
Abstract
Flavor recognition times were measured in 23 young, healthy participants of both sexes using an electromyography-based system. The participants were instructed to chew a gummy candy, which was randomly selected among six commercially available types, and to press a button immediately on flavor recognition. A total of 107 normally distributed, flavor recognition times were analyzed, with an average time of 7.5 seconds (± 2.34 seconds, standard deviation). No significant differences were found among the six types of candies in terms of recognition time. Analysis of the association between flavor recognition and chewing phase showed that 70 (65%) of the analyzed 107 recognition signals occurred between 0.2 seconds before and 0.4 seconds after the end of jaw closing. Recognition signals occurring during the jaw-opening phase prolonged its duration by an average of 21%, whereas those occurring during the jaw-closing phase did not influence it.
Introduction
In humans, simple reaction times are usually measured through a conventional method, according to which participants are asked to push a button as soon as they recognize a delivered sensory stimulus. Although research on simple reaction times to visual and acoustic stimuli has a long history and has generated a massive amount of data, the reaction times to chemical stimuli remain unclear, despite the fact that everyday life foods contain various flavors that are perceived through the interaction between taste, smell (aroma), and trigeminal sensations (Prescott, 1999; Small & Prescott, 2005). Fundamental results concerning simple reaction times to chemical stimuli are as follows: the response times of young healthy participants to a 1.0 M sucrose solution varied between 0.5 and 1.0 seconds (Bujas, Szabo, Ajdukovic, & Mayer, 1989; Yamamoto & Kawamura, 1981), whereas their response times to eucalyptol and benzaldehyde were about 1.4 seconds (Keita, Frasnelli, La Buissonniere-Ariza, & Lepore, 2013). In contrast to the conventional method, a new approach comprising a model of probability summation has provided valuable findings (Marks, Veldhuizen, Shepard, & Shavit, 2012; Shepard, Veldhuizen, & Marks, 2015; Veldhuizen, Siddique, Rosenthal, & Marks, 2017).
Flavor perception depends on human-related aspects, such as the orthonasal and retronasal perception routes, and on food-related aspects, such as the volatile and nonvolatile components present (Salles et al., 2011). During eating and drinking, the retronasal route plays an important role in flavor perception (Frasnelli, van Ruth, Kriukova, & Hummel, 2005; Halpern, 2004; Sun & Halpern, 2005). Volatile components emitted from the food to the mouth are transported to the olfactory epithelium, generating neural signals that are conveyed to the central nervous system. The transport of volatile substances to the olfactory epithelium is influenced by two major functions related to flavor perception: breathing and chewing. Periodicity is a prominent feature of both these functions, and previous studies have looked into the effect of breathing on flavor perception at the phase level of periodicity (i.e., inspiration and expiration; Masaoka, Satoh, Akai, & Homma, 2010; Sun & Halpern, 2005). Previous studies have also analyzed the effects of mastication on brain activity (Sakamoto, Nakata, Honda, & Kakigi, 2009; Sakamoto, Nakata, & Kakigi, 2009; Takada & Miyamoto, 2004), but few have looked into the functional relationships at the level of phase periodicity (i.e., jaw opening and jaw closing). Thus, this study aimed to examine the influence of the two chewing phases on flavor recognition time.
Materials and Methods
Participants
The participants were 23 healthy, young adults (14 males and 9 females, average age 22.1 ± 4.71 years), who participated in this study as an experimental group after providing informed consent. Eleven of the 23 participants were also included as a control group (three males and eight females). The study protocols were approved by the Ethics Committee of the University of Niigata Rehabilitation Graduate School.
Test Foods
Six commercially available gummy candies (Meiji Co., Ltd., Tokyo, Japan) with different fruit flavors (apple, grape, orange, (European) pear, pineapple, and strawberry) were used for this study. The candies had similar shapes and sizes, with an approximate height, width, and thickness of 9.5, 23.6, and 15.0 mm, respectively. Each candy was wrapped with wafer paper (Kokko Oblaat Co., Ltd., Shizuoka, Japan) to prevent direct contact with the oral tissues before chewing. The textural properties of the test foods were demonstrated in a previous study from our group (S. Miyaoka, Yamazaki, Ito, & Miyaoka, 2014).
Procedures
The procedures used in this study for the experimental group were essentially identical to those described in a previous study by our group (S. Miyaoka & Miyaoka, 2013). (a) Each participant was first instructed to rinse his or her mouth with tap water. (b) A surface electromyogram (EMG) was recorded from the masseter (Mass) muscle, and the Mass EMG signals were amplified, filtered, fully rectified, and stored on a digital recorder. (c) Each participant held a pressing button, which was connected to the recorder through a battery. (d) One candy was randomly selected from among the six available candies and given to each participant. The participants were instructed to hold the candy between the molars until they were asked to start chewing and to press the button immediately on flavor recognition. (e) Finally, after the task, the participants were instructed to report the recognized flavor and to describe any special impressions that they might have experienced. Before the task, the subjects received no information regarding the taste, odor, and flavor of the candies. The 11 participants in the control group were asked to press the button at any time they wanted but not to respond to gummy candies delivered randomly. In both the experimental and the control groups, each experimental session consisted of three to six trials with a minimum intertrial interval of 4 minutes used for the flavor report and description (for about 2.5 minutes) and a brief break (for about 1.5 minutes).
Data and Statistical Analyses
A total of 181 recognition times were collected from the participants, among which 107 showed a normal distribution (S. Miyaoka, Iwamori, & Miyaoka, 2018), and were extracted and analyzed. The number of recognition times varied largely among participants with an average of 4.7 ± 3.6 (range, 0–12) recognition times and also varied among the used gummy candies (see Table 2 in S. Miyaoka et al., 2018 for details). The flavor recognition time was measured on each participant’s usual working side in this study because our previous study showed that the chewing side did not significantly affect flavor detection or recognition times (S. Miyaoka & Miyaoka, 2013).
The chewing phases were determined electromyographically on the basis of the Mass activity. The end of the Mass activity was regarded as the end of the jaw-closing phase and used as a reference for the analysis. Onset and offset of the jaw-closing phase were determined by the Mass EMG in each participant (Figure 1). The following four steps were used for the determination: (a) the average and standard deviation of the Mass EMG were calculated from numerical data (positive values only) during a resting period for about 30 seconds at the beginning of each session, (b) threshold for each participant was set by an equation of “average + 3.0 × standard deviation,” (c) onset of the jaw-closing phase was determined if the Mass EMG firstly exceeded the threshold and exceeded EMG signals were converted to pulses, and (d) offset was determined if the Mass EMG firstly fell the threshold. There was an additional condition for the determination, that is, the onset and offset were adopted only when successive EMGs lasted and ceased, respectively. For example, three pulses shown by closed circles in Figure 1 were not adopted according to the additional condition. The threshold was reset if recording conditions were largely altered by unpredictable reasons resulting from mainly electrodes.

Determination of onset and offset of the jaw-closing phase. Onset and offset of the jaw-closing phase (indicated by open arrows) were determined by an electromyogram of the masseter (Mass). Hatched squares above the “Raw Mass EMG” indicate the jaw-closing phases. These phases were determined by the “Converted pulses,” which were given if the “Raw Mass EMG” exceeded a threshold which was determined by the average and standard deviation of a resting period at the beginning of each session, and the equation for the threshold was “average + 3 × standard deviation.” Converted pulses indicated by closed circles exceeded the threshold but were not adopted. See the text for details. Raw Mass EMG: raw masseter electromyogram.
A χ2 test, analysis of contingency tables, was used to examine the association between flavor recognition and the two chewing phases—jaw opening and jaw closing. The χ2 test was also used to examine sampling biases in the analyzed 107 recognition times and followed by Ryan’s multiple-comparison test for a specific difference. Paired t tests were used to examine the following parameters: number of recognition signals occurring during each chewing phase, duration of each chewing phase that preceded flavor recognition signals or during which flavor recognition signals occurred, and time between the flavor recognition signals and the beginning of the jaw-closing phase. p < .05 was considered to be statistically significant.
Results
A total of 107 recognition times for six fruity-flavored gummy candies were analyzed (S. Miyaoka et al., 2018). In the experimental group, 93 and 14 of the 107 recognition times occurred during the jaw-opening phase and during the jaw-closing phase, respectively. The χ2 test showed that the numbers of recognition times differed significantly between the two chewing phases with respect to the average durations of these phases (0.54 seconds for jaw-opening phase and 0.45 seconds for jaw-closing phase; p < .01). The χ2 test also found a significant difference in the 107 recognition times among the six gummy candies (p < .01), and Ryan’s multiple-comparison test detected a specific difference between grape- and pear-flavored gummy candies only (p < .01). However, their average recognition times and numbers that occurred during the two chewing phases were not significantly different between these two gummy candies. In the control group, 36 and 50 presses occurred during the jaw-opening and the jaw-closing phase, respectively, but the numbers of presses did not differ between the two chewing phases with respect to the average durations of these phases (0.46 seconds for jaw-opening phase and 0.54 seconds for jaw-closing phase).
Flavor recognition times ranged between 2.6 and 12.3 seconds (7.5 ± 2.35 seconds). Seventy (65%) of the analyzed 107 recognition signals occurred between 0.2 seconds before (i.e., −0.2 seconds) and 0.4 seconds after (i.e., +0.4 seconds) the end of jaw closing. A Pearson’s χ2 test indicated a significant difference between the observed and expected frequencies of the seven intervals using the class values from −0.3 to +0.9 seconds (p < .01). Furthermore, the frequency at the class value of 0.1 seconds (n = 32) was significantly higher than expected (p < .05), whereas the frequencies at the class values from −0.3 to −0.1 (n = 1) and 0.9 seconds (n = 1) were lower than expected (p < .05). The average duration of the jaw-opening phase where recognition signals occurred corresponded to 121.6% of the duration of the preceding phase (Figure 3), indicating that the occurrence of recognition signals prolonged the duration of the jaw-opening phase. However, signals occurring during the jaw-closing phases did not affect the duration. A paired t test detected a significant difference in the average duration of the jaw-opening phases (p < .01), whereas no significant differences were found concerning the duration of the jaw-closing phases.
Discussion
In this study, masseter activity was used as a reference for the jaw-closing phase (Figure 1) because higher temporal resolution was expected in the EMG activity rather than jaw movement curves. However, the masseter is merely one of the components for the jaw-closing phase. The temporalis and medial pterygoid are included in the components as well, and the electromyographic onsets of these jaw-closing muscles slightly differ from each other during natural chewing in humans (Moller, 1966; Vitti & Basmajian, 1977). According to a previous study (Moller, 1966), the medial pterygoid fired first, the masseter followed, and the (posterior) temporalis fired finally during chewing of an apple. Less than a 0.1-second difference was seen between the first and last firing of these muscles in a chewing cycle. The average time difference suggests that selection of either one of the jaw closing muscles does not seriously affect the interpretation of timing of recognition in relation to jaw closing that is analyzed by 0.2-second class intervals (Figure 2). Moreover, we analyzed the Mass EMG and jaw movement curves with an accelerometer (vertical direction only) concomitantly in 60 chewing cycles that were previously recorded during chewing of an apple-flavored gummy candy in healthy young adults. The analysis showed that the average time of firing of the Mass EMG slightly preceded (19.4 ± 67.6 milliseconds) that of jaw closing. The result may support the validity for the use of the Mass EMG as the reference for chewing phases in this study.
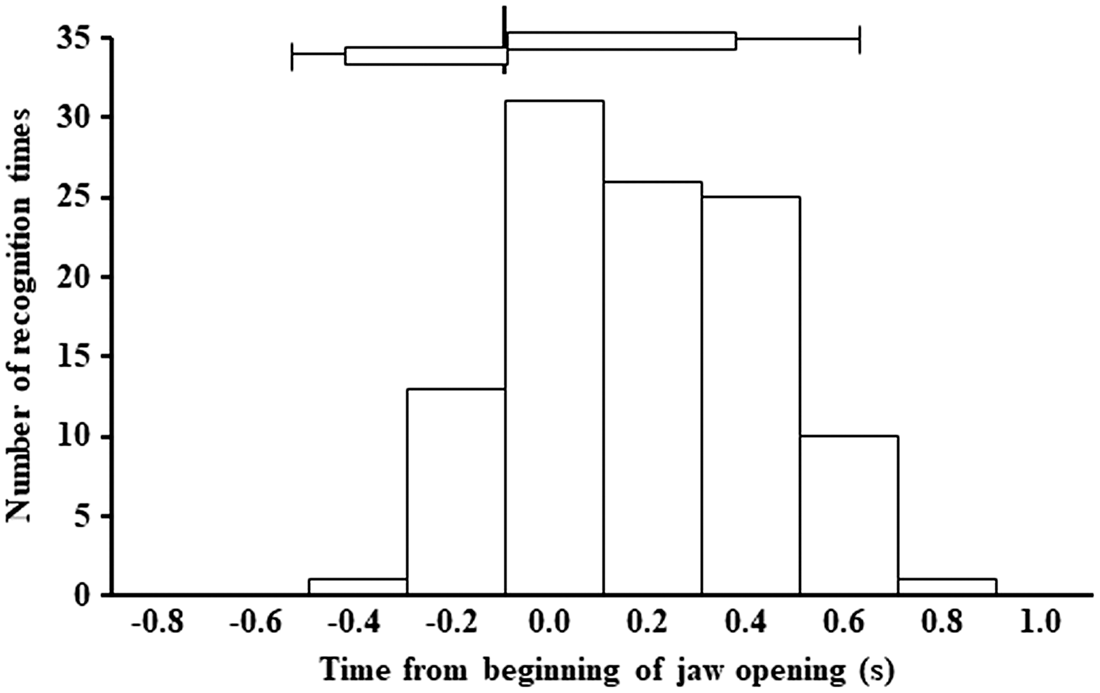
Histogram showing the frequency distribution of the flavor recognition signals, according to the chewing cycle. Time “0” corresponds to the end of the jaw-closing phase, defined by the masseter muscle activity. Frequency bars on positive scales (n = 94) indicate recognition signals occurring during the jaw-opening phase, whereas those on negative scales (n = 13) indicate recognition signals occurring during the jaw-closing phase. Horizontal bars with error bars indicate the average durations of the jaw-opening (right side) and jaw-closing (left side) phases and their standard error of the mean. See the text for details.
In general, the reaction time comprises two features: “perceptual latency,” which corresponds to the time from stimulus onset to motor system activation, and “motor delay,” which corresponds to the time from motor system activation to the final response action (i.e., pressing the button, in this study; Jaskowski, Pruszewicz, & Swidzinski, 1990; Menon, Luknowsky, & Gati, 1998). In this study, the mode (n = 31) was exhibited in the interval between 0.0 and 0.2 seconds of the flavor recognition time frequency distribution (Figure 2). However, the motor delay must be carefully considered when interpreting the mode because the primary interest of reaction time studies is the perceptual latency (and not the motor delay). The end of the masseter muscle activity was used as the reference point for frequency distribution. Thus, motor delay must be subtracted from the mode to approach the real perceptual latency. The documented average motor delay ranges between 120 and 140 milliseconds (Jaskowski et al., 1990; Wise, Weinrich, & Mauritz, 1986). It is believed not to differ largely among sensory modalities because the motor pathway between the cortical motor areas and the effectors (i.e., muscles of the hand and fingers) should present reaction times common to different sensory modalities. Should we presume a motor delay of about 130 milliseconds, then the overall distribution and part of the mode (Figure 2) would be moved to the left. Therefore, flavor recognition occurs most frequently during the periods before and after the end of jaw closing.
There are at least two possible explanations for the functional coupling between flavor recognition and the chewing phases shown in this study (Figure 2). The first is related to mouth movement. A previous study using flavored solutions showed that mouth movements affect the intensity of retronasal olfactory perception (Burdach & Doty, 1987). The authors compared the relative olfactory intensity of each solution with and without mouth movements and found that mouth movements enhanced it. Several studies have looked into the role of oral functions on the perception intensity of food smells and flavors (Foster et al., 2011; Haahr et al., 2004; Salles et al., 2011). We recently used an electromyography-based measuring system and reported that the occurrence of flavor recognition signals for fruit-flavored gummy candies tended to be higher in the jaw-closing phase than in the jaw-opening phase (Y. Miyaoka, Miyaoka, & Ashida, 2016). The present results (Figure 2) suggest that the jaw-closing movement contributes positively for flavor recognition. However, it remains unclear whether that contribution arises from the mechanical influence of the jaw movement for the orthonasal olfaction of flavors.
The second possibility is related to respiratory movements. Previous studies in adult humans and awake rabbits investigated the functional coupling between mastication and respiration during chewing (Fontana et al., 1992; McFarland & Lund, 1993, 1995). These studies reported inconsistent results by determining strong (Fontana et al., 1992), weak (McFarland & Lund, 1995), or absent (McFarland & Lund, 1993) coupling between the two functions. Such functional coupling may support the findings of this study because at least one of the previous studies showed a positive functional coupling between inspiration and jaw opening in the adult human (Fontana et al., 1992). The present results (Figure 2) can be explained in the light of the functional coupling between inspiration and jaw-opening, but in the near future, a precise analysis of the timing among flavor recognition, chewing, and breathing should be performed.
In addition, we found that recognition signals influenced the duration of the jaw-opening phase (Figure 3). Although previous studies have investigated the relationship between the oral processing of foods and their flavor release (Foster et al., 2011; Salles et al., 2011), few have examined the relationship between chewing phases and flavor recognition, making it harder to hypothesize which neural mechanism underlies this association (Figure 3). There seems to be a possibility that the recognition task itself was involved in the prolongation of the jaw-opening phase, but it is likely to be excluded because pressing a button arbitrarily in the control group did not affect the duration of jaw-opening phase. Neurophysiological studies in the field of mastication have shown that some trigeminal reflexes are differentially evoked during the jaw-opening and jaw-closing phases, suggesting functional differences in the afferent signals from the peripheral receptors and in the excitability states of the central nervous system between the two chewing phases (e.g., Haraguchi et al., 1994; Lund et al., 1983; van der Bilt, Ottenhoff, van der Glas, Bosman, & Abbink, 1997; van der Glas, van der Bilt, Abbink, Mason, & Cadden, 2007; Yamada, Yamamura, & Inoue, 2005). Moreover, both serotonin and norepinephrine may be involved in the central nervous mechanism for the prolongation of the jaw-opening phase (Figure 3) because an iontophoretic application of these substances to the digastric motoneurons facilitated their activity evoked by cortical stimulation (Katakura & Chandler, 1990). However, further discussion is clearly beyond the scope of this study.

Average duration of the JO and JC phases in relation to the occurrence of recognition signals. Average (plus standard error of the mean) durations of the JO and JC phases with the occurrence of recognition signals (closed bars) and the phases just before the occurrence (open bars), respectively. **p < .01. JO = jaw opening; JC = jaw closing.
In conclusion, the flavor recognition time for the six fruit-flavored gummy candies was analyzed in 23 young, healthy participants using an electromyography-based measuring system. Seventy (65%) of the analyzed 107 recognition signals occurred from −0.2 to +0.4 seconds from the end of jaw closing. Recognition signals occurring during the jaw-opening phase prolonged its average duration by 21%, whereas its occurrence did not affect the jaw-closing phase.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially supported by Scientific Research from the Ministry of Education, Science and Culture of Japan under Grants-in-Aid (No. 26350106 to S. M. and No. 16K00830 to Y. M.).