
Abstract
Objective
To examine the association between the percentage of peripheral natural killer cells and thyroid autoimmunity in women with a history of miscarriage and assess whether B-cells mediate this association.
Methods
This retrospective cross-sectional study was conducted on 927 women with a history of miscarriage treated at the Affiliated Hospital of Guizhou Medical University (2021–2023). Peripheral T-cell, B-cell, and natural killer cell subsets and thyroid antibodies were assessed. Multivariable logistic regression, restricted cubic splines, and mediation analyses were performed.
Results
Women with thyroid autoimmunity had lower percentage of peripheral natural killer cells and higher percentage of B-cells than those without thyroid autoimmunity (both p < 0.001). The percentage of peripheral natural killer cells demonstrated a consistent inverse association with thyroid autoimmunity after adjustment (adjusted odds ratios: 0.00; 95% confidence interval: 0.00–0.02; all p < 0.001). Restricted cubic spline analysis indicated a nonlinear dose–response relationship that was stronger in women aged ≥35 years and in those without recurrent miscarriages. B-cells partially mediated the association between the percentage of peripheral natural killer cells and thyroid autoimmunity (31.4% of the total effect; ab = −0.166; p < 0.001).
Conclusion
Lower percentage of peripheral natural killer cells is linked to higher odds of thyroid autoimmunity in women with a history of miscarriage, and B-cells partly explain this relationship, supporting the use of peripheral immune profiling to identify thyroid autoimmunity risk.
Introduction
Thyroid autoimmunity (TAI) is a common autoimmune disorder that is characterized by the presence of antithyroid peroxidase (anti-TPO) and/or antithyroglobulin (anti-TG) antibodies. TAI is recognized as an important factor affecting female reproductive health, particularly the health of women with a history of miscarriage.1,2 Although the exact mechanism remains debated, autoantibodies may contribute to adverse outcomes by inducing complement activation at the placental interface. These disorders not only compromise the physical health of affected individuals but also exert negative psychological impacts on them. Moreover, these disorders impose a substantial burden on the healthcare systems and societal resources. 3
Natural killer (NK) cells are large granular lymphocytes that play a central role in innate immunity. Mature peripheral NK cells (pNKs) in the blood account for approximately 5%–20% of all circulating lymphocytes and constitute a critical component of the body’s first line of immune defense. 4 NK cells have attracted increasing attention owing to their dual regulatory roles, balancing tissue remodeling via angiogenic factors (e.g. vascular endothelial growth factor (VEGF) and placental growth factor (PLGF)) and immune surveillance through cytotoxic granule release. 5 Additionally, NK cells reportedly regulate immune responses through the secretion of interferon-gamma (IFN-γ) and tumor necrosis factor-alpha (TNF-α) and via Fas ligand (FasL)–mediated cytotoxicity. 5 TAI is driven primarily by aberrant activation of T-cells and B-cells. 6 However, accumulating evidence suggests that NK cells, serving as a bridge between innate and adaptive immunity, play critical roles in the onset and progression of TAI by infiltrating thyroid tissue and inducing follicular cell apoptosis via the perforin–granzyme pathway. 7 However, the relationship between pNK counts and TAI remains to be sufficiently explored in clinical settings. Most of the existing studies have focused on populations with Graves' disease (GD) or Hashimoto’s thyroiditis (HT),8,9 and only a few have investigated this association in women with a history of miscarriage. Immune dysregulation may compromise maternal immune tolerance, disrupt embryo implantation or pregnancy maintenance, and ultimately increase the risk of miscarriage. 10 One study has reported a positive correlation between TAI and elevated pNK activity in women with recurrent miscarriage (RM), suggesting that the use of the percentage of peripheral natural killer cells (pNK%) as a biomarker for immunotherapy in this context should be approached with caution. 11
B-cells play a central role in the pathogenesis and progression of TAI. These cells act as the primary source of thyroid-specific autoantibodies such as thyroid peroxidase antibody (TPOAb) and thyroglobulin antibody (TgAb). The activation and aberrant proliferation of B-cells are considered key mechanisms underlying the development of TAI.12,13 NK cells have been reported to contribute to TAI pathogenesis through infiltration of the thyroid tissue and follicular cell apoptosis, mechanisms supported primarily by experimental and animal studies. Their potential immunoregulatory role in modulating B-cell responses, possibly through cytokine networks and antibody-dependent cellular cytotoxicity (ADCC), has also been described; 14 however, the precise mechanisms in human peripheral blood and whether changes in NK cell abundance influence TAI development via modulation of B-cell function remain to be elucidated. Emerging evidence suggests that the crosstalk between innate and adaptive immune cells shapes the autoimmune responses in the thyroid microenvironment.15,16 In this context, elucidating the dynamic interaction between NK and B-cells may provide novel insights into the immunopathogenesis of TAI.
However, only a few large-scale studies have specifically investigated the association between NK cells and TAI in women with a history of miscarriage; in particular, studies incorporating nonlinear analyses and mediation modeling are scarce. Therefore, based on the clinical data of outpatient women with a history of miscarriage at the Affiliated Hospital of Guizhou Medical University, this study aimed to examine the strength of the association between the pNK% and TAI, assess the potential nonlinear dose–response relationships, and evaluate the mediating effect of B-cells in this association. The objective was to elucidate the potential immunoregulatory role of the peripheral immune status in the development of TAI and provide a theoretical basis for early identification and intervention in high-risk populations.
Materials and methods
Study population
This retrospective analysis included all consecutive female patients with a history of miscarriage who visited the Reproductive Medicine Center of the Affiliated Hospital of Guizhou Medical University between January 2021 and December 2023 and fulfilled the inclusion criteria. No selective sampling was performed. Electronic medical records were retrospectively reviewed, and a total of 1530 records were initially identified. After excluding records with missing TAI data (n = 186), incomplete covariate information (n = 320), and no confirmed history of miscarriage (n = 97), a total of 927 participants were included in the final analysis. The study was approved by the Ethics Committee of the Affiliated Hospital of Guizhou Medical University, Guiyang, China (Approval Number: 2025 Ethics Review No. 284; Approval Date: 24 June 2025). All patient data were deidentified prior to analysis to protect patient privacy and confidentiality. No individual patient information that could lead to identification is presented in this manuscript. The study was conducted in accordance with the Declaration of Helsinki (1975, revised in 2024).
Reporting guidelines
The reporting of this cross-sectional study conforms to the Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) guidelines for observational studies. 17
Data collection
The data collection process consisted of two components: (a) medical history and (b) measurement of laboratory parameters. The medical history included information on age and number of miscarriages, whereas the laboratory parameters data collection included T-cell, B-cell, and NK cell (TBNK) lymphocyte subsets (both absolute and relative counts), thyroid autoantibodies, three infertility-related autoantibodies (anticardiolipin, antiendometrial, and antisperm antibodies), and a full panel of antinuclear antibodies. All blood samples were collected during the nonmenstrual phase of the cycle. However, specific cycle days (follicular vs. luteal) were not strictly controlled or recorded for all participants.
TBNK lymphocyte subset detection
A total of 50 μL of the tripotassium ethylenediaminetetraacetic acid (K3EDTA)–anticoagulated whole blood sample was mixed with 10 μL of a labeled antibody cocktail (Multitest 6-color TBNK reagent kit: CD3 fluorescein isothiocyanate (FITC)/CD16 phycoerythrin (PE) + CD56 PE/CD45 peridinin-chlorophyll protein–cyanine 5.5 (PerCP-Cy5.5)/CD4 phycoerythrin–cyanine 7 (PE-Cy7)/CD19 allophycocynanin (APC)/CD8 allophycocyanin–cyanine 7 (APC-Cy7); Becton Dickinson). The mixture was then incubated at room temperature in the dark for 20 min. Subsequently, 1 mL of the lysing solution (Easy-Lyse, DAKO) was added, and the sample was incubated for an additional 10 min at 4°C–8°C. After incubation, the sample was immediately prepared for analysis. Flow cytometric analysis was performed using BD FACSCanto II and FACS Diva 2.0 software.
Thyroid autoantibody detection
A 3-mL peripheral venous blood sample was collected from each participant into a serum separator tube (SST) containing separation gel. The sample was allowed to clot at room temperature for 30 min and then centrifuged at 4000 r/min (approximately 895 ×g; centrifugal radius, 5 cm) for 10 min. The serum was carefully separated and stored at −20°C until analysis. TgAb and TPOAb levels were determined within 4 h of sample collection using a fully automated dual-probe radioimmunoassay analyzer (Model CN202M/KZ4GC-1200, Beijing Haifuda Technology Co. Ltd.), along with the corresponding commercial TPOAb and TgAb reagent kits.
Thyroid autoantibodies were also determined using a fully automated chemiluminescence immunoassay analyzer (CL-6000i, Mindray Biomedical Electronics Co., Ltd.; Shenzhen, China). Additional analysis of the TPOAb and TgAb levels was conducted using an IM-8000 imaging measurement system (IM-8000, Keyence China Co., Ltd.), along with the corresponding commercial reagent kits. All procedures were performed strictly according to the manufacturers’ protocols provided for TPOAb and TgAb. If the antibody concentration exceeded the upper detection limit, the serum sample was diluted at a ratio of 1:10 to 1:20 and then retested.
Detection of antiendometrial, anticardiolipin, and antisperm antibodies
A 3-mL peripheral venous blood sample was collected from each participant, and the serum was isolated through centrifugation. The presence of antiendometrial, anticardiolipin, and antisperm antibodies was assessed using immunogold colloidal assay kits (Sanming Bofeng Biotechnology Co. Ltd.; China). The test results were interpreted according to the appearance of red lines: the presence of both control and test lines indicated a positive result; the appearance of only the control line indicated a negative result; absence of the control line or presence of the test line alone was considered to indicate invalid results.
Antinuclear antibody (ANA) panel detection
The ANA panel (immunoglobulin G (IgG) type) was assessed using an immunoblot assay kit (Euroimmun, Hangzhou Medical Diagnostic Co. Ltd.; China) following the manufacturer’s instructions. The detection kit (Model: ANA Profile 1) enables the simultaneous detection of antibodies against a panel of autoantigens, including nuclear ribonucleoprotein/Smith antigen (nRNP/Sm), Smith antigen (Sm), Sjögren’s-syndrome-related antigen A (Ro) (SS-A) (native and Ro-52), Sjögren’s-syndrome-related antigen B (La) (SS-B), topoisomerase I scleroderma-70 antigen (Scl-70), histidyl-tRNA synthetase (Jo-1), centromere protein B (CENP B), double-stranded DNA (dsDNA), nucleosome, histone, and ribosomal P protein. Serum or anticoagulated plasma samples (EDTA, heparin, or citrate) were diluted in sample buffer at a ratio of 1:100 and then incubated with the antigen-coated membrane strips. After washing, alkaline phosphatase–conjugated goat anti-human IgG was added as the secondary antibody. The substrate nitro blue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate (NBT/BCIP) was applied for colorimetric detection. All reagents were equilibrated to room temperature (18°C–25°C) for 30 min prior to use. The diluted samples and positive controls were freshly prepared and used within the same working day. Serum samples were stored at 2°C–8°C and tested within 14 days of sample collection.
Definition of covariates
The covariates included in this study were CD3+ T%; CD3+CD8+T%; age; number of miscarriages; antisperm, antiendometrial, and anticardiolipin antibodies; and a comprehensive panel of both nuclear and cytoplasmic autoantibodies. The nuclear autoantibodies consisted of ANA, anti-Scl-70, anti-SSA, anti-histone, anti–proliferating cell nuclear antigen antibody (anti-PCNA), anti-CENP-B, anti-Sm/nRNP, anti-Ro-52, anti-SSB, anti-dsDNA, anti-Sm, anti–polymyositis/scleroderma antibody (anti-PM-Scl), and antinucleosome antibodies. The cytoplasmic autoantibodies included antimitochondrial antibodies, antiribosomal P protein antibodies, and anti-Jo-1 antibodies. 18
Miscarriage frequency was defined according to the relevant clinical standards in China as follows: recurrent spontaneous abortion (RSA) was defined as ≥3 consecutive spontaneous miscarriages, each occurring before 28 weeks of gestation and involving a fetus weighing <1000 g. In this study, women with ≤2 miscarriages were categorized into the “miscarriage group,” whereas those with ≥3 miscarriages were classified into the “RSA group.”
Outcome variable
TAI status was defined as positive when the serum anti-TPOAb concentrations were ≥34 IU/mL and/or the serum anti-TgAb concentrations were ≥115 IU/mL. Participants not meeting either threshold were classified as TAI negative. 19
Statistical analyses
Quantitative variables that did not follow a normal distribution were expressed as medians and interquartile ranges (IQRs) and were analyzed using nonparametric tests. Categorical variables were summarized as counts and percentages and were compared using the chi-square test. To examine the associations between the pNK% and TAI, binary logistic regression models were employed. Effect estimates were reported as odds ratios (ORs) with 95% confidence intervals (CIs). A restricted cubic spline (RCS) model was adopted to explore the potential nonlinear relationship between pNK% and TAI, and dose–response analysis was conducted to further characterize the association. Subgroup analyses were performed stratified by age (<35 vs. ≥35 years) and number of miscarriages (<3 vs. ≥3) to assess the consistency across strata. To investigate potential mediation pathways, mediation analysis was conducted using the ‘bruceR’ package in R, which is based on Model 4 of the PROCESS macro developed by Hayes (i.e. a simple mediation model). The B-cell percentage (B%) was considered a potential mediator of the relationship between pNK% and TAI. The significance of the mediation effect was evaluated using a bootstrap approach with 10,000 resamples; a 95% bootstrap CI not containing zero was considered evidence of a statistically significant mediation effect. All models were adjusted for potential confounders, including age (years), cluster of differentiation (CD) CD3+ T%, CD3+CD8+T%, number of miscarriages, antisperm antibody, antiendometrial antibody, anticardiolipin antibody, nuclear autoantibodies, and cytoplasmic autoantibodies. Variance inflation factors (VIFs) were calculated to determine the multicollinearity, with a VIF >5 indicating potential collinearity issues. Normally distributed continuous variables were compared using independent (unpaired) Student’s t-test and non-normally distributed ones using the Mann–Whitney U test.
Covariates were selected based on biological plausibility, prior literature, and potential confounding. Age and miscarriage history were included as TAI risk factors; CD3+ T% and CD3+ CD8+ T% to reflect lymphocyte composition; and autoantibodies as indicators of systemic autoimmune activity. All VIFs were <5.
All the statistical analyses were performed using R software (version 4.3.0) and Jamovi (version 2.4.1).
Results
Baseline characteristics of the study population
In total, 927 participants were included in the final analysis, with 297 in the TAI-positive group and 630 in the TAI-negative group. Differences in the immune cell subsets and autoantibody distributions between the two groups are presented in Table 1.
Baseline characteristics of the study population stratified by the TAI status (n = 927).
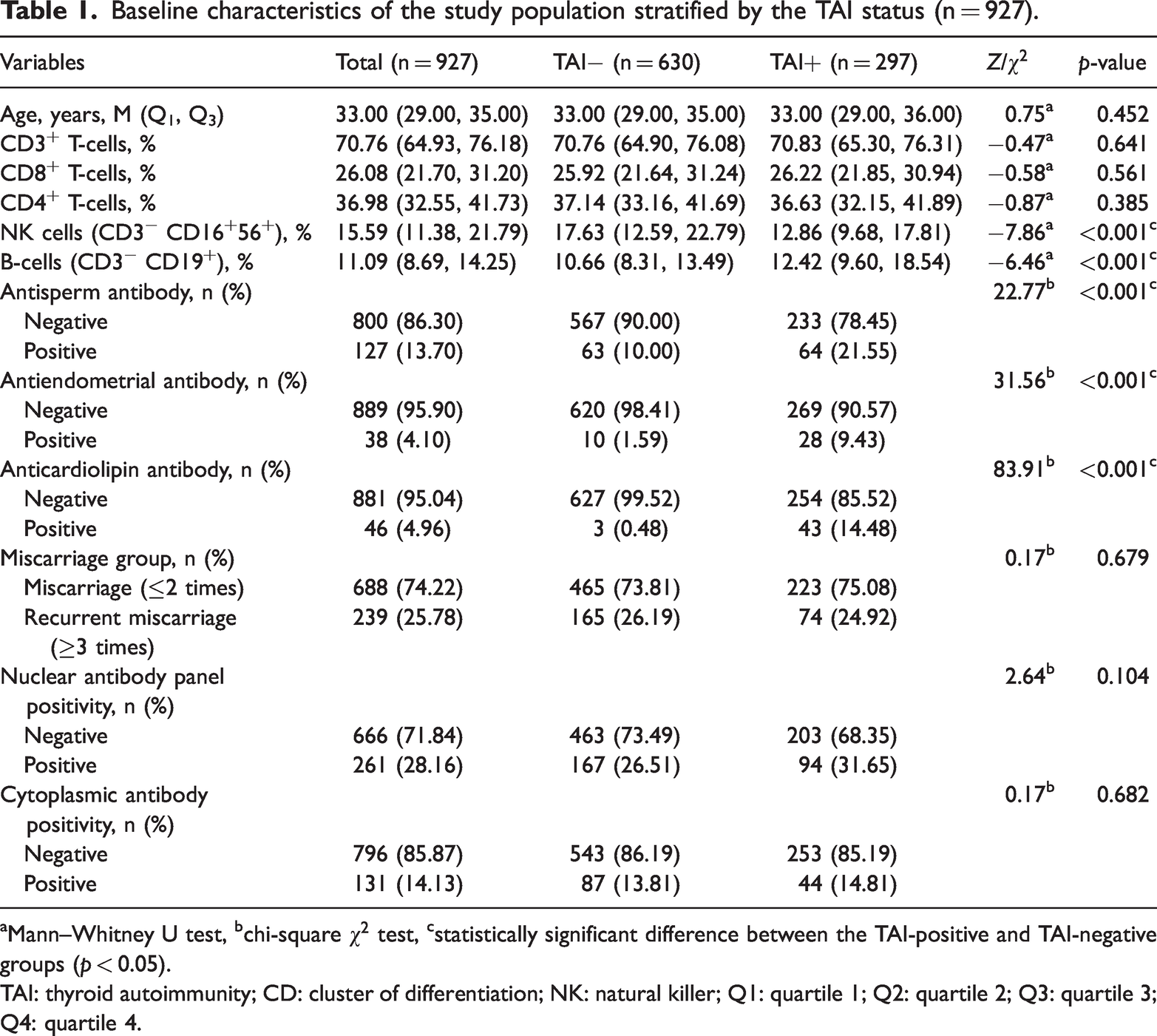
Mann–Whitney U test, bchi-square
TAI: thyroid autoimmunity; CD: cluster of differentiation; NK: natural killer; Q1: quartile 1; Q2: quartile 2; Q3: quartile 3; Q4: quartile 4.
Compared with the TAI-negative group, the TAI-positive group presented a significantly lower pNK% and a significantly greater B% (both p < 0.001). In terms of autoantibodies, the positivity rates for antisperm, antiendometrial, and anticardiolipin antibodies were significantly greater in the TAI-positive group (all p < 0.001). However, no significant differences were detected between the groups in terms of cytoplasmic antibody positivity or the proportion of participants with RSA (all p > 0.05).
Association between thyroid autoantibodies and pNK%
Logistic regression analyses revealed a consistent and significant association between the pNK% and TAI across all models (Table 2). According to the unadjusted model (Model 1), the OR for pNK% was 0.00 (95% CI: 0.00, 0.01, p < 0.001). After adjusting for age and number of miscarriages (Model 2), the association remained significant, with an OR of 0.00 (95% CI: 0.00, 0.01, p < 0.001). In the fully adjusted model (Model 3), in which additional control for CD3+T%, CD3+CD8+T%, antisperm antibodies, antiendometrial antibodies, anticardiolipin antibodies, nuclear autoantibodies, and cytoplasmic autoantibodies was applied, the association persisted (OR = 0.00, 95% CI: 0.00, 0.02; p < 0.001).
Associations between the pNK% and TAI according to the logistic regression models.
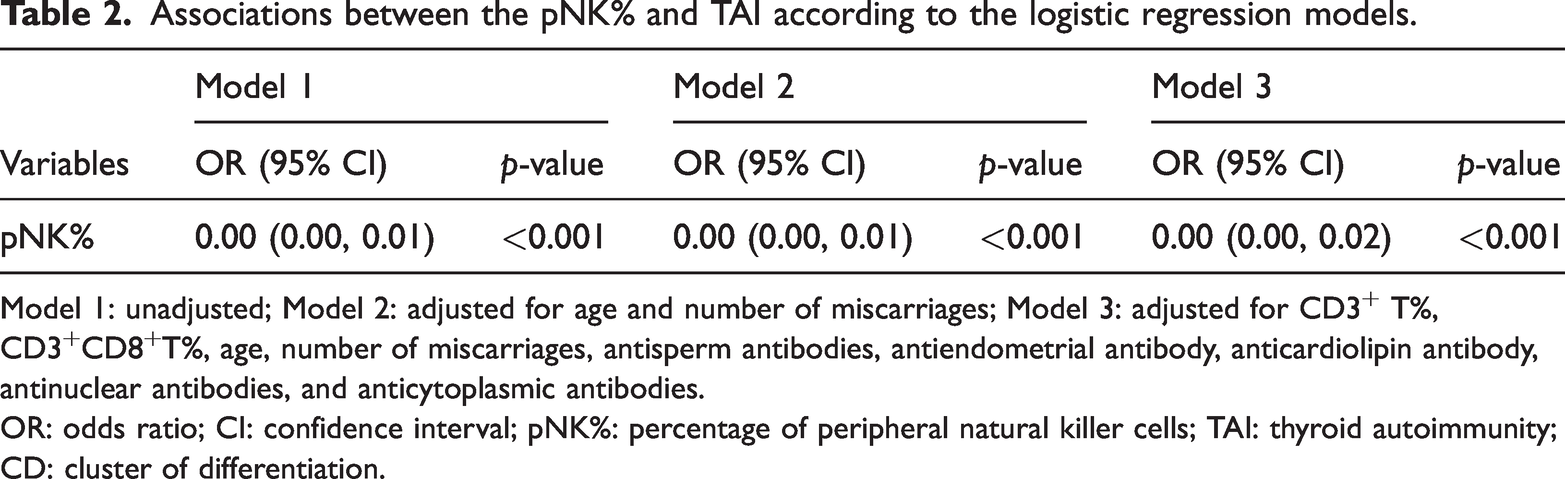
Model 1: unadjusted; Model 2: adjusted for age and number of miscarriages; Model 3: adjusted for CD3+ T%, CD3+CD8+T%, age, number of miscarriages, antisperm antibodies, antiendometrial antibody, anticardiolipin antibody, antinuclear antibodies, and anticytoplasmic antibodies.
OR: odds ratio; CI: confidence interval; pNK%: percentage of peripheral natural killer cells; TAI: thyroid autoimmunity; CD: cluster of differentiation.
These findings suggested a robust inverse relationship between the pNK% and presence of TAI.
Dose–response relationship between thyroid autoantibodies and pNK%
As shown in Figure 1, the RCS analysis revealed a significant dose–response relationship between the pNK% and TAI. In the overall population, a statistically significant nonlinear association was observed (p for overall trend < 0.001; p for nonlinearity = 0.003), suggesting that the relationship between pNK% and the TAI was not simply linear.
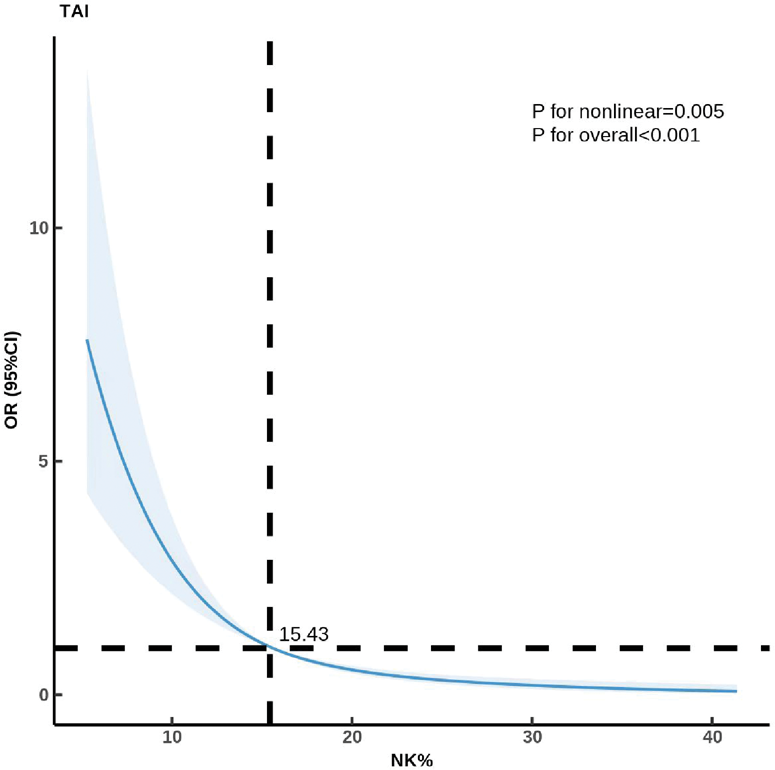
Nonlinear dose–response relationship between the percentage of peripheral NK cells and the risk of thyroid autoimmunity (TAI) in the overall population. NK: natural killer; NK%: percentage of peripheral natural killer cells.
Stratified analysis
As shown in Figure 2, in the age-stratified analysis, a significant nonlinear relationship between the pNK% and TAI was observed among women aged ≥35 years (advanced maternal age (AMA) group; p for overall trend <0.001; p for nonlinearity = 0.001). In contrast, women aged <35 years (non-AMA group) presented a linear association (p for overall trend <0.001; p for nonlinearity = 0.342). Similarly, in the analysis stratified by miscarriage history, a significant nonlinear association was identified in women with <3 miscarriages (No-RSA group; p for overall trend <0.001; p for nonlinearity = 0.004). However, among women with RSA (RSA group, ≥3 miscarriages), the relationship was linear (p for overall trend = 0.004; p for nonlinearity = 0.380). These findings suggest that the impact of the pNK% on TAI risk follows distinct dose–response patterns across subgroups defined by age and reproductive history.
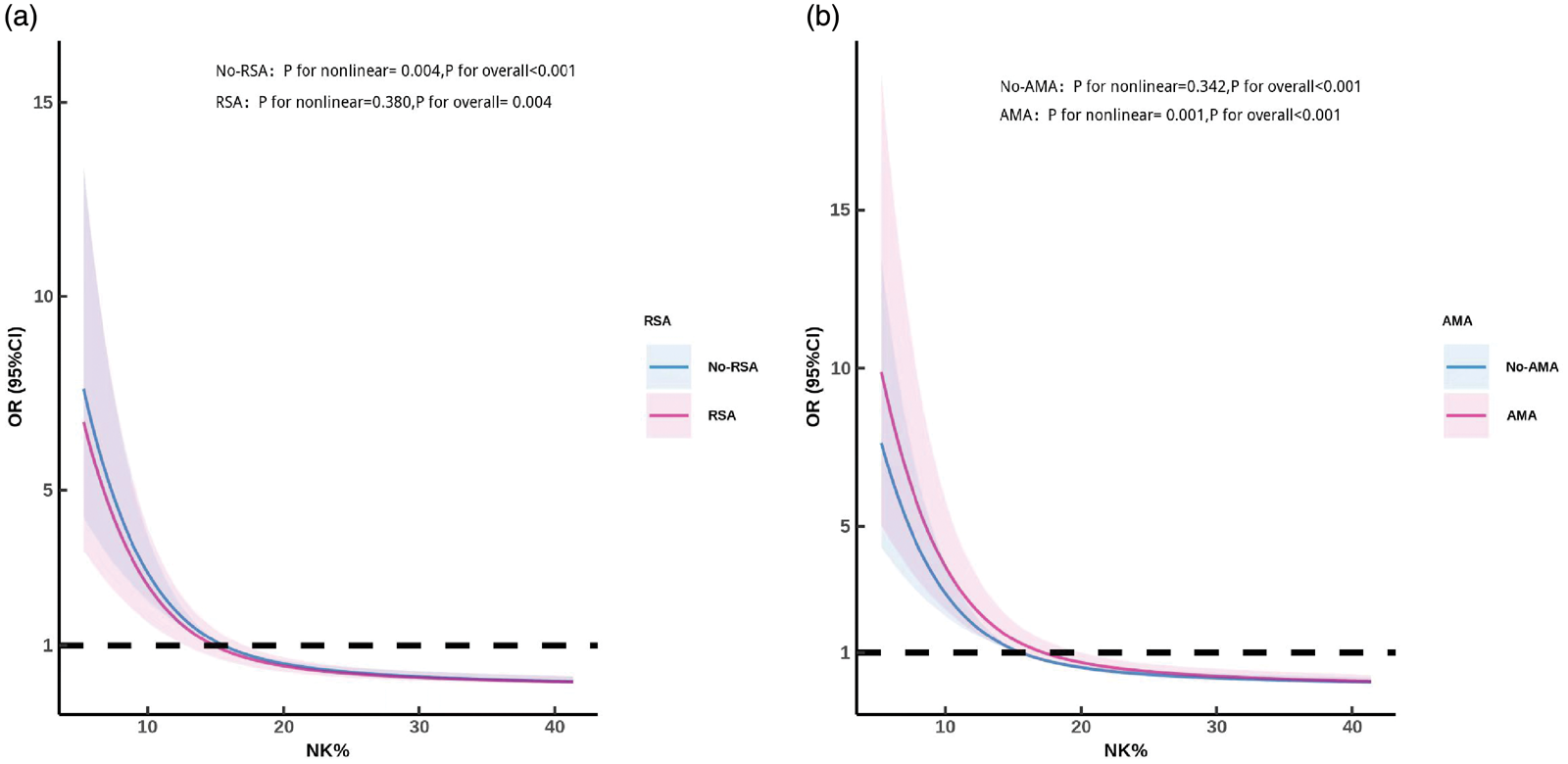
Stratified dose–response relationships between the peripheral NK cell percentage (pNK%) and risk of thyroid autoimmunity (TAI) (a) stratified by the history of recurrent spontaneous abortion (RSA) and (b) stratified by age (<35 and ≥35 years). Women aged ≥35 years were classified as having advanced maternal age (AMA). NK: natural killer; NK%: percentage of peripheral natural killer cells.
Mediation analysis
As shown in Table 3, mediation analysis revealed that the pNK% exerted a significant negative total effect on TAI (β = −0.528, standard error (SE) = 0.058, 95% CI (−0.619, −0.401), p < 0.001). Notably, this effect was partially mediated through B%. The indirect effect via B% was statistically significant (β = −0.166, SE = 0.042, 95% CI (−0.248, −0.081), p < 0.001), suggesting that the pNK% partially influences TAI by modulating B-cell levels. The direct effect of the pNK% on TAI remained significant after controlling for B% (β = −0.362, SE = 0.069, 95% CI (−0.473, −0.214), p < 0.001), consistent with a partial mediation model. These findings indicate that NK cells influence TAI through both direct and indirect mechanisms involving B-cells, underscoring a potential immunoregulatory pathway that warrants further investigation. These findings indicate that B-cells partially mediated the association between the pNK% and TAI, accounting for approximately 31.4% of the total effect (mediation proportion =−0.166/−0.528 ≈ 31.4%).
Analysis of the effect of the percentage of peripheral NK cells on TAI mediated by B-cells.

SE: standard error; NK: natural killer; TAI: thyroid autoimmunity.
Discussion
In this study, involving 927 women with a history of miscarriage from the Affiliated Hospital of Guizhou Medical University, the associations between peripheral immune cell subsets, specifically, the pNK% and B%, and TAI were investigated. The findings revealed that individuals with TAI had significantly lower pNK% and higher B% than those without TAI, and this association remained robust after adjustment for age, number of miscarriages, and various autoantibodies (antisperm, antiendometrial, anticardiolipin, nuclear, and cytoplasmic antibodies). Further analyses using RCS demonstrated a nonlinear dose–response relationship between the pNK% and TAI, whereas the mediation analysis indicated that B-cells partially mediated this association. These results provide novel insights into the immune mechanisms underlying TAI in women of reproductive age and suggest an association between pNK–B-cell interactions and TAI.
NK cells were traditionally regarded as a component of the innate immune system, playing a pivotal role in antiviral and antitumor immunity by recognizing the “non-self” epitopes and inducing target cell lysis. 20 However, recent studies have recognized that NK cells also participate in the regulation of adaptive immunity, playing critical roles in complex immunological contexts, such as autoimmunity and maternal immune tolerance during pregnancy. 21 This study revealed a significant decrease in the pNK% and a corresponding increase in the B% in the TAI-positive group. This finding is consistent with previous studies that have reported immune dysregulation in various autoimmune diseases, such as multiple sclerosis (MS), 22 systemic lupus erythematosus (SLE), 23 Sjögren’s syndrome (SS), 24 rheumatoid arthritis (RA), 25 and type 1 diabetes mellitus (T1DM). 26 Patients with autoimmune thyroid disease often exhibit reduced numbers and impaired function of pNKs. Recent studies have demonstrated that deficiencies in NK cells, either in terms of reduced abundance or impaired cytotoxicity, are associated with autoimmune thyroid disorders, suggesting that the dysregulation of the innate immune system is associated with these disorders. 27 In the context of TAI, the dysregulation in the number of NK cells may not only be associated with the production of thyroid autoantibodies but also impairs maternal immune tolerance. In women with a history of miscarriage, abnormalities in NK cell activity and quantity, coexisting with TAI, may exacerbate the immune imbalance associated with pregnancy.
Logistic regression analysis confirmed a significant and consistent inverse association between the pNK% and TAI, which remained robust across all the models. The low ORs suggested that even small reductions in the pNK% cells are associated with substantially higher TAI risk. RCS modeling further revealed a nonlinear relationship, with a steeper risk increase at lower NK cell levels. In Fas-deficient mice (previously known as lpr), NK cells were demonstrated to suppress autoreactive B-cells in vitro, whereas the in vivo NK cell depletion exacerbated autoimmune disease. 28 The immunoregulatory role of NK cells in limiting inflammation could be related to their reported protective effect in some autoimmune diseases.29,30 Women with a history of miscarriage represent a unique population with an increased risk of TAI; however, the underlying immune mechanisms remain poorly understood. This study systematically quantified the relationship between the pNK% and TAI in women with a history of miscarriage. The inverse association remained significant after adjusting for multiple covariates, suggesting a strong association and predictive value of pNK% with TAI.
Stratified analysis revealed significant heterogeneity in the association between the pNK% and TAI, which was jointly driven by age and miscarriage history. These findings suggest that both age and miscarriage history influence the association between NK cells and TAI. Considering the potential collinearity between AMA and cumulative miscarriage risk, we adjusted for these confounding factors in the model. The results indicate that both age and reproductive history significantly alter the association pattern, with nonlinear and linear trends observed in older women and women with RM, respectively. These findings suggest that immune aging and reproductive history interact to regulate NK cell function, rather than functioning as completely independent mechanisms. Mechanistically, age-related immune changes in women aged ≥35 years may be associated with changes in the cytotoxicity and regulatory function of NK cells, potentially correlating with susceptibility to TAI. 31 Although classical immune aging is often associated with late aging, new evidence has suggested that the reproductive immune aging process begins earlier and influences immune tolerance mechanisms. 32 In addition, a history of RM may be associated with adaptive changes in immune regulation.
Mediation analysis revealed a significant inverse association between the pNK% and TAI (total effect = −0.528, SE = 0.058, 95% CI (−0.619, −0.401), p < 0.001). B-cells partially mediated this relationship, with an indirect effect of −0.166 (SE = 0.042, 95% CI (−0.248, −0.081), p < 0.001), contributing approximately 31.4% to the total effect. The direct effect of pNK% on TAI remained significant after adjusting for B-cells (−0.362, SE = 0.069, p < 0.001), consistent with a partial mediation model. Mechanistically, NK cells have been reported to interact with B-cells through cytokine networks (including IFN-γ and TGF-β) and, under specific experimental conditions, through perforin–granzyme–mediated cytotoxicity.33,34 However, the relative contribution of these pathways in human peripheral blood remains incompletely characterized. Although some studies have reported limited interleukin-10 (IL-10) production by certain NK cell subsets under specific stimulation, the predominant sources of IL-10 in the peripheral immune compartment are regulatory B-cells (Bregs) and regulatory T-cells (Tregs), and direct cytolytic elimination of autoreactive B-cells by NK cells in vivo has not been definitively established. Reduced pNK% may therefore be associated with B-cell dysregulation through indirect immunomodulatory crosstalk rather than through direct cytotoxicity; however, this hypothesis requires functional validation. These findings highlight the potential role of the NK–B-cell axis in the immunopathogenesis of TAI in women with a history of miscarriage, providing novel insights into immune dysregulation in reproductive failure. Consistent with the existing literature, our study observed reduced pNK% in women with TAI. NK cells are critically involved in the pathogenesis of autoimmune thyroiditis, and reduced peripheral NK/K-cell levels and cytotoxic activity have been reported in patients with Hashimoto’s disease.9,35 This reduction may not solely reflect impaired hematopoiesis but rather result from the migration and sequestration of NK cells to inflamed thyroid tissues, 36 where they participate in the local immune response via cytotoxic interactions with thyrocytes. 7 Notably, in addition to the pNK%, this study revealed significant associations between TAI and several other autoantibodies, including antisperm, antiendometrial, and anticardiolipin antibodies. Women with autoimmune thyroid disease have a relatively high prevalence of non-organ-specific autoantibodies, which may be associated with an increased risk of adverse pregnancy outcomes.37–40 These findings suggest that the aberrant expression of thyroid autoantibodies is not an isolated event; it reflects a broader state of systemic immune activation, which is often accompanied with the presence of additional autoantibodies.
This study has several strengths. First, this study included a relatively large sample of 927 women with a history of miscarriage, and the exclusion of incomplete or ineligible cases ensured high data integrity and representativeness, which enhanced the reliability of the findings. Second, this study not only assessed the association between NK cells and TAI but also incorporated a mediation analysis to examine the role of B-cells in relation to this association, providing insights into the potential immunological pathways involved and offering a foundation for future mechanistic studies. Third, the robustness of the results was supported by several multivariable logistic regression models adjusted for potential confounders such as age, miscarriage history, and various autoantibodies.
Nevertheless, certain study limitations should be noted. First, due to the cross-sectional design, a causal relationship between NK cells and TAI cannot be established. Although our data reveal a significant association, the underlying mechanism pathways still need to be fully elucidated. Existing literature suggests that NK cells indirectly modulate TAI through cytokine-mediated immunoregulatory networks rather than predominantly through direct cytolysis. The mediation effect of B-cells observed in our analysis should therefore be interpreted as a statistical association consistent with NK–B immunomodulatory crosstalk, rather than direct evidence of NK-mediated elimination of autoreactive B-cells. The contribution of regulatory subsets such as Bregs and Tregs, which are well established as the predominant peripheral IL-10 producers, warrants further investigation. Without longitudinal data or functional assays (such as NK cytotoxicity assays or cytokine analysis), we cannot confirm whether the observed changes in the pNK% are the primary driver of TAI or a secondary consequence of the autoimmune inflammatory environment. Prospective studies incorporating mechanistic endpoints are necessary to dissect these temporal and functional relationships. Second, the lack of strict control over the phase of the menstrual cycle during blood sampling comprises another study limitation. Immune parameters, particularly NK cytotoxicity and lymphocyte subpopulations, fluctuate during the follicular and luteal phases due to the influence of estrogen and progesterone. Although samples were collected outside the menstrual active phase to avoid confounding factors of acute inflammation, variations in cycle phase may have introduced heterogeneity in the immunological data. Although the sampling time was consistent for both case and control groups—reducing the likelihood of differential bias—future investigations should systematically record menstrual cycle phase at sampling, conduct stratified analyses by cycle phase, and incorporate functional NK assays (such as cytotoxicity assays and intracellular cytokine staining for IFN-γ, TNF-α, and granzyme B) along with prospective follow-up to strengthen causal inference and better dissect the immunoregulatory pathways linking NK cells, B-cells, and TAI. Third, thyroid function tests (thyroid stimulating hormone (TSH), free thyroxine (FT4), free triiodothyronine (FT3)) were not consistently available in our retrospective dataset, as these tests were ordered based on individual clinical indications rather than as part of a standardized workup. Consequently, we were unable to stratify our analyses by thyroid functional status (euthyroid, subclinical hypothyroidism, or overt hypothyroidism). This precludes us from determining whether the inverse association between the pNK% and TAI is driven by autoimmunity per se, underlying thyroid dysfunction frequently associated with TAI, or both. Prospective studies incorporating concurrent and standardized thyroid function assessment are warranted to clarify this distinction and elucidate whether the NK–TAI relationship varies across thyroid function strata. Moreover, NK and B-cell levels may vary across pregnancy stages or with repeated miscarriage events; however, this study captured only the immune cell status at a single time point. Finally, as all participants were recruited from a single center, the findings may be subject to regional selection bias; external validation in other populations is, therefore, warranted.
Conclusion
This study revealed a significant inverse association between the pNK% and TAI, which remained robust after adjusting for multiple covariates. These findings suggest that a reduced number of pNKs are associated with the onset and progression of TAI, potentially through indirect immunomodulatory pathways involving B-cells, although the precise molecular mechanisms remain to be elucidated. Moreover, this study underscores the potential interplay between NK and B-cells in TAI immunopathogenesis, offering novel insights into immune dysregulation among women of reproductive age. These results may facilitate early risk identification and targeted immunological interventions in high-risk populations, particularly the female population during the periconceptional and pregnancy periods. Future prospective studies and mechanistic research are needed to validate these findings and further clarify the immunoregulatory role of NK cells in the pathogenesis of autoimmune thyroid disease.
Footnotes
Acknowledgments
We would like to thank all members of the Reproductive Medicine Center at the Affiliated Hospital of Guizhou Medical University for their technical assistance and support. Special thanks to the patients who participated in this study and made this research possible.
Authors’ contributions
X. Guo: conceptualization; methodology; investigation; data curation; and writing – original draft. T. Yang: formal analysis; writing – original draft; and writing – review & editing. X. Zhong: methodology and resources. J. Wang: investigation; data curation; and resources. S. Zhao: supervision; project administration; and writing – review & editing. G. Huang: conceptualization; supervision; project administration; and writing – review & editing. S. Zhao and G. Huang are corresponding authors. All authors read and approved the final manuscript.
Consent for publication
Not applicable.
Data availability statement
The deidentified datasets generated and analyzed during the current study are available from the corresponding author upon reasonable request and subject to institutional ethical approval. Individual participant data will not be made publicly available to protect patient privacy in accordance with institutional data protection policies.
Declaration of conflicting interests
The authors declare that they have no competing interests.
Ethics approval and consent to participate
The study was approved by the Ethics Committee of the Affiliated Hospital of Guizhou Medical University (2025(284)). Written informed consent was obtained from all participants prior to enrollment in the study. All procedures involving human participants were conducted in accordance with the ethical standards of the institutional and/or national research committee and the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards.