
Abstract
Considering the wide and increasing utilization of drugs which act upon the central nervous system, a pressing, infrequently investigated question is their potential impact on sleep-associated memory consolidation. Here the influence of approved pharmaceuticals on memory consolidation during sleep in healthy humans, rats, and mice was investigated. A systematic review of the literature was conducted according to PRISMA guidelines. Criteria to be met included healthy subjects, drug application after learning and a comparable placebo condition. Exclusion criteria included but were not limited to repetitive dosing and additional non-pharmacological interventions. A total of 65 studies were included, whereby a limited number of studies lent themselves to a meta-analysis. The risk-of-bias assessment showed a lack of reporting predominantly in animal studies. Meta-analysis was conducted for the influence of zolpidem on consolidation of the word-pair association task in human adults, which showed an improvement in memory consolidation (0.57 (95% confidence interval 0.04–1.10)). Most importantly, for memory consolidation, results point toward a favorable position of second-generation antidepressants compared to their first-generation counterparts. Knowledge of such cognitive effects in healthy subjects should encourage consideration in designing studies with patients and/or in patient care.
Introduction
The ability to form memory contents is crucial for organisms to adapt and thrive in a changing environment. A plethora of evidence indicates that sleep constitutes a critical neurobiological state for memory consolidation, at the systems- and cellular level (Brodt et al., 2023; Hoedlmoser et al., 2022; Marshall, 2020). Considering an increase in chronic and subchronic use of the investigated pharmaceuticals (Alabaku et al., 2023; Bushnell et al., 2025; Hafferty et al., 2019; Qingyu et al., 2025), knowledge of potential effects on sleep-associated memory consolidation is highly relevant. However, comprehensive reviews examining the impact of approved and clinically relevant pharmaceuticals on sleep-associated memory consolidation remain scarce. This gap defines the objective of our analysis.
The formation of memories consists of at least three prominent phases: encoding, consolidation, and recall. Encoding and recall typically take place during the active waking state, while consolidation, not only in hippocampus-dependent tasks, is supported by offline neuronal reactivation processes that unfold during sleep, but also during quiet wakefulness. These processes involve the reactivation and reorganization of neuronal networks, facilitating the stabilization and integration of newly acquired information (Halpern et al., 2025; Klinzing et al., 2019; Rasch and Born, 2013; Tonegawa et al., 2018; Wamsley, 2019). Although initial studies investigated hippocampal reactivation during deep non-rapid eye movement (NREM) sleep, the concept of reactivation during sleep as a mechanism for offline learning has been extended to non-hippocampus-dependent tasks, whereby temporal relationships between encoding and reactivation are task-dependent (Klinzing et al., 2019; Paller et al., 2021).
Macro (sleep stages) as well as micro dynamics (within a sleep stage) of sleep brain rhythms are largely generated and influenced by brainstem nuclei and their neuromodulatory systems (Monti et al., 2008b). For instance, the temporal dynamics of brainstem locus coeruleus (LC) activity and noradrenaline release have been linked to brain rhythms of sleep and memory consolidation (Eschenko et al., 2012; Kjaerby et al., 2022; Swift et al., 2018). In the same vein, the balance of the main inhibitory and excitatory neurotransmitters of the brain, γ-aminobutyric acid (GABA) and glutamate are involved in sleep regulation and shape long-term synaptic plasticity (Feld and Born, 2020; Helfrich et al., 2021).
The objective of this systematic review is to assess the influence of three approved pharmaceutical groups: antidepressants, anesthetics and sedatives/hypnotics on sleep-related memory consolidation in healthy subjects (humans, rats, and mice) versus placebo. We focused on antidepressants and sedatives/hypnotics, because these drugs are strongly modulated by sleep relevant neurochemicals. Furthermore, their use has increased markedly in young adults (Bushnell et al., 2024; Chua et al., 2024; Kwok et al., 2025) while studies on adverse central nervous system (CNS) effects appear underrepresented in the literature. Regarding anesthesia, although normal sleep is not induced, some mechanisms and electrophysiological phenomena overlap (Alkire et al., 2008; Brown et al., 2010; Moody et al., 2021; Tononi and Sporns, 2003). Further interest in this group arose due to increased use in non-hospital settings, also for procedural sedation with subanesthetic doses (Mandarino et al., 2026; Pribish et al., 2020; Quintero et al., 2025; Vasileiou et al., 2009). Drugs with high affinity to specific receptors are limited in their use in humans, therefore we chose to study only drugs approved for human use. A general property is the affinity of these drugs to receptors beyond the primary target, limiting a priori a clear mechanistic interpretation.
The main mechanisms of action of the investigated drug classes are indicated in influencing both sleep and neuroplasticity: Effects of the antidepressant group are largely attributed to the modification of monoamine levels, namely of noradrenaline and serotonin. These neurotransmitters have been extensively implicated in memory consolidation at both cellular and systems levels (Coray and Quednow, 2022; Higa et al., 2024; McGaugh, 2002; McGaugh and Roozendaal, 2009; Power et al., 2002). For sedatives/hypnotics and anesthetics GABA receptor modulation and or the relationship between GABA receptor modulation and N-methyl-
To provide a comprehensive overview of the field, as is our objective, we included a broad range of memory paradigms. While this inclusive approach enhances scope, it also necessitates caution in interpreting results, as differential underlying neurophysiological mechanisms limit direct comparability across studies.
Methods
We present a systematic review with meta-analyses for studies of sufficient homogeneity regarding included drug, population, and the reported outcome measures. The PRISMA statement (Page et al., 2021) was used to enhance the documentation of the research process and proposes a number of items that should be included when reporting a systematic review: A review protocol was written and a preregistration on PROSPERO took place before the start of the review process. No automation tools were used in any part of the process.
Inclusion and exclusion criteria
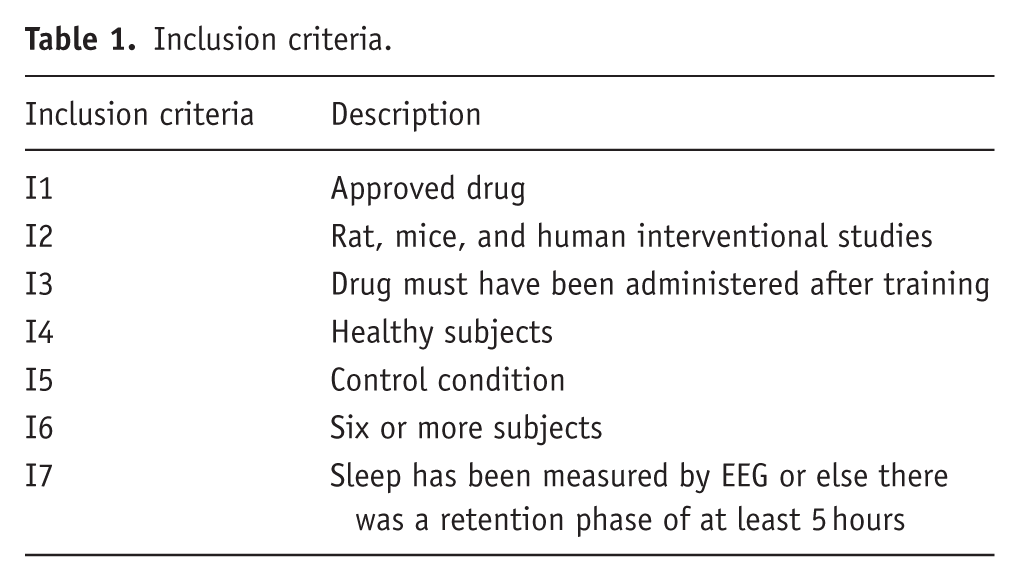
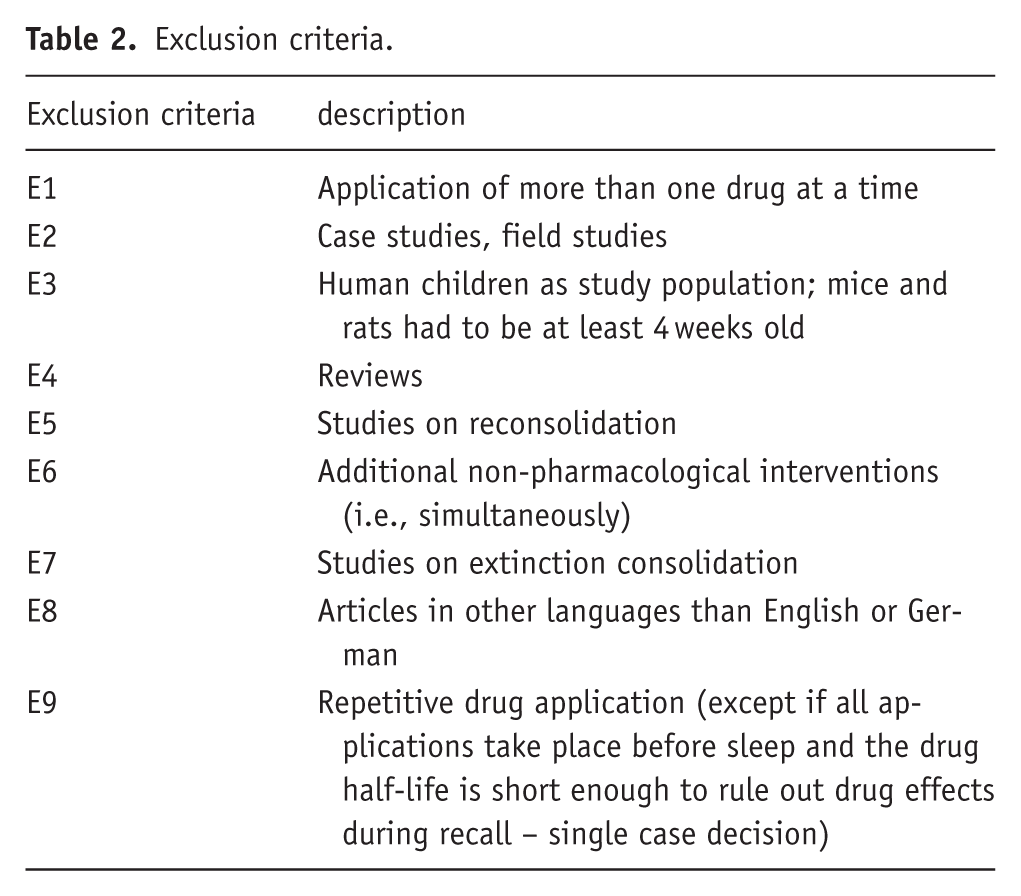
This research focuses on the influence of pharmaceuticals on memory consolidation. The original studies had to meet specific requirements to ensure their design was suitable for identifying the influence of the pharmaceutical on the consolidation phase. To exclude that any effect on memory consolidation was due to a modulatory effect on learning or retrieval processes the timing of drug application was required to abide by two rules: (1) It must occur after termination of the encoding phase and (2) a sufficient time (retention) interval must pass before the (recall) test to avoid any influence of the drug on the retrieval process. These temporal aspects are reflected in the inclusion and exclusion criteria I3, I7, E9. The latter takes drug half-life into account (Tables 1 and 2).
Inclusion criteria.
Exclusion criteria.
Notably, sleep EEG and stages were rarely reported, thus necessitating an inclusion criterion based on a minimal retention time. Figure 1 reveals that despite differences in temporal structure and circadian phase (Soltani et al., 2019), sleep in all three species includes rapid eye movement (REM) and NREM sleep (Bastianini et al., 2014).

Cross-species NREM and REM sleep composition. Exemplary sleep architecture of an 8-hour sleep period in a human subject (top), and of a 12-hour lights-on period in a rat (middle) and mouse (bottom).
When subject age was not provided, an adult population was assumed. All included studies were grouped by drug group (antidepressants, anesthetics, and sedatives/hypnotics) and further separated according to the individual drug, species, and memory task.
Search methodology
First, key terms were identified. These included not only drug names but also drug group names such as “antihistamines” or “antidepressants.” This ensured that all substances belonging to the searched group were identified. All terms describing the drugs or drug groups were combined with the operator “OR” and the key terms “memory” and “consolidation” were added by using the operator “AND.”
On the chosen databases, PubMed and Web of Science, the final searches were executed on the following dates:
- sedatives/hypnotics and anesthetics: June 30, 2022 (PubMed and Web of Science)
- antidepressants: October 3, 2022 (PubMed) and October 6, 2022 (Web of Science).
The complete search terms can be found in the Supplemental Material.
Studies on anesthetics and sedatives/hypnotics were searched collectively because of the similar subject area and later separated according to drugs at the level of the full-text screening. Morphine was initially included in the search for anesthetics because it is used intraoperatively. According to the anatomical therapeutic chemical (ATC) classification, however, it does not belong to the general anesthetics and was therefore subsequently removed from the analysis. Some benzodiazepines are not classified under hypnotics and sedatives in the ATC code, but under anxiolytics. Nevertheless, all benzodiazepines were included in the evaluation to represent the entire class of chemical substances. The PubMed MeSH terms function also automatically completes the search queries. The exact search history including each inclusion or exclusion decision is available upon request.
Selection of studies included
For the purpose of this review the definition for antidepressants is any drug that is included in the German ATC code N06A (without N06AP (herbal antidepressants) or N06AH (homeopathic antidepressants)); N05AN01 (Lithium), an antipsychotic, was additionally included because it is used as maintenance therapy not only for bipolar but also for unipolar depression (Abou-Saleh et al., 2017; Coppen, 2000). An anesthetic is defined as any drug included in the German ATC code N01A, and hypnotics and sedatives are defined as any drug included in the German ATC code N05C (without N05CP (herbal hypnotics and sedatives)); N05BA01 (Diazepam) and N05BA06 (Lorazepam) were additionally included. Ketamine potentially fits into two categories. Only (S)-Ketamine is included in the antidepressant category because no preparations for depression containing the racemate or (R)-Ketamine are approved. Preparations with the ketamine racemate are included in the anesthetics category, since the racemate is used in clinical practice. Additional information can be found in the Supplemental Tables 1 and 2.
The program Rayyan, which allows co-working of several editors was used to manage literature during the review process (Ouzzani et al., 2016). For the study selection, first all articles were screened by title and abstract after duplicates were removed. The screening was performed blindly by two independent researchers. The articles were rated: “included,” “excluded,” or “maybe.” Articles that received the rating “included” or “maybe” from at least one researcher were reviewed as a full-text article in a second step. The number of articles excluded in this second step is shown in the Supplemental Material with reasons for their exclusion (Supplemental Figures 2 and 3). All articles receiving discordant ratings from reviewers following full-text screening were reviewed by a third researcher, who made the final decision on study inclusion or exclusion. The third researcher was not blinded to the previous reviewers’ decisions but rather read the given reasoning.
Methods of data extraction
Data was extracted from the articles manually by one researcher using a predefined data extraction sheet. Data were compiled in an excel file.
Risk-of-bias analysis
In animal studies, use of methods to minimize risk of bias is not yet standard practice, hence neither is their reporting. Also, some aspects of randomized controlled trials in humans cannot be accounted for in animal studies. Since this review includes animal and human interventional studies we selected the Syrcle risk-of-bias tool for animal studies that is best appropriate for the purpose of this review (Hooijmans et al., 2014). The following items were chosen: D1: Were baseline characteristics between groups similar? (selection bias); D2: Randomization? (selection bias); D3: Were caregivers/investigators blinded? (performance bias); D4: Were outcome assessors blinded? (detection bias); D5: Were subjects/data excluded and why? (attrition bias). The studies were assessed by one researcher. A random sample of about 33% of the studies was also assessed by a second researcher independently.
The results of the risk-of-bias assessment were then visualized using an online tool (McGuinness and Higgins, 2021).
Synthesis and analysis
Synthesis
Studies are summarized in Tables 3 to 5. The study characteristics include author, publication year, investigated drug, study population, their sex and age, time of experiment, retention interval, use of EEG/local field potential (LFP)/polysomnography (PSG) for sleep measurement, memory task, retention performance, effect size, sample size and mechanism of action. Each pharmaceutical category was grouped individually.
Quantitative analysis
Statistical analysis was only conducted when more than two studies were found that were homogeneous in the categories:
Investigated drug
Investigated species including strain of animals
Task used to investigate consolidation
Available data (means and standard deviations, or alternatively t-values and number of subjects) were extracted from the studies. Statistical analyses were conducted using Meta-Essentials v1.5 (Suurmond et al., 2017), “workbook 3 – differences between independent groups with continuous data” CC BY (https://creativecommons.org/licenses/by-nc-sa/4.0/). Weights were automatically assigned to the studies. A random effects model was chosen due to different retention intervals. Assuming that consolidation is a time-dependent process, measured effects likely differ if retention intervals vary. Hedges’ g was chosen as the effect measure, because it provides less distorted estimates in small samples than Cohens’ d.
Precision of the results was discussed using the 95% confidence interval (CI). Heterogeneity was discussed using I2, Tau, and the prediction interval (PI).
Results
The search process led to 367 entries for antidepressants and 868 entries for anesthetics and hypnotics and sedatives combined (see Supplemental Figures 2 and 3 for corresponding PRISMA 2020 flow diagrams).
Exclusion after full-text screening was mainly due to a failure to meet inclusion criteria I1 (approved drug) and I3 (drug must have been administered after training).
Overall study quality was good in the reported risk-of-bias items. The item least reported was D4 (detection bias) with 54 overall “no information” entries, of which 42 were in animal studies and 12 in human studies. The best reported item was D3 (performance bias) in human studies with only one study not providing information. Reporting was best overall and in animal studies in category D1 (selection bias) with 28 and 26 “no information” entries, respectively. In general, reporting in human studies was substantially more comprehensive than in animal studies (cp. Supplemental Figure 1a and b).
The congruence between the two researchers for risk-of-bias assessments was 82%.
Although nighttime and daytime were often mentioned, it was rarely described whether training or testing or both were performed during these times. With retention intervals of 24 or 48 hours, it was assumed that both training and testing were performed in the specified phase.
The following results were derived from qualitative analysis unless statistical analyses are specifically referred to.
Antidepressants
For antidepressants, 25 reports of 25 studies were included in the analysis, a quantitative analyses was not performed in this category. The results for all antidepressants are shown in Table 3. Five human studies and 20 animal studies were found. Fourteen different antidepressants were investigated. Memory tasks included passive avoidance tasks, novel object recognition tasks, fear conditioning, finger tapping tasks, mirror tracing tasks, visual discrimination tasks, autoshaping, and word paired-associate tasks (WPA). Three studies did not state subjects’ sex, four studies included female and male subjects, and most studies only included male subjects. Six studies used EEG/LFP/PSG measurements. Across all studies measuring EEG and/or PSG the suppression of REM sleep was most characteristic.
Results antidepressants.
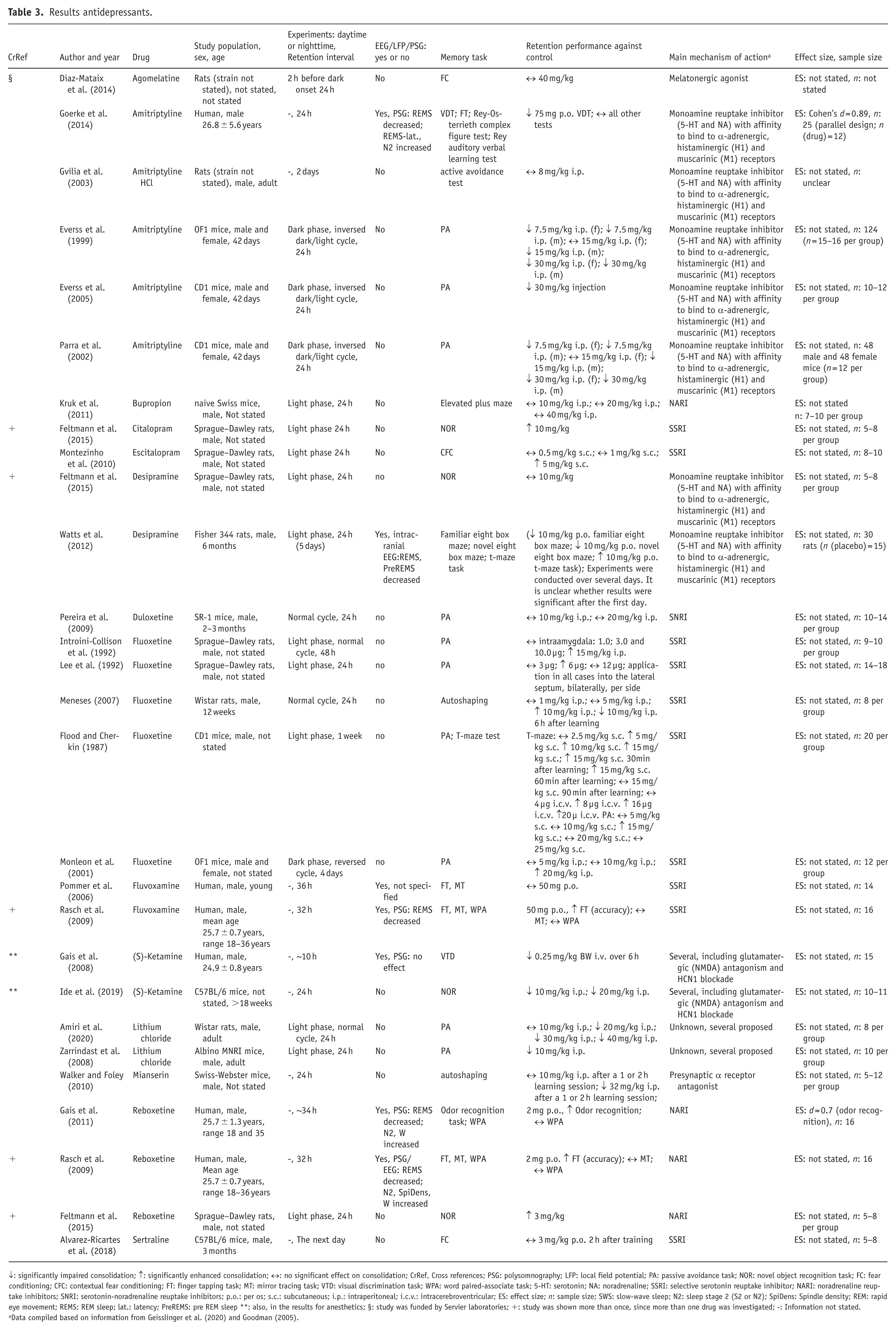
↓: significantly impaired consolidation; ↑: significantly enhanced consolidation; ↔: no significant effect on consolidation; CrRef, Cross references; PSG: polysomnography; LFP: local field potential; PA: passive avoidance task; NOR: novel object recognition task; FC: fear conditioning; CFC: contextual fear conditioning; FT: finger tapping task; MT: mirror tracing task; VTD: visual discrimination task; WPA: word paired-associate task; 5-HT: serotonin; NA: noradrenaline; SSRI: selective serotonin reuptake inhibitor; NARI: noradrenaline reuptake inhibitors; SNRI: serotonin-noradrenaline reuptake inhibitors; p.o.: per os; s.c.: subcutaneous; i.p.: intraperitoneal; i.c.v.: intracerebroventricular; ES: effect size; n: sample size; SWS: slow-wave sleep; N2: sleep stage 2 (S2 or N2); SpiDens: Spindle density; REM: rapid eye movement; REMS: REM sleep; lat.: latency; PreREMS: pre REM sleep **: also, in the results for anesthetics; §: study was funded by Servier laboratories; +: study was shown more than once, since more than one drug was investigated; -: Information not stated.
Data compiled based on information from Geisslinger et al. (2020) and Goodman (2005).
Dividing the antidepressants into groups, with one containing second-generation (selective serotonin reuptake inhibitor (SSRI), noradrenaline reuptake inhibitor (NARI), serotonin-noradrenaline reuptake inhibitor (SNRI)), and one containing antidepressants developed longer ago (tricyclic and tetracyclic antidepressants), shows a tendency for improved consolidation with second-generation antidepressants and a tendency toward impairment in the other group.
Anesthetics
For anesthetics, 24 reports of 23 studies were included in the analysis. Quantitative analyses were not performed in this group. The results for anesthetics are shown in Table 4. Four human studies and 19 animal studies were found. Six different anesthetics were investigated, and the memory tasks included WPA, finger tapping tasks, 2D face location tasks, visual texture discrimination tasks, novel object recognition tasks, Morris water maze and passive avoidance tasks. One study did not state subjects’ sex, four studies included female and male subjects and most studies only included males. Six studies used EEG/LFP/PSG measurements. Sodium oxybate was investigated in four of these, increased SWS and/or decreased spindle density were most characteristic.
Results anesthetics.
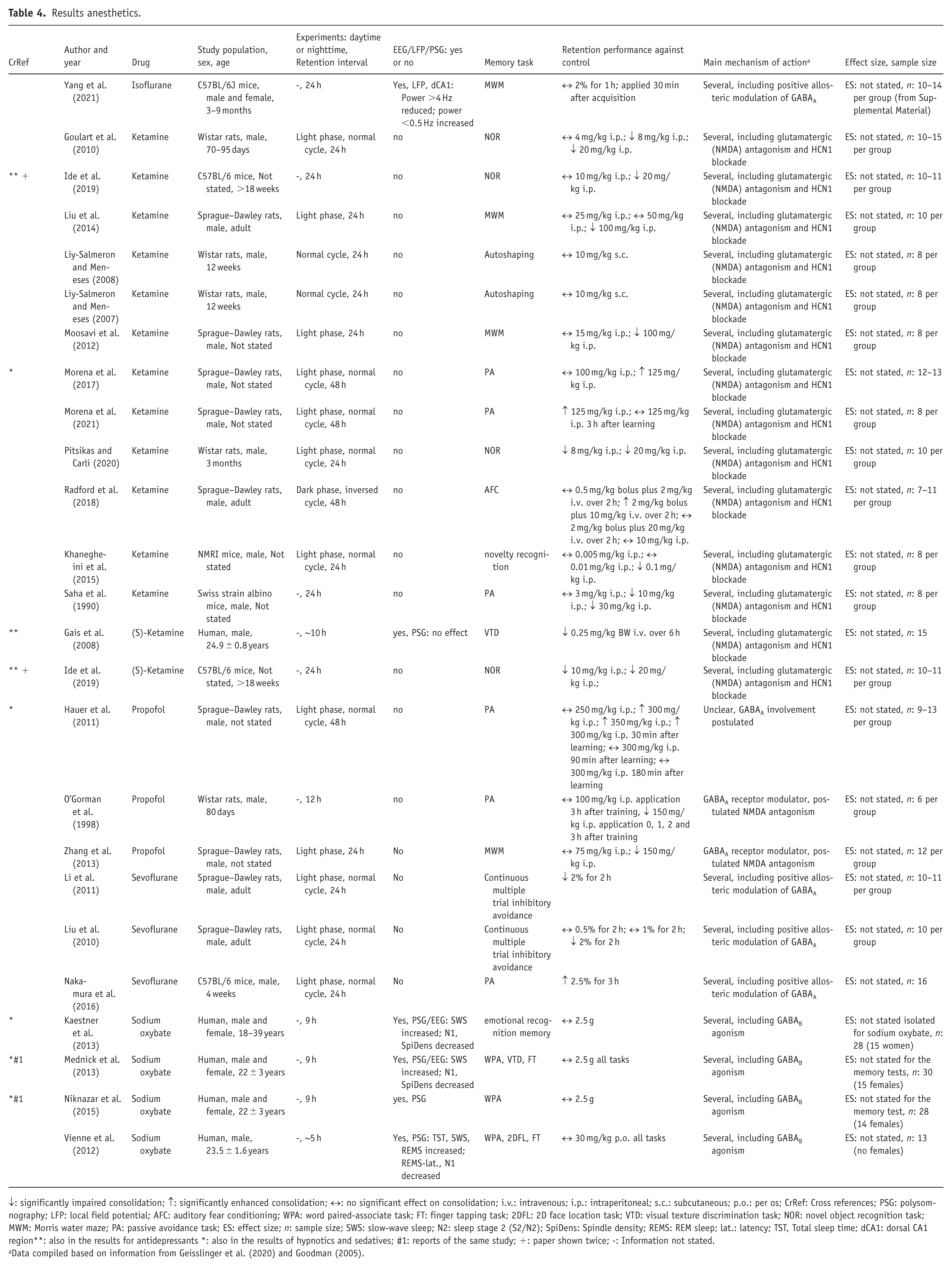
↓: significantly impaired consolidation; ↑: significantly enhanced consolidation; ↔: no significant effect on consolidation; i.v.: intravenous; i.p.: intraperitoneal; s.c.: subcutaneous; p.o.: per os; CrRef: Cross references; PSG: polysomnography; LFP: local field potential; AFC: auditory fear conditioning; WPA: word paired-associate task; FT: finger tapping task; 2DFL: 2D face location task; VTD: visual texture discrimination task; NOR: novel object recognition task; MWM: Morris water maze; PA: passive avoidance task; ES: effect size; n: sample size; SWS: slow-wave sleep; N2: sleep stage 2 (S2/N2); SpiDens: Spindle density; REMS: REM sleep; lat.: latency; TST, Total sleep time; dCA1: dorsal CA1 region**: also in the results for antidepressants *: also in the results of hypnotics and sedatives; #1: reports of the same study; +: paper shown twice; -: Information not stated.
Data compiled based on information from Geisslinger et al. (2020) and Goodman (2005).
Sedatives/hypnotics
For hypnotics and sedatives, 24 reports of 22 studies were included in the analysis and five reports of three studies were subjected to quantitative analysis. The results for all sedatives/hypnotics are shown below in Table 5. Eleven human drug studies and 11 animal drug studies were found. Ten different sedatives/hypnotics were investigated, and the memory tasks included WPA, passive avoidance tasks, auditory fear conditioning, contextual fear conditioning, visual texture discrimination task and finger tapping tasks. One study did not state the subjects’ sex, eight studies included females and males and most studies only included males. Nine studies used EEG/LFP/PSG measurements. Across all human studies measuring EEG/PSG an increment in SWS, N2 or spindle density often concurrent with a decrease in REM sleep or an increase in REM sleep latency were most frequently reported.
Results sedatives/hypnotics.
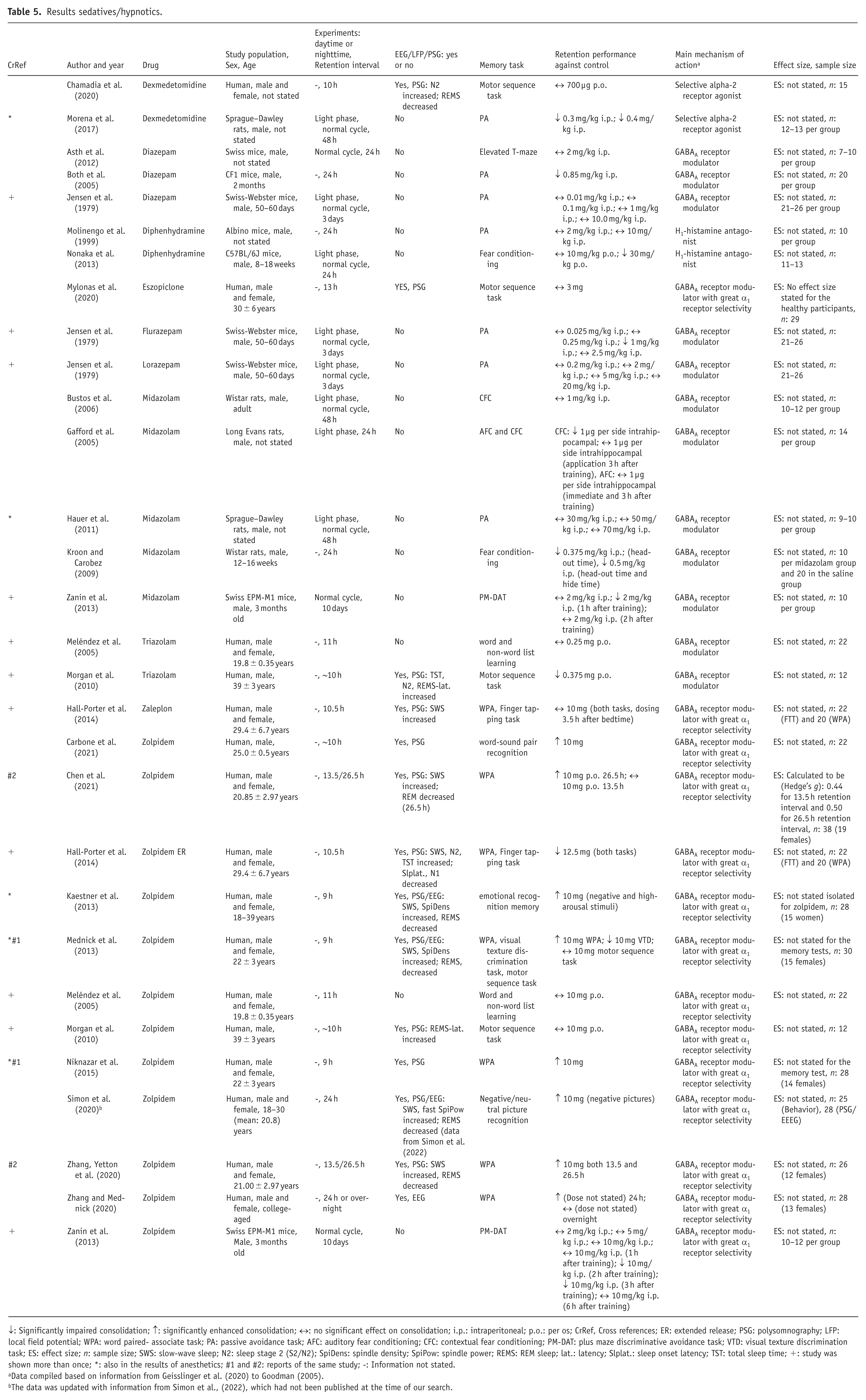
↓: Significantly impaired consolidation; ↑: significantly enhanced consolidation; ↔: no significant effect on consolidation; i.p.: intraperitoneal; p.o.: per os; CrRef, Cross references; ER: extended release; PSG: polysomnography; LFP: local field potential; WPA: word paired- associate task; PA: passive avoidance task; AFC: auditory fear conditioning; CFC: contextual fear conditioning; PM-DAT: plus maze discriminative avoidance task; VTD: visual texture discrimination task; ES: effect size; n: sample size; SWS: slow-wave sleep; N2: sleep stage 2 (S2/N2); SpiDens: spindle density; SpiPow: spindle power; REMS: REM sleep; lat.: latency; Slplat.: sleep onset latency; TST: total sleep time; +: study was shown more than once; *: also in the results of anesthetics; #1 and #2: reports of the same study; -: Information not stated.
Data compiled based on information from Geisslinger et al. (2020) to Goodman (2005).
The data was updated with information from Simon et al., (2022), which had not been published at the time of our search.
Results of quantitative analysis
Five reports on effects of zolpidem on young adults (Chen et al., 2021; Mednick et al., 2013; Niknazar et al., 2015; Zhang and Mednick, 2020; Zhang et al., 2020) were combined in a meta-analysis. Mednick et al. (2013) and Niknazar et al. (2015) as well as Chen et al. (2021) and Zhang et al. (2020) are reports of the same study and combined as one in the statistical analysis (Table 6). One otherwise eligible study was not included in the quantitative analysis because it used zolpidem extended release (ER; Hall-Porter et al., 2014).
Characteristics of studies included in the statistical analysis.
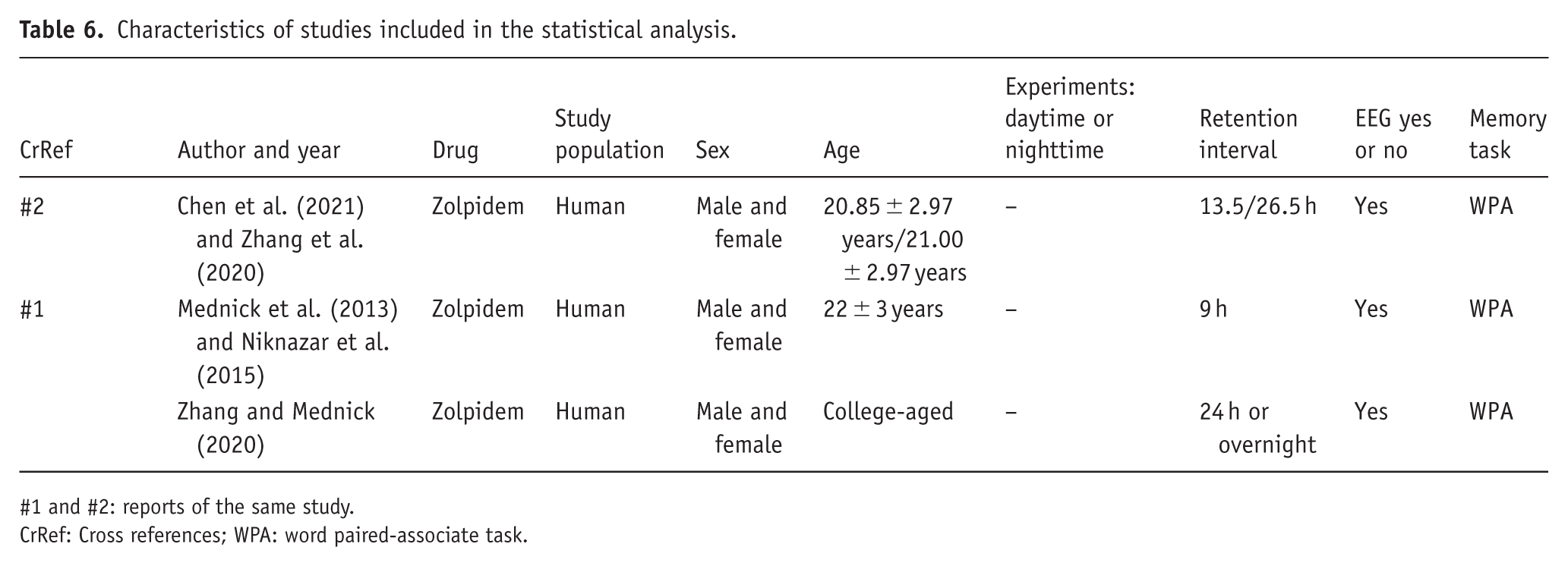
#1 and #2: reports of the same study.
CrRef: Cross references; WPA: word paired-associate task.
The combined effect size estimate shows an effect of zolpidem on consolidation of the WPA of 0.57 (95% CI 0.04–1.10; Figure 2). The CI is wide due to the relatively small overall sample size. Yet, even at the low end of the 95% CI, the effect size still hints at a beneficial effect of zolpidem on memory consolidation compared to placebo, even if small.

Statistical analysis. Forest plot of the effect sizes of the studies on zolpidem. Two of the studies (Chen et al., 2021; Zhang and Mednick, 2020) had two separate retention intervals as indicated by the number of hours of the retention interval or the time of day of testing. The same participant groups were used to establish the different test time points in Zhang and Mednick (2020) and in Chen et al. (2021). Zhang et al. (2020) report on the same data as Chen et al. (2021). We chose the data from Chen et al. (2021) for the meta-analysis since they included a larger number of participants in their statistics.
The heterogeneity was I2 = 58.97% and Tau = 0.10. The PI (–0.45 to 1.60) indicates substantial between-study heterogeneity, suggesting that future studies may find effects ranging from negligible or slightly negative to very large. This might reflect the different retention intervals (from 9 to 26.5 h), different sleep periods (one nap, three nocturnal) and potentially different doses (once not reported).
Discussion
The aim of this systematic review was to provide an objective overview of the influence of specific drug classes on sleep-associated memory consolidation in healthy human adults, rats, and mice. A total of 65 studies were included in the qualitative analysis, reported by drug group first and by individual drug at a second level. Despite differential mechanisms, dose- and species-dependent differences, a comprehensive overview on the effects of these drugs, approved for human use is so far lacking. Enough studies to perform a meta-analysis were only available for the drug zolpidem. The meta-analysis of these three studies on a homogeneous species (=humans) all testing for declarative, hippocampus-dependent memory consolidation in the WPA task found an improvement of memory consolidation.
Antidepressants
Within the drug group antidepressants, a tendency emerged for second-generation antidepressants to enhance memory consolidation as compared to placebo. These included SSRIs (citalopram, escitalopram, fluoxetine, fluvoxamine, sertraline), NARIs (bupropion, reboxetine) and the SNRI duloxetine (lithium, agomelatine, and esketamine do not belong in this pharmacological class). In contrast, first-generation antidepressants, namely tri- and tetracyclic antidepressants (amitriptyline, desipramine, mianserin) either had no effect or impaired memory consolidation, with only one exception.
A benefit of second versus first-generation antidepressants presents an important finding, since a symptom of major depressive disorder is memory impairment (Austin et al., 2001; Beblo et al., 2020). Moreover, for young adults, for whom depression is one of the most common mental health problems and whose brain is still maturing, choice of medication should take potential effects on mnemonic processes and maybe additionally on related structures into account.
The differential effects of first- and second-generation antidepressants, likely reflect the critical difference between these drug generations regarding their affinity to receptors outside of their main mechanism of action. In addition to acting as inhibitors on noradrenergic and serotonergic reuptake transporters they bind to muscarinic, histaminergic, and alpha-adrenergic receptors (Geisslinger et al., 2020; Goodman, 2005) possibly explaining the adverse effects on memory consolidation. For instance, H1-histamine receptor antagonists impair memory consolidation (da Silveira et al., 2013; Nonaka et al., 2013) and histamine receptors in the ventral hippocampus, a relevant hippocampal region for sleep-associated memory consolidation, were found to mediate this impairment (Alvarez and Banzan, 2008; Binder et al., 2019). In addition to acting as a histaminergic antagonist Goerke et al. (2014) argued that impaired consolidation of the procedural task, with amitriptyline in humans was due to the reduced cholinergic tone reflected in reduced REM sleep. In rodents, doses of amitriptyline as used in this study (5 mg/kg i.p.) suffice to strongly suppress REM sleep (Obál et al., 1985). REM sleep reduction was likewise reported here for desipramine (Watts et al., 2012). Whereas REM sleep generation relies significantly on cholinergic activity of the brain stem laterodorsal/ pedunculopontine tegmental nucleus (Wang et al., 2021), norepinephrine and serotonin suppress REM sleep onset and maintenance (Mallick et al., 2008; Monti et al., 2008a). Interestingly, within the included first-generation drugs only consolidation performance of a striatum-dependent task increased (Colavito et al., 2013; Porsolt et al., 1993; Watts et al., 2012).
SSRIs lead to an acute increase in extracellular serotonin levels. Investigating effects of serotonin on declarative memory formation in their recent systematic review (Coray and Quednow, 2022) emphasize discordant effects on consolidation, apparently depending not only on serotonin receptor subtypes, but also on the specific memory task. Cognitive effects were reported to also reveal a bell-shaped dose response curve. Regarding the consolidation step of memory formation, serotonin is reported to support particularly protein synthesis-dependent late phase processes with effects possibly mediated by glutamatergic, GABAergic and/or cholinergic neurotransmission (Seyedabadi et al., 2014). The discrepant findings, that acutely administered SSRIs are frequently reported to impair performance on declarative tasks (Coray and Quednow, 2022), yet in the present review at worst no effect on memory was found, likely reflect brain-state dependent effects of manipulating brain neurochemistry, for example, brain-state dependent effects of increased acetylcholine levels (Gais and Born, 2004; Hasselmo, 1999), and/or on brain electrophysiology that reflect differential network interactions. Improved consolidation of a procedural skill memory task was likely linked to the increased spindle density after both the SSRI fluvoxamine and SNRI reboxetine administrations, found despite REM sleep suppression (Rasch et al., 2009).
Beyond serotonergic mechanisms, activity of the LC has emerged as relevant for sleep-associated processes. In the present study, we reported that the sedative drug dexmedetomidine, a selective alpha-2-receptor agonist, that suppresses noradrenergic activity in the LC (Chiu et al., 1995; Morena et al., 2017; Song et al., 2017), impaired memory consolidation in rats in a passive avoidance task. In humans, oral administration of this drug, also failed to facilitate consolidation of motor task as was found for the placebo condition, although NREM stage N2, and spindle density were enhanced (Chamadia et al., 2020). Pharmacological suppression of noradrenergic transmission by intraperitoneal injection of clonidine and propranolol, decreased not only the occurrence of sleep spindles and slow oscillations, but also their temporal coupling, a relevant electrophysiological signature of memory consolidation during NREM sleep (Durán et al., 2021; Eschenko et al., 2012). Indeed, patterns of LC noradrenergic activity reveal both macro- and micro scale brain-state-associated dynamics that are crystalizing as functionally relevant. In addition to its distinctive NREM and REM sleep-dependent activity, the LC reveals heightened neuronal firing dependent upon phase of the sleep slow oscillation. Omission of LC quiescence during REM sleep as well as prior to spindles through optogenetic stimulation disturbed spindle generation, delta and theta power, as well as the consolidation of spatial memories (Swift et al., 2018). More recently, infraslow oscillations of noradrenergic activity (~0.2 Hz) during NREM sleep were shown relevant for memory consolidation (Kjaerby et al., 2022).
Taken together, the beneficial effect on sleep-associated memory consolidation of antidepressants with mechanisms of action that selectively inhibit reuptake of monoamines, may be attributable to enhanced noradrenergic and/or serotonergic signaling.
Anesthetics
The results in the anesthetics group were mixed for each drug, except for sodium oxybate (n = 4), and isoflurane (n = 1), both of which did not influence memory consolidation in any of the present studies. Although GABAB receptor-mediated activity, through which sodium oxybate mainly exerts it actions, may be required for synaptic plasticity (Heaney and Kinney, 2016; Sanchez-Vives et al., 2021), the present results suggest that acute administration of sodium oxybate at the given systemic concentrations does not affect memory consolidation in young to middle aged adults. To note, EEG recordings during sleep following administration of sodium oxybate, but also of the GABA reuptake inhibitor tiagabine, were likely to induce slow rhythms, but physiological properties of such rhythms, for example, homeostatic SWS response or spindle-related activity appear to diverge from normal sleep (Feld et al., 2013; Kaestner et al., 2013; Mednick et al., 2013; Vienne et al., 2012).
Table 4 reveals that despite different main mechanisms of drug action the five rodent studies revealing increased retention performance over control, employed an aversive task (passive avoidance or acoustic fear conditioning) as well as a relatively long retention interval for their drug (48 hours for ketamine and propofol; 24 hours for sevoflurane). The other listed studies employed neutral tasks, such as spatial tasks or conditioning with neutral stimuli. Together, these findings reflect the relevance of memory content and task for drug effects. Specifically, stimulus salience and relevance of receptors distributed in the amygdala, but also in the hippocampus or nucleus accumbens, that is., structures on which passive avoidance and fear conditioning tasks strongly rely, influence the mnemonic effect (McGaugh, 2000; Meiering et al., 2024; Moustafa et al., 2013).
Dosages and nature of study are furthermore important to consider. The ketamine studies revealing enhanced consolidation used anesthetic-level doses. For subanesthetic doses of ketamine, as used by many rodent studies in Table 4 (⩽30 mg/kg i.p. ketamine; El Iskandrani et al., 2015; Wu et al., 2020) consolidation performance after spatial learning decreased. This applies also to the consolidation of a visual-procedural task in humans. ln this human study, the subanesthetic ketamine administration was initiated at sleep onset and did not modify sleep stage composition (Gais et al., 2008). The contrasting findings on consolidation using subanesthetic versus anesthetic ketamine levels reflect differential dose-dependent effects at neuronal and neural network levels of the non-competitive NMDA-receptor antagonist and are an ongoing subject of investigation (Obert et al., 2024; Zhou and Duan, 2024). Differential effects of ketamine may however, also extend to the influence on other systems, such as the endocanabinoid system, that has been investigated in relation to ketamine’s antidepressant effects (Lang and Borgwardt, 2013; Sharafi et al., 2022; Short et al., 2018).
For propofol, combined findings of the three studies indicate that lower dosages (150 mg/kg i.p.) of propofol in rats as compared to higher dosages inhibit memory consolidation (Hauer et al., 2011; O’Gorman et al., 1998; Zhang et al., 2013), whereby the very lowest dose in each study remained without an effect on consolidation. In mice, dynamic changes in thalamic-cortical rhythmic activity were reported dependent upon anesthesia depth. Studies show that slow waves of natural NREM sleep are not a unitary phenomenon, and that composition of slow-wave types differs with NREM sleep depth (Kim et al., 2019; Seok et al., 2022). It could therefore be that the thalamo-cortical slow-wave activity of propofol-induced deep anesthesia (corresponding in rats to 300 mg/kg BW i.p.; Guang et al., 2025; Murphy et al., 2011) differs qualitatively regarding memory consolidation from the activity of lighter anesthesia depth. Furthermore, temporal factors such as the relationship between retention time to terminal elimination half-life may play a role. Impaired memory retrieval was found at subhypnotic dosages (Liu et al., 2016), and theta activity was suppressed on emergence from anesthesia, possibly affecting retrieval performance (Flores et al., 2017). Yet most importantly Hauer et al. (2011) concluded from further investigations in their study that the memory enhancement was attributed to the elimination by propofol of an endocannabinoid-degrading enzyme (fatty acid amide hydrolase).
For the volatile anesthetics isoflurane and sevoflurane memory effects are mainly attributed to GABAA agonistic activity, most likely mediated by the alpha5 GABAAR subunit that shows a high density in the hippocampus. Differential findings regarding sevoflurane could be due to the heightened aversion associated with the electric shocks of the hippocampus-dependent continuous multiple trial inhibitory avoidance task (Li et al., 2011; Liu et al., 2010). To note, since differences in the minimum alveolar concentration between mice and rats as well as with age are indicated (Cesarovic et al., 2010; Kashimoto et al., 1997) the effects induced by sevoflurane may not be identical between the studies.
Taken together, despite our mixed findings of anesthetics, our results fall in line with others in supporting the need to look beyond classical neurochemicals to explain effects on cognitive performance (Kim et al., 2019; Seok et al., 2022; Zhang et al., 2024).
Sedatives/Hypnotics
Except for dexmedetomidine and diphenylhydramine, all drugs investigated in the included studies act primarily through modulation of the GABAA receptor. Given the central role of GABAergic neurotransmission in the generation of sleep spindles (Gottesmann, 2002; Sanchez-Vives et al., 2021), it could be expected that sedative drugs influence memory consolidation. Consistent with this assumption, optogenetic silencing of GABAergic neurons in the medial septum albeit during REM sleep has been shown to impair contextual memory consolidation, likely through a concomitant reduction in theta oscillations (Boyce et al., 2016). An enhancement in consolidation was however, only found for zolpidem, as revealed in the small meta-analyses. Benzodiazepines either had no effect or impaired memory consolidation across a range of tasks, including passive avoidance, skill learning, and hippocampus-dependent paradigms (corresponding refs from Table 5).
In agreement with above findings for antidepressants the H1-receptor antagonist in this drug group, diphenhydramine, impaired memory consolidation, at least in higher doses. Plausible mechanisms could be disturbed interregional brain communication, as deducted from decreased coupling between brain rhythms of NREM sleep, and/or a suppression of REM sleep-associated long-term potentiation (Masuoka et al., 2019). In Chamadia et al. (2020) time spent in REM sleep was decreased.
Zolpidem belongs to a group of drugs called z-drugs, encompassing also zaleplon, zopiclone and eszopiclone. Unlike benzodiazepines, these compounds exhibit greater selectivity for the α1 subunit of the GABAA receptor. Zaleplon and eszopiclone were each examined in one study included in this review, but neither was associated with improved memory consolidation. In the case of zaleplon, the tasks employed were comparable to those used in the zolpidem study; however, chronopharmacological differences complicate comparisons. Specifically, zaleplon was administered 3.5 hours after bedtime, whereas zolpidem was given 30 minutes before bedtime in an ER formulation (Hall-Porter et al., 2014). In that study, zolpidem-ER was found to impair memory consolidation relative to both placebo and zaleplon, while no difference was observed between zaleplon and placebo. Notably, to equate experimental conditions, participants receiving zolpidem-ER were also awakened 3.5 hours after bedtime, a manipulation that may itself have disrupted memory consolidation compared to undisturbed sleep. The study using the ER formula (Hall-Porter et al., 2014) was therefore excluded from the meta-analysis.
Benzodiazepines, the other major group of GABAA modulators, had either no effect, or impaired memory consolidation for diverse tasks involving passive avoidance, skill learning and hippocampus-dependent tasks (Table 5). Similar to findings of the one human study administering a benzodiazepine here in which light NREM sleep was enhanced (Morgan et al., 2010), Leong et al. (2022) report in their review enhanced spindle activity across most studies with benzodiazepines. In part, benzodiazepines even revealed greater enhancements of spindle activity than z-drugs (Leong et al., 2022). These findings show that spindle activity of light NREM sleep is not always a reliable correlate for enhanced memory consolidation.
Note that both benzodiazepines and z-drugs have clinically been associated with cognitive impairment in long-term use (Atkin et al., 2018). But as far as the authors know, there is no information on spindle activity with long-term usage of these drugs. More recently, zolpidem was reported to enhance the temporal coordination between slow oscillations and sleep spindles, as well as hippocampal–prefrontal temporal coupling -electrophysiological processes that are closely linked to memory consolidation (Kersanté et al., 2023). Future research will be required to determine whether these effects, relative to benzodiazepines, arise from the higher receptor subunit specificity of zolpidem, differences in regional receptor distribution, or alternative mechanisms.
Despite its beneficial effect on memory consolidation, zolpidem was recently reported to suppress the vasomotor activity relevant for glymphatic clearance by suppressing the noradrenergic infraslow oscillation and associated micro-arousals (Hauglund et al., 2025). It is yet to be investigated, whether the likelihood of neurodegenerative disorders including memory disorders such as Alzheimer’s disease are heightened by (long-term) use of medications (Heneka et al., 2010; Yu et al., 2025).
In summary, the results underscore the concept, that (pharmacologically induced) increased spindle activity generated by increased GABAergic signaling alone does not facilitate memory consolidation, but rather that the interaction with other sleep rhythms such as the sleep slow oscillation is essential (Kersanté et al., 2023; Staresina, 2024).
Limitations
Mechanisms of sleep-associated memory consolidation are still a matter of intense research with studies showing a significant dependence on temporal aspects, such as time between learning and sleep duration of post-learning retention interval of sleep the composition of ongoing brain rhythms in sleep (Deng et al., 2025; Diekelmann and Born, 2010; Eschenko et al., 2006; Poe et al., 2010; Rasch and Born, 2013), time of day per se and neuroendocrine interactions (Feld and Born, 2020; Lazar et al., 2015; Thorn et al., 2004; van der Zwet et al., 2025), and their interactions with the type of memory task (Marshall et al., 2020; Myhrer, 2003). On the other hand, most studies on the investigated pharmacological drug classes and memory tasks have not focused on these aspects. Due to the paucity of corresponding studies, neither polysomnography nor brain electrical activity could be used as inclusion criteria. For the same reason, but also due to the diversity of drugs, the review as a whole did not discriminate between types of memory, as hippocampus-dependent versus non-hippocampus-dependent. Only for a few drugs did literature allow for a discussion on concordant brain electrical activity and underlying mechanisms. In the present review, task-dependent effects for ketamine were apparent: Two studies reported an impairment in memory consolidation at the highest dose using the Morris water maze (Liu et al., 2014; Moosavi et al., 2012), while an improvement in consolidation of a passive avoidance task similarly using a regimen with the highest ketamine dose (Morena et al., 2017). Studies have also shown that, in addition to species-specific differences in drug responses, strain-dependent differences in memory consolidation occur, for example, for dopaminergic drugs (Gasbarri et al., 1997). These aspects further hamper any a priori generalization of the present findings. Importantly, new and distinct mechanisms of action for the investigated drug classes are still being disclosed, suggesting the necessity of more drug-specific investigations of their cognitive impacts during both wakefulness and sleep.
A further limitation is the relatively low number of participants, only reaching 13 in some human studies. Moreover, the participants are exclusively healthy, with no representation from those suffering from disease. Also, age ranges are not strictly limited to young adults. Consequently, the findings of the study are unable to provide any insights into the effects of the drugs in patients. As indicated above, the number of studies eligible for inclusion in the meta-analysis was suboptimal, leading to a wide CI. A placebo-controlled design was part of the inclusion criteria. All but one study with human subjects reported blinding of investigators. However, the reviewed animal studies present a contrasting picture; blinding is scarcely reported. Since the latter, especially during analyses is relevant to ensure unbiased results, mention in publications should become the norm.
Conclusion
Investigating the effect drugs have on memory consolidation not only helps to further characterize the drugs, but it also supports the understanding of the neurophysiology of the formation of memory. The drugs included in this review are a part of clinical prescribing in a large scale. While direct conclusions from this study for clinical use remain limited, results may provide a foundation for future considerations regarding effects on memory consolidation. For instance, second-generation antidepressants compared to first-generation antidepressants appear to be the favorable choice. Overcoming the limitations of this study, future research should include a wider scope of drug classes including off-label usage, larger populations, and have a more balanced participation of sexes and in later studies, include patients as opposed to healthy populations. Most of the included human studies investigated young adults. For future advancements on the assessment of drug actions on sleep-associated memory consolidation, we emphasize including concordant electrophysiological measures during the consolidation period, to enable estimations of quality/state of sleep or wakefulness, sleep rhythms as well as additional measures of brain electrical activity, potentially in combination with imaging techniques. Animal studies would be strengthened by more thorough and transparent reporting of potential risks of bias. In addition, findings should generally be evaluated alongside those of continuously advancing non-pharmacological approaches, such as weak non-invasive brain stimulation.
Supplemental Material
sj-docx-1-jop-10.1177_02698811261456185 – Supplemental material for Effects of antidepressants, anesthetics and sedatives/hypnotics on sleep-associated memory consolidation – A systematic review
Supplemental material, sj-docx-1-jop-10.1177_02698811261456185 for Effects of antidepressants, anesthetics and sedatives/hypnotics on sleep-associated memory consolidation – A systematic review by Janina Mischke, Kerstin Luedtke and Lisa Marshall in Journal of Psychopharmacology
Footnotes
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: DFG (MA2053/11-1).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Data is available upon reasonable request.
Pre registration
This review was registered at PROSPERO (registration number CRD42022293271).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.