delays between familiarization and test (10, 30, 60, and 120 seconds) despite substantial hippocampal lesions, suggesting the preferential involvement of other brain regions in recognition memory (i.e., the adjacent perirhinal and parahippocampal cortices) during early development (Zeamer, Heuer, & Bachevalier, 2010). Interestingly, however, the experimental lesions described in this study seem to have spared a large part of the subicular complex, a region of the hippocampal formation that has been shown to exhibit relatively early structural maturation in monkeys (Jabès et al., 2011; Lavenex, Banta Lavenex, & Amaral, 2004). Interestingly, the subiculum is one of the major output structures within the hippocampal circuitry (with the entorhinal cortex) and the principal origin of connections with subcortical areas (Amaral & Lavenex, 2007). Despite the obvious need for additional data to determine the neurobiological substrates that might subserve recognition memory at early ages, the current available evidence suggest that at least part of the hippocampal formation circuitry is relatively mature at birth and might subserve the early emergence of recognition memory around birth (Figure 3).
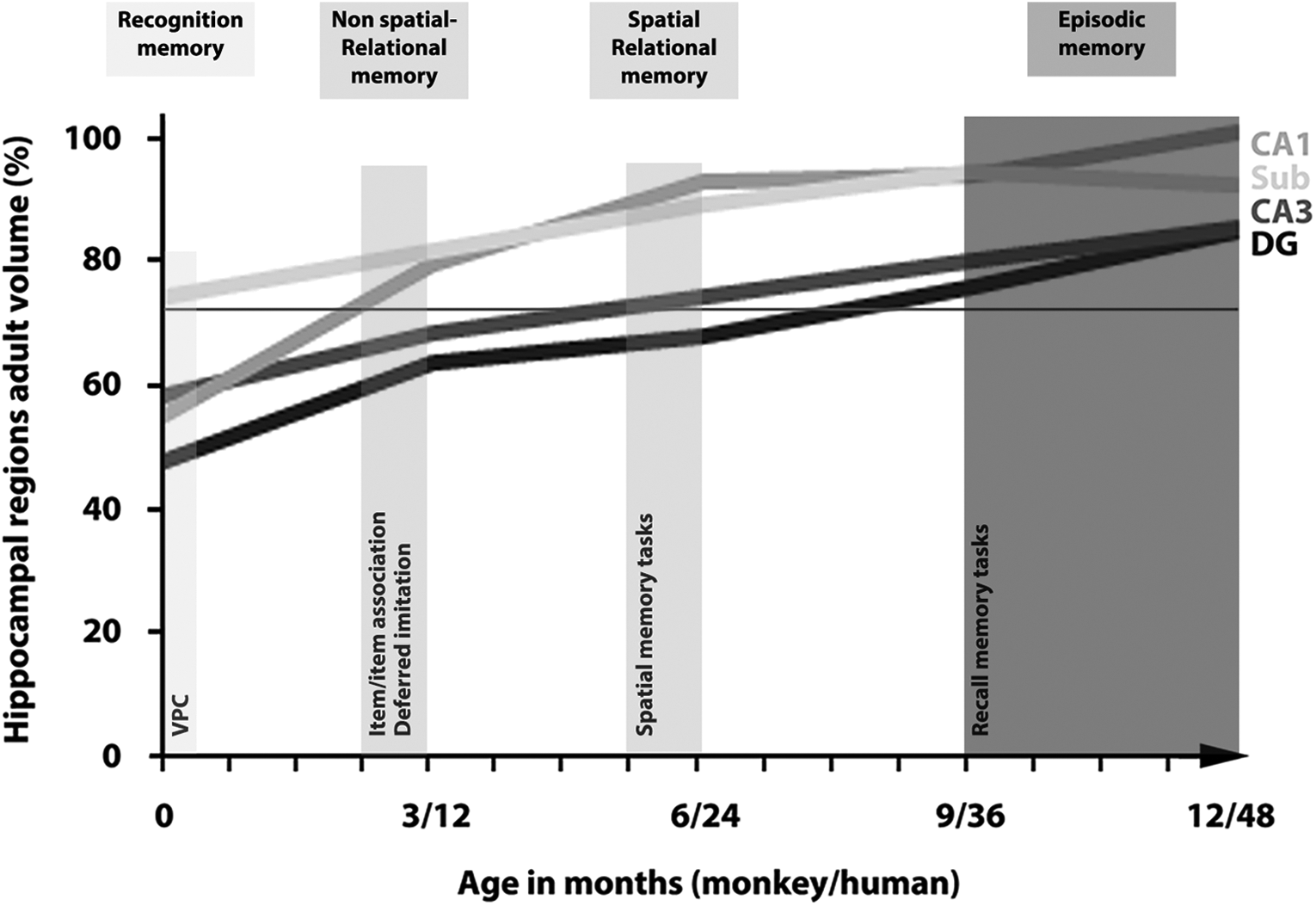
Schematic representation of the parallel development of the hippocampal regions in monkeys and the emergence of different explicit memory functions in humans. One year in monkeys corresponds to 4 years in humans (Fortman et al., 2001). DG: dentate gyrus; CA3, CA1: fields of the hippocampus; Sub: subiculum.
Basic relational memory functions
Relational memory, the representation of arbitrary or accidental relations among the constituent elements of an event (the relations between different objects on a picture or the temporal relations between actions; for a review see Konkel & Cohen, 2009), seems to emerge later than recognition memory. For example, it has been reported to be present at about 9 months of age in humans (Richmond & Nelson, 2009). In that study, infants looked at different pictures representing a face superimposed on a particular background (e.g., a pond with a water lily) during the familiarization phase. Then, during testing, infants were shown either (a) pictures of three familiar faces superimposed on one of the familiar backgrounds, that is, the face that had been presented with this particular background during familiarization (test background) and two other familiar faces, or (b) three familiar faces superimposed on a novel background. Using an eye-tracker system, this study showed that 9-month-old infants looked preferentially at the face that matched the test background. Thus, 9-month-old human infants are able to learn the relation between a given stimulus (a face) and a distinct background. Using a similar paradigm, Koski, Olson, and Newcombe (2013) failed to reproduce these results with older children. Indeed, 4-year-olds’ preferential looking at the correct face–background pair was at chance level. Moreover, 4-year-olds showed longer fixation to the face matching the test background only when they were consciously able to recall the correct face–background pair presented during familiarization.
Although the findings of Koski et al. (2013) add important new information about memory development in preschool-age children, it is difficult to interpret them in the context of findings from infants. Although it has been shown that preferential looking is a reliable tool for infants, it is less clear whether children and adults approach this task using the same set of neural and cognitive structures as do infants. For example, motivational factors could influence looking behavior, as might top-down processes that could lead children and adults to approach the task quite differently than do infants.
Deferred imitation tasks assessing the ability to reproduce puppets’ actions previously performed by an experimenter, that is, the ability to process relational information to create a representation of novel unrelated actions (Goertz, Kolling, Frahsek, Stanisch, & Knopf, 2008), also suggest that some forms of relational memory emerge between 6 and 12 months of age (Barr, Dowden, & Hayne, 1996). Indeed, some 12-month-old but not 6-month-old infants are able to reproduce, after a significant delay, a sequence of three actions that has previously been shown to them by an experimenter with a puppet; that is, they may learn the temporal sequence (the relations) among the constituent elements of a brief event.
Finally, spatial relational memory (or allocentric spatial memory; Nadel & Hardt, 2004; O’Keefe & Nadel, 1978), the ability to find a location in the absence of local cues marking that location, that is, the ability to find a location in relation to distant environmental cues (Schenk, Grobéty, Lavenex, & Lipp, 1995), emerges next. Indeed, 9-month-old monkeys (Lavenex & Banta Lavenex, 2006) and 25-month-old children (Ribordy, Jabès, Banta Lavenex, & Lavenex, 2013) have been shown to be able to find hidden food in an open-field arena in the absence of local cues. Accordingly, the ability of children to locate a hidden toy in a sandbox when distant environmental cues are available has been shown to emerge at around 21 months of age (Newcombe, Huttenlocher, Drummey, & Wiley, 1998).
In monkeys, the CA1 region of the hippocampal formation exhibits major structural changes between birth and 6 months of age (corresponding to 0–24 months of age in humans; Fortman et al., 2001), suggesting its possible involvement in nonspatial and spatial relational memory. Interestingly, it has been argued that CA1 is (a) involved in relational memory as assessed by contextual fear conditioning in rats (Daumas, Halley, Frances, & Lassalle, 2005); (b) necessary for the memory of sequences of events that compose unique episodes (Kesner, Hunsaker, & Ziegler, 2010), similar to what studies by Hayne and colleagues have tested using deferred imitation (Barr et al., 1996; Hayne, Boniface, & Barr, 2000); and (c) implicated in spatial memory, as CA1 “place-cells” have been reported to fire according to a rat’s location in the environment (O’Keefe & Dostrovsky, 1971; see also Jabès et al., 2011 and Lavenex & Banta Lavenex, 2013 for elaboration).
The gradual structural maturation of CA1 (between birth and 6 months of age in monkeys) might therefore subserve the incremental emergence of basic relational memory functions (spatial and nonspatial) observed between 9 and 25 months of age in humans (Figure 3). We will further develop this argument in the next section.
Complex relational memory functions
The ability to establish nonspatial relational memory representations (as assessed by the arbitrary association of items, like a face and its background or deferred imitation) might be rudimentary at first. Indeed, it has been suggested that this ability subsequently evolves during the first 2 years of life in humans to become more elaborate with age (Bauer, San Souci, & Pathman, 2010; de Haan, Mishkin, Baldeweg, & Vargha-Khadem, 2006; Jones & Herbert, 2006). For example, although 12-month-old children are able to perform a deferred imitation task after a 24-hour delay (Barr et al., 1996), their performance is disrupted if the puppet used to learn the sequence of actions (a pastel pink rabbit puppet) is different from the one used during the testing phase (a pale grey mouse puppet; Hayne, MacDonald, & Barr, 1997). In this study the two puppets shared many characteristics, such as the same overall shape, pastel color, eyes, and noses. Interestingly, if the two puppets are completely different (different overall shape, color, eyes, noses, ears, and mouths), 18-month-olds’ performance is also disrupted, but that of 21-month-olds is not. The same observations have been made with respect to recognition memory tasks. For example, it has been shown that a change in the background on which images are presented between the familiarization and testing phases disrupts recognition of these images at 6 and 12 months of age, but not at 18 and 24 months of age (Robinson & Pascalis, 2004). In addition, 6–9-month-old, but not 12–18-month-old, children’s recognition performance is disrupted if the familiarization and testing phases occur in two different rooms (Jones, Pascalis, Eacott, & Herbert, 2011). Similarly, a change in rooms does not disrupt 12–18-month-olds' performance in a deferred imitation task (Hayne et al., 2000) but it does disrupt the performance of 9-month-olds (Patel, Gaylord, & Fagen, 2013).
These data suggest that infants might learn the relation between items and their context, but that this relational representation is unitary at first (see below), such that retrieval is disrupted if components of the learned event are changed between the learning and test phases. In other words, relational memory is at first extremely specific to the context in which learning occurs and gradually becomes more and more “flexible” (Cohen, Poldrack, & Eichenbaum, 1997), allowing the generalization of learning to other conditions (for other examples, see Herbert & Hayne, 2000). This flexibility is a fundamental component of relational memory, which is thought to depend on the integrity of the hippocampal formation (Eichenbaum, 2004). The flexible use of memory is an ability attributed to the hippocampal formation, based on work with adults with hippocampal lesions (Pascalis, Hunkin, Bachevalier, & Mayes, 2009; Reber, Knowlton, & Squire, 1996; however, it should be noted that the extent of the lesions in these studies might exceed the hippocampal boundaries). We have proposed that CA1 subserves basic relational memory. Does the subsequent maturation of hippocampal regions upstream from CA1 subserve the qualitative changes observed in relational memory abilities? Interestingly, the neuroanatomical organization of CA1 has led to the hypothesis that this hippocampal region might subserve the integration of its inputs into a unitary representation (Rolls, 2010), whereas CA3, and its large associational network, stores the different parts of a representation separately. In rats, CA3 neurons have approximately 12,000 connections with other CA3 neurons (Rolls, Stringer, & Trappenberg, 2002). It has been proposed that CA3 is an auto-association network that facilitates the retrieval of a whole representation by activation of only a small part of this representation (i.e., reflecting the process of pattern completion 2 in a partial cue condition; Rolls & Kesner, 2006). In order to subserve this retrieval function, CA3 representations are stored in an unstructured manner, that is, a large number of memories can be stored and interference between memories is kept as low as possible due to the sparseness of its connections (Rolls, 2010). Therefore, parts of a representation are associated but remain distinct from each other. Such a network is thus resistant to the loss of information (Morris, 2007). Similarly, such a network allows the formation of a coherent representation that is not dependent (low interference) on its distinct parts. Thus, CA3 might allow the retrieval of a coherent representation that is kept intact even in a partial cue condition (flexibility). In addition, the dentate gyrus has been shown to subserve pattern separation 3 (Morris, 2007). For recent evidence on the role of CA3 and the dentate gyrus see Neunuebel & Knierim (2014). In contrast, CA1 cells, which have no associational connections, combine all information together into one crystallized representation (Rolls, 2010).
The argument that relational memory increases in complexity after 2 years of age in humans has been proposed by Lavenex and colleagues in the context of allocentric spatial memory development (Lavenex & Banta Lavenex, 2013; Ribordy et al., 2013). Indeed, whereas children 25–41 months of age were able to solve a basic allocentric spatial memory task, that is, locate one reward location among four possible locations, it is only after 42 months of age that children could locate three reward locations among 18 possible locations (Ribordy et al., 2013). These authors proposed that CA1 can subserve a basic form of allocentric spatial memory, at first through its direct projection to the entorhinal cortex 4 . Then, the subsequent maturation of CA3 and the dentate gyrus might result in the improvement of allocentric spatial memory abilities. As discussed above, the dentate gyrus and its projections to CA3 have been linked to pattern separation (Clelland et al., 2009), which might be necessary to disambiguate items, spatial locations, or episodes (see also below).
In sum, the behavioral and structural data available suggest that (a) the emergence of basic, context-specific relational memory might reflect the maturation of the CA1 region and its related circuits (see previous section and Figure 3), and (b) the emergence of a more “flexible”, context-independent relational memory might reflect the maturation of the CA3 and dentate gyrus regions and their related circuits.
Episodic memory
One of the latest-emerging hippocampal-dependent memory functions is episodic memory, the memory for events that occur in a unique spatio-temporal context (Tulving, 1972). Most adults are unable to recall events that happened before 2–3 years of age (Rubin, 2000; Jack & Hayne, 2010), a phenomenon often referred to as infantile amnesia (the inability to recall early life events). The underlying mechanisms leading to infantile amnesia are unclear (i.e. retrieval vs. storage impairments, for example). As reviewed above, infants have impressive memory abilities very early in life. But the information encoded early in life, however, might not be accessible to conscious retrieval later on (Newcombe & Fox, 1994). Infants might encode only bits and pieces—impressions if you will—of early life experiences (Freud, 1905/2001), but not coherent representations of past experiences that they can later recall (Fivush, Haden, & Adam, 1995). Thus, age-related qualitative changes in the kind of information encoded and how it is encoded might lead to the decline of infantile amnesia around preschool age (Bauer, 2002).
The neuroanatomical data that we have summarized so far indicate that most components of the hippocampal circuitry have reached relative volumetric maturity by 1 year of age in monkeys, which corresponds to roughly 4 years of age in humans. This development may ultimately depend on maturation of the dentate gyrus and its projections to CA3 (Jabès et al., 2011; Lavenex & Banta Lavenex, 2013). Together with recent computational models of neurogenesis and dentate gyrus function (see below), the current data suggest that this hippocampal region constitutes the last piece of the puzzle that is the formation of episodic memories. For example, the dentate gyrus has been shown to be involved in the formation of temporal associations between events (Morris, Curtis, Churchwell, Maasberg, & Kesner, 2013). In addition, continued neurogenesis in the dentate gyrus has been proposed to subserve the temporal coding of events and the formation of episodic memories throughout life (Deng, Aimone, & Gage, 2010; Rangel et al., 2014; Weisz & Argibay, 2009). It has been shown in rats that newly generated granule cells exhibit higher synaptic plasticity than do mature granule cells (Ambrogini et al., 2004; Schmidt-Hieber, Jonas, & Bischofberger, 2004). Moreover, it has been shown that neurons are more likely to be activated by events during their early maturation (Kee, Teixeira, Wang, & Frankland, 2007; Tashiro, Makino, & Gage, 2007). Therefore, events occurring close in time might be encoded by the same population of newly generated immature neurons, whereas events occurring several days apart might be preferentially encoded by distinct groups of newly generated neurons born at different points in time (Aimone, Wiles, & Gage, 2009). As the dentate gyrus granule cell population does not exhibit mature characteristics (neuron number, soma size) before at least 1 year of age in monkeys (Jabès et al., 2010), the balance between immature and mature neurons might not be optimally tuned to subserve episodic memory ability until late postnatal development. In addition, the number of newly generated neurons will likely disturb the existing networks by establishing new connections, which might dramatically impact information stored in these networks (for a review see Josselyn & Frankland, 2012). Whereas the temporal coding of events might first be subserved by CA1, allowing the encoding and remembering of a sequence of events happening over a short period of time (minutes), such as the learning of sequence of actions with a puppet as demonstrated by Hayne and colleagues in 12-month-old infants (Barr et al., 1996), the ability to create distinct memories of personal life episodes happening over a longer but restricted period of time (several hours or a day) might be dependent on the maturation of the dentate gyrus. Figure 3 illustrates the distinction between the emergence of relational memory and deferred imitation skills (the recall of a sequence of actions), which we propose to be dependent on the maturation of CA1, and the emergence of episodic memory (the recall of personal life events), which might depend on the maturation of the dentate gyrus.
Constructive constraints to the ontogeny of memory
As emphasized by Nelson (1995), it is fundamental to remember that the brain is an integrated system. A one-to-one relation between structure and function is therefore unlikely to be found. In an attempt to simplify, we have described the development of the hippocampal formation independently from the rest of the brain. However, the hippocampal formation is not an isolated brain structure; it is connected to a wide range of neocortical and subcortical brain regions. Thus, we can expect that the maturation of other brain regions and functions might impact hippocampal formation function. In this section we will give a few examples of the importance of considering the brain as a whole when studying the link between structure and function.
Different relational memory abilities, all proposed here to be dependent on CA1 circuits at first and to become more flexible as CA3 and the dentate gyrus reach maturity, are, however, emerging at different moments of postnatal development. For example, nonspatial relational memory emerges between 6 and 12 months of age and gradually becomes more sophisticated and flexible between 12 and 21 months of age; in contrast, basic spatial relational memory emerges around 21 months of age and subsequently becomes more elaborate after 42 months of age. The differential developmental profile of these two relational memory processes (spatial and nonspatial) suggests that they also depend on the maturation of other brain regions and circuits. For example, visual function, as assessed with the preferential looking paradigm, seems to emerge very early in infants (from the first days; see Pascalis & de Schonen, 1994). In contrast, locomotion (sitting, crawling, walking) emerges later (from around 7–10 months; see Bayley, 1936). Therefore, nonspatial relational memory as assessed with the preferential looking paradigm, dependent on visual abilities, might emerge earlier than spatial relational memory, dependent on motor abilities 5 .
The development of other cognitive functions seems to have an impact on learning, as was proposed long ago by Piaget (1964/1997), leading to additional qualitative changes in the different memory functions subsequent to their emergence. For example, although 12-month-old children can perform a deferred imitation task, they seem to recall less information than 18-month-olds (mean imitation scores increase with age; Barr et al., 1996). For example, the development of the PFC could account for such qualitative changes. Indeed, it has been established that PFC is implicated in episodic memory in adults (Badre & Wagner, 2007; Murray & Ranganath, 2007). It has been further suggested to be involved in the cognitive control of memory (Badre & Wagner, 2007) and specifically in the formation of detailed representations of experiences (Ofen et al., 2007). Interestingly, the PFC exhibits important structural changes late during development (for a review see Fuster, 2002; Ofen & Shing, 2013). Specifically, the functional interaction between the MTL and the PFC still increases during late postnatal development (Menon, Boyett-Anderson, & Reiss, 2005), potentially leading to the development of more effective memory processes. Functional changes have been shown to occur during a recall memory task in the PFC, the parietal cortex, as well as in the hippocampal formation between late childhood and adulthood (DeMaster, Pathman, & Ghetti, 2013). Thus, the protracted development of the PFC might lead to the development of richer representations, including the encoding of more contextual details, such as sensory, perceptual, cognitive, or emotional information (Drummey & Newcombe, 2002). For example, it has been shown that memory abilities continue to improve after the offset of infantile amnesia (Hayne & Imuta, 2011; Jack & Hayne, 2010; Newcombe et al., 2007; Pathman, Doydum, & Bauer, 2013; Sluzenski, Newcombe, & Kovacs, 2006). Indeed, children 3–7 years of age recall significantly fewer memories, which are less robust and less flexible, a phenomenon referred as childhood amnesia (Bauer, 2007; Newcombe et al., 2007). Such changes have been reported even until young adulthood (Ofen et al., 2007). Therefore, we might predict that the postnatal development of the PFC and its connections with the MTL might be involved in the gradual qualitative changes observed in infants and until young adulthood.
Finally, the postnatal development of connections between the hippocampal formation and other unspecified neocortical brain regions might also lead to qualitative changes in memory processing. For example, it has been shown that retention time increases as a function of age (Scarf, Gross, Colombo, & Hayne, 2013; for a discussion see Bauer, 1996). Various unimodal and polymodal neocortical regions that send information to the hippocampal formation receive feedback information after hippocampal processing (Lavenex et al., 2002), and are thus thought to subserve consolidation of this processed information in long-term memory (Squire et al., 2004; Wang & Morris, 2010; Winocur et al., 2010). It is therefore reasonable to hypothesize that hippocampal projections toward the neocortex (via the entorhinal cortex) or the targeted neocortical regions themselves might mature gradually throughout postnatal development. Consistent with this proposal is the observation that structural changes occur throughout childhood and adolescence in the neocortex (for a review see Marsh, Gerber, & Peterson, 2008).
Concluding remarks
In this work we highlighted the extraordinary and quite successful efforts researchers in the field of memory development have made in the past 20 years to improve our understanding of the ontogeny of explicit memory. Here we propose and review several hypotheses linking the maturation of hippocampal regions with the extensive literature on the emergence and refinement of explicit memory functions.
The present data indicate that despite its late maturation, the hippocampal formation might subserve a wide range of memory functions as early as the first days of life. Indeed, it is now clear that the hippocampal formation is not a unitary structure but, rather, is composed of several subregions, all developing at different points in time. Nelson (1995) originally proposed that early explicit memory likely depends on those regions of the brain, fostering the early development of so-called pre-explicit memory, followed by a more mature form of explicit memory that develops over the first 1–2 decades of life. The available data confirm this view, suggesting the differential maturation of specific circuits subserving the gradual emergence of explicit memory functions. While some hippocampal circuits are relatively mature at birth (e.g., the subiculum), potentially subserving (pre-)explicit memory functions (i.e., recognition memory), others exhibit a more protracted developmental profile. It is proposed that the maturation of CA1 circuitry might subserve the emergence of rudimentary explicit memory functions at first (nonspatial and spatial relational memory). This basic form of explicit memory will gradually become more complex and flexible due to the maturation of CA3 and the dentate gyrus circuitry, which will ultimately lead to the emergence of episodic memory and the ability to build and remember representations of our everyday experiences.
Future work aiming to describe the structural maturation of the brain circuits subserving memory (i.e., the hippocampal formation and its major connections with subcortical areas or regions of the frontal cortex for example), in conjunction with advances in behavioral and neuroimaging measures (i.e., electroencephalography/event-related potential, functional near-infrared spectroscopy, and magnetoencephalography) that can be applied to very young infants, will certainly further our understanding of memory organization and development in the years to come.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.