
Abstract
It has been argued that movement can be implied by static cues of images depicting an instance of a dynamic event. Instances of implied motion have been investigated as a special type of stimulus with common processing mechanisms to those of real motion. Timing studies have reported a lengthening of the perceived time for moving as opposed to static stimuli and for stimuli of higher as compared to lower amounts of implied motion. However, the actual comparison of real versus implied motion on timing has never been investigated. In the present study, we compared directly the effect of two hypothetically analogous ballet steps with different amounts of movement and static instances of the dynamic peak of these events in a reproduction task. The analysis revealed an overestimation and lower response variability for real as compared to implied motion stimuli. These findings replicate and extend the apparent duration lengthening for moving as compared to static stimulation, even for static images containing implied motion, questioning whether or not the previously reported correspondence between real and implied motion transfers in the timing domain. This lack of correspondence was further supported by the finding that the amount of movement presented affected only displays of real motion.
Introduction
Imagine you are a hero in a comic book and you are running away from the bad guys, what kind of image would convey this dynamic event? This is a tangible example of an image implying motion. The use of a single image to suggest movement has been of great interest for both artists (e.g., Dobrez, 2013; Nather & Bueno, 2012c; Reason, 2004) and scientists (e.g., Cutting, 2002; Nather & Bueno, 2008). The seminal memory tests of Freyd and coworkers (Freyd, 1983; Freyd & Finke, 1984) have illustrated the dynamic nature of mentally extrapolating (i.e., “representational momentum”) an action that is implied in frozen-action photographs (see Freyd, 1987; Thornton & Hubbard, 2002, for reviews). More recently, Acik, Bartel, and Konig (2014) using eye-tracking demonstrated similar fixation selectivity when comparing real versus implied motion stimuli, concluding that static cues can be sufficient in revealing movement in a scene. Similarly, Elseways et al.’s (2014) visual working memory study using dance stimuli also supported that recognition of movement based only on posture is possible, even though somewhat poorer than that of real moving stimuli. The link between real and implied motion has also been supported through data showing common neuronal substrates (i.e., medial temporal/medial superior temporal cortex) in both humans (e.g., Cattaneo, Schiavi, Silvanto, & Nadal, 2017; Concerto et al., 2016; Kim & Blake, 2007; Kourtzi & Kanwisher, 2000; Lorteije et al., 2006; Lu, Li, & Meng, 2016; Mineo et al., 2018; Osaka, Matsuyoshi, Ikeda, & Osaka, 2010; Proverbio, Riva, & Zani, 2009; Senior et al., 2000; Urgesi, Moro, Candidi, & Aglioti, 2006; see also David & Senior, 2000; Kourtzi, 2004, for reviews) and monkeys (e.g., Jellema & Perrett, 2003; Krekelberg, Dannenberg, Hoffmann, Bremmer, & Ross, 2003). Moreover, common direction-selective circuits for both real and implied motion have been suggested by motion and positional after-effect experimentation in both adults and infants (e.g., Lorteije et al., 2007; Pavan, Cuturi, Maniglia, Casco, & Campana, 2011; Shirai & Imura, 2014, 2016; Winawer, Huk, & Boroditsky, 2008).
The role of implied motion in perception has recently drawn the attention of researchers in the timing domain given the tight link of motion and timing. Specifically, Nather and coworkers (see Bueno & Nather, 2012, for a short review) have investigated extensively the effect of implied motion on duration estimates utilizing dance stimuli (i.e., images of Degas’ ballerinas-statues with various body postures implying different amounts of movement/induced intensity). For instance, Nather, Bueno, Bigand, and Droit-Volet (2011), via a bisection task (where short and long standard durations are learned by the participants and, subsequently, a series of test intervals are discriminated as more similar to the short or long standards), reported a greater overestimation for statues depicting movement of higher as compared to lower intensity for short durations (i.e., 0.4 to 1 s), while no such effect was noted for longer intervals (i.e., 2 to 8 s). These differences were attributed to the transient effect of arousal induced by high intensity images that deteriorated for longer intervals. However, when a reproduction task was utilized, an overestimation of higher as compared to lower intensity images was found for an interval of 36 s (Nather & Bueno, 2011; see also Nather & Bueno, 2012a; but see Nather, Bueno, De Abreu, & Gomes, 2010, for a null effect). This finding cannot be accounted for by an arousal modulation and was, thus, interpreted based on the “change model” (e.g., Poynter, 1989), where body positions implying motion of higher as compared to lower intensity were considered as stimuli that involved more changes/events resulting in an interval overestimation. Further efforts to clarify such inconsistencies showed that the: intervals presented (e.g., Nather & Bueno, 2012b, examined intervals of 9, 18, 27, and 45 s and found an overestimation for stimuli of higher intensity in implied motion only for the 27 s), task implemented (e.g., eye-tracking along with force-platform in a reproduction task; Nather, Bueno, & Bigand, 2009, 2013; Nather et al., 2010), and type of stimuli utilized (e.g., cubist-abstract paintings, optical art, animals vs. human figures; Nather, Anelli, Ennes, & Bueno, 2015; Nather, Fernandes, & Bueno, 2012, 2014; Nather, Mecca, & Bueno, 2013), could modulate time estimation of implied motion. Overall, this line of research does not allow for any integrative conclusions on the role of implied motion on timing, as results are often contradictory.
Similarly, timing studies utilizing naturalistic images—other than dance—with or without implied movement have also resulted in inconsistencies. For instance, Moscatelli, Polito, and Lacquaniti (2011) utilized images of athletes with or without implied movement in a time discrimination task (for intervals in the range of 0.5–1.1 s) and even though participants were more accurate for images depicting implied motion (i.e., lower just noticeable difference scores), no significant difference in implied versus static images was noted (i.e., similar point of subjective equivalence). Yamamoto and Miura (2012), however, when comparing images of human and animal characters in an either running or standing position (for intervals ranging 0.4–1 s) reported overestimations for the former as compared to the latter stimuli. The exact opposite result was reported in Chen, Pizzolato, and Cesari’s (2014) reproduction experiment, where images of a pole-vaulter (highly dynamic posture), fencer (less dynamic posture), and scrambled images (static) were compared in both sub- (0.3–1 s) and supra-second (1.1–1.8 s) intervals. That is, while no effect was found for longer durations, in the subsecond range, images depicting highly dynamic actions were underestimated in comparison to less dynamic or static ones. These findings were interpreted as the result of differential attentional allocation, with highly dynamic pictures being more attentional capturing and, thus, leading to interval underestimations (e.g., Chen et al., 2014).
Although time estimation with implied motion stimulation has led to conflicting results, the same is not true for experiments comparing real motion versus static (without implied motion) presentations, where a time dilation for the former as compared to the latter has been repeatedly illustrated (e.g., Brown, 1995; Kanai, Paffen, Hogendoorn, & Verstraten, 2006; Kaneko & Murakami, 2009; Lhamon & Goldstone, 1975; Mitrani & Stoyanova, 1982; Roelofs & Zeeman, 1951). This dilation effect has been attributed to the number of changes (i.e., speed, motion coherence, and spatial and temporal frequency; i.e., “change model”; e.g., Poynter, 1989) present in a moving as compared to a static stimulus. Under this theoretical framework and the previously reported correspondence of real and implied motion (e.g., David & Senior, 2000; Freyd, 1987; Kourtzi, 2004; Thornton & Hubbard, 2002), one wonders how a direct comparison of real and implied motion would affect time estimations given that both types of stimulation contain actual or implied changes, respectively (cf. Moscatelli et al., 2011).
In the present study, therefore, we sought to address this gap in the literature by comparing—through a reproduction task—displays of a dancer performing ballet steps with varying degrees of movement and static snapshots of these steps that imply the analogous ballet movements. Inspired by the Nather et al. studies (e.g., Bueno & Nather, 2012), we utilized ballet movements given their highly dynamic nature. According to the real-implied motion correspondence (e.g., David & Senior, 2000; Kourtzi, 2004) and the “change model” (e.g., Poynter, 1989), we hypothesized that both real and implied motion presentations will have a similar effect on duration estimates, given that both types of stimuli (i.e., real and implied) were derived from the same ballet step and, thus, should involve the same number of real or implied changes. Furthermore, we expected stimuli with higher amounts of motion, hence containing greater number of changes (both for real and implied conditions) to be overestimated 1 as compared to those with lower amounts of movement (see Bueno & Nather, 2012, for a review; Nather & Bueno, 2011, 2012a, 2012b; Nather et al., 2011; Yamamoto & Miura, 2012). This hypothesis is also in accordance with the theory of magnitude (e.g., Walsh, 2003), which proposes a tight link between space and time due to the experienced statistical contingencies of the physical world (e.g., Bueti & Walsh, 2009). Under this notion, time, space, and quantity share a positive relationship, hence overestimations are expected for stimuli with higher as compared to lower amounts of movement (i.e., more movement or greater displacement to lead to lengthened time estimates; e.g., Rammsayer & Verner, 2014, 2015; Sgouramani & Vatakis, 2014, 2015b; Xuan, Zhang, He, & Chen, 2007; but see Yates, Loetscher, & Nicholls, 2012, for opposite task-driven results).
Methods
Participants
Twenty-one participants (M = 27.7 years, age range: 23–35; 13 females) without any dance experience took part in the experiment. All participants reported normal or corrected-to-normal visual acuity. The experiment was performed in accordance with the ethical standards laid down in the 2013 Declaration of Helsinki and informed consent was obtained from all participants. The study was approved by the University’s Ethics Committee.
Stimuli and Apparatus
To directly compare real and implied motion, we created stimuli that were derived from the same source stimulus for both conditions (i.e., no background or any other visual feature differences; cf. Yamamoto & Miura, 2012). In addition, we carefully controlled for the body posture of the dancer and, in particular, we focused on the position of arms and legs based on Nather et al.’s (2013) eye-tracking findings on the most attentional capturing elements of the ballerina’s statues. Thus, in the real motion conditions, the video displays of the two ballet steps presented (i.e., battement tendu derriere and first arabesque) differed on the height of the extended leg from the floor (i.e., the leg extended backwards either remained on the floor or was lifted up to 90°) and the height of the arms (i.e., the arms were placed either below or above shoulder level). For the implied motion conditions, static images were derived from the last frame of the video displays of the respective real motion conditions. These instances depicted the final pose; thus, the image of the dancer with the leg extended on the floor or lifted and with the arms below or above shoulder level, respectively (i.e., lower vs. higher amounts of movement, respectively; Vatakis & Sgouramani, 2015; see Figure 1).

Static frames extracted from the original videos displaying versions of (a) lower and (b) higher (as rated) amounts of movement for both the real and implied (highlighted on the right hand end) conditions.Note: Please refer to the online version of the article to view the figures in colour.
The stimuli were recorded in a dance studio using a Sony PMW-EX1 high definition camera. The original video clips of real motion lasted 840 ms. Six different intervals were used (840, 1040, 1280, 1520, 1760, and 2000 ms) for both the real and implied motion conditions, resulting in a total of 12 stimulus presentations. In order to avoid the interference of factors such as speed, stimulus change, and spatial-related cues in the timing task, the above-mentioned intervals were created by adding static frames at the beginning and end of each video clip (see Sgouramani & Vatakis, 2014). These static frames were extracted from the first and last frames of the original video clip using Adobe Premiere Pro CS5.5. No sound stimulation was presented.
To ensure there was indeed a difference in the perceived movement for the images presented, an online survey was conducted (using Survey Gizmo; http://www.surveygizmo.com). Forty-three nondancers, who did not participate in the main experiment, completed the survey. The participants viewed and rated the stimuli in terms of the amount of implied motion they contained using a Likert scale from 1 to 7 (with 1 being no motion/static and 7 being extremely high amounts of motion). The analysis showed that the images of the battement tendu step were rated as implying significantly less motion (M = 2.28) than those depicting an arabesque step (M = 4.42), t(42) = –8.47, p < .001, as expected.
Stimuli were presented on an Acer, ASPIRE 7750G laptop. The experiment was conducted using Presentation programming software (Version 15.0; Neurobehavioral Systems Inc.). Ten repetitions of each condition were divided into two blocks with a short break in between. The order of stimulus presentation was randomized using shuffling without replacement.
Experimental Procedure
Participants were seated approximately 60 cm from the computer screen in a dimly light room and performed a reproduction task. They were asked to watch a video clip of a particular duration, which was then followed by the presentation of a blue square, hence the transition from the encoding to the reproducing phase was easily noticeable. Their task was to reproduce the duration of the video clip presented by allowing the blue square to remain on the screen for an equal amount of time. When the participants thought the elapsed time was equal to the video clip duration, they pressed the “Enter” key and the next trial was initiated. The participants were instructed not to use any counting strategies. Experimentation did not exceed the 20 min with a break included.
Results
Two psychometric measures were derived from the raw participant’ data, the accuracy (i.e., estimated time divided by the original duration in each condition) and the coefficient of variation (CV; i.e., the standard deviation divided by the mean duration judgment). Accuracy indicates whether participants had underestimated (<1) or overestimated (>1) the physical duration of a given interval. CV is a measure of the participants’ response variability, with higher CV’s indicating greater response variability. Analysis was performed using a three-way repeated measures analysis of variance (ANOVA) with the factors of Motion (real vs. implied motion), Amount of Motion (more vs. less motion), and Duration (840, 1040, 1280, 1520, 1760, and 2000 ms). For all the analyses reported here, Bonferroni corrected t tests (with point of statistical significance set to p < .05) were used in all post-hoc comparisons.
The accuracy analysis revealed a significant main effect of Motion, F(1,20) = 10.14, p = .005, η2 = 0.33, with stimuli containing real motion being judged as lasting longer (M = 1.15) than those containing implied motion (M = 1.04). A significant main effect of Amount of Motion was also obtained, F(1,20) = 18.35, p <.001, η2 = 0.47, with displays containing higher amounts of movement (M = 1.12) being judged as lasting longer than those that contained lower amounts of movement (M = 1.07). In addition, a significant main effect of Duration was obtained, F(5,100) = 79.58, p < .001, η2 = 0.79, with shorter durations being overestimated as compared to longer intervals (M = 1.49, 1.25, 1.09, 0.99, 0.91, and 0.84 for 840, 1040, 1280, 1520, 1760, and 2000 ms, respectively), a finding consistent with Vierordt’s law (1868; i.e., as interval duration increases the associated temporal estimation decreases). Moreover, the interaction of Duration by Motion was significant, F(5,100) = 28.49, p <.001, η2 = 0.58, where for the first four intervals tested (i.e., 840, 1040, 1280, and 1520), stimuli containing real motion (M = 1.63, 1.31, 1.15, and 1.03, respectively) were estimated as lasting longer than those containing implied motion (M = 1.34, 1.19, 1.03, and 0.95, respectively), while for the two longest durations tested (i.e., 1760 and 2000 ms) this difference failed to reach significance (M = 0.93, 0.83, and 0.90, 0.85 for real and implied motion, respectively; see Figure 2(a)). Finally, the interaction of Motion and Amount of Motion was also significant, F(1,20) = 18.39, p < .001, η2 = 0.47], with stimuli containing higher amounts of movement being judged as lasting longer than those containing lower amounts of movement (M = 1.19 and 1.10, respectively), but only for the real motion conditions. No such differences were noted for the implied motion stimuli presented (M = 1.05 and 1.03 for higher and lower amounts of movement, respectively; see Figure 2(b)). All other interactions did not reach significance—Duration by Amount of Motion: F(5,100) = 1.12, p > .05, η2 = 0.05, and Duration by Amount of Motion by Motion: F(5,100) = 0.97, p >.05, η2 = 0.04.
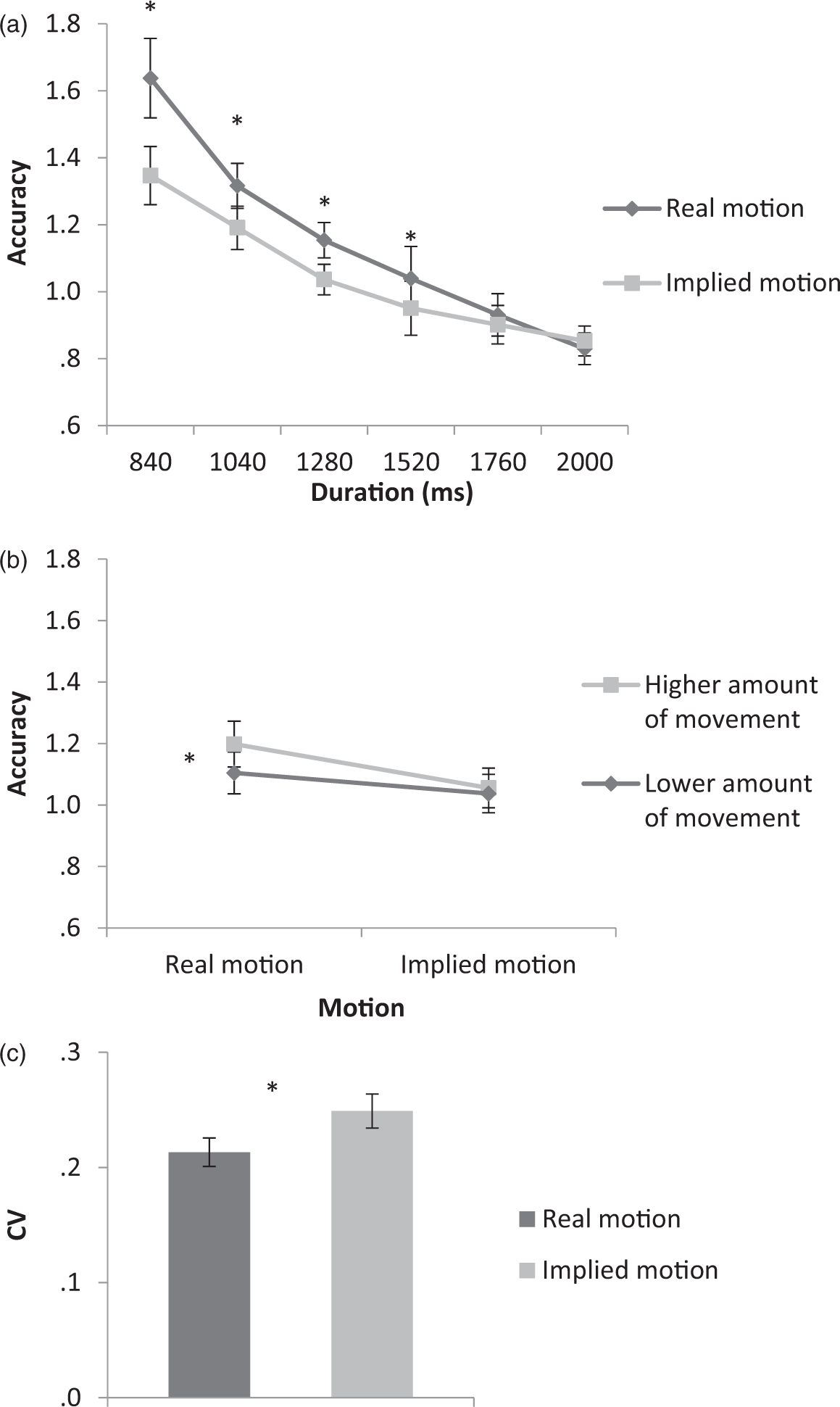
Mean accuracy of the interaction of Motion (real vs. implied) by (a) Duration (840, 1040, 1280, 1520, 1760, and 2000 ms) and (b) Amount of Motion (higher vs. lower), as well as (c) the mean CV of the Motion (real vs. implied) conditions. The error bars represent the standard errors of the means. Significant differences (p < .05) are highlighted by an asterisk. CV = coefficient of variation.
The CV analysis showed a significant main effect of Motion, F(1,20) = 8.66, p = .008, η2 = 0.30, with participants having higher response consistency when viewing the real (M = 0.21) as compared to the implied (M = 0.24) motion stimuli (see Figure 2(c)). All other main effects and interactions failed to reach significance—Duration: F(5,100) = 1.83, p >.05, η2 = 0.08, Amount of Motion: F(1,20) = 1.71, p >.05, η2 = 0.07, Duration by Motion: F(5,100) = 1.66, p >.05, η2 = 0.07, Amount of Motion by Motion: F(1,20) = 0.12, p >.05, η2 = 0.72, Duration by Amount of Motion: F(5,100) = 1.23, p >.05, η2 = 0.05, and Duration by Amount of Motion by Motion: F(5,100) = 1.02, p >.05, η2 = 0.04.
Discussion
In the present study, we investigated—for the first time—time estimates for ballet steps of real as compared to implied motion and with different amounts of movement using a reproduction task. Real motion stimuli were overestimated and had a higher response consistency (i.e., lower CV) when compared to stimuli of implied motion. In addition, when ballet steps containing real motion were presented, participants overestimated the stimuli that contained higher as compared to lower amounts of movement, while no such differences were noted for ballet steps containing implied motion. These results raise the potential non-correspondence between real and implied motion stimulation in the timing domain as demonstrated in other perceptual domains (see David & Senior, 2000; Freyd, 1987; Kourtzi, 2004; Thornton & Hubbard, 2002).
The overestimation noted in our data for real as compared to implied moving ballet steps contradicts our original hypothesis of similar modulating patterns for both real and implied motion on duration estimates, but is in line with previous findings of an apparent lengthening of the duration of real motion stimuli as compared to static—without implied motion—ones (e.g., Brown, 1995; Kanai, Paffen, Hogendoorn, & Verstraten, 2006; Kaneko & Murakami, 2009; Lhamon & Goldstone, 1975; Mitrani & Stoyanova, 1982; Roelofs & Zeeman, 1951). This latter comparison has been reported many times for abstract stimuli and the present work replicates and extends these findings for more complex stimuli such as those of ballet steps utilized in this study. Notably, even though we utilized images depicting the analogous motion to that of the real moving stimuli, this implied motion effect did not influence participants’ timing estimates (see also Vicary et al., 2014, for analogous differences in visual working memory tests of dynamic events being more accurately encoded and retrieved in real as compared to implied motion conditions). This result can be accounted for by the predictions made by the “change model” (e.g., Poynter, 1989), according to which real movement can be considered more informative given the (actual and not implied) spatiotemporal changes, which presumably lead to interval overestimations. Additionally, our results are compatible with the contextual change hypothesis according to which the remembered stimulus’ duration is mediated by the amount of contextual change embedded in the stimulus (e.g., Block, 1978; Block & Reed, 1978). In our case, one can argue that real motion presentations encompass a greater amount of contextual changes in comparison to implied motion and were, thus, judged as lasting longer.
The CV results we obtained yield additional support to this real versus implied motion distinction (i.e., real motion stimuli being more informative might have led to better prediction strategies adopted by the participants), at least in terms of time estimation, with greater response consistency for real as compared to implied motion presentations. Hence, a direct comparison of real and implied motion, at least in the timing domain, tentatively argues for distinct processing mechanisms or differential involvement of the same brain areas for real or implied motion (see Zeki, 1999b, in the context of opt art, arguing that while real movement is registered in both V1 and V5, illusionary movement mainly involves V5 and to a smaller degree V1).
Our data analysis also showed—as originally hypothesized—that real motion stimuli that contain higher amounts of movement—as rated (see Methods)—were judged as lasting longer than stimuli containing lower amounts of movement. This finding is compatible with the theory of magnitude (e.g., Walsh, 2003) and previous studies that corroborated that size, space, and quantity of stimulation are linked to time estimates via a positive relationship (e.g., Rammsayer & Verner, 2014, 2015; Sgouramani & Vatakis, 2015b, 2014; Xuan et al., 2007). According to this framework, the estimation of time, size, spatial displacement, and quantity is based on shared accumulation strategies connected to a single mental magnitude system (Walsh, 2003). A common neuronal substrate (e.g., inferior parietal cortex) for these procedures has been advocated by Walsh supporting further this connection. Our results can also be accounted for by the “change model” framework (e.g., Poynter, 1989), as stimuli with higher as compared to lower amounts of movement inherently contain a greater number of spatiotemporal changes (e.g., leg/arms height) and, thus, interval overestimations occur.
Interestingly, while for the real motion conditions, different modulations of participants’ timing estimates are being reported as a function of the amount of movement involved, a null effect was obtained for implied motion presentations. These findings are in conflict with our expectations and previous literature (e.g., Chen et al., 2014; Nather et al., 2011; Yamamoto & Miura, 2012; but see Moscatelli et al., 2011). We speculate that three key points may shed some light on the source of these inconsistencies. First, some evidence support that the order of stimulus presentation may modulate participants’ timing performance (e.g., Nather & Bueno, 2011; Nather et al., 2015). Thus, contradicting results could be due to presentation order effects, which in our case were controlled for by proper stimulus randomization. Second, the utilization of static images implying motion showing either the starting or end point of the corresponding dynamic event has been previously debated (e.g., Acik et al., 2014; Jellema & Perrett, 2003; Urgesi, Moro, Candidi, & Aglioti, 2006), with no firm conclusions. In our study, given the nature of the movement selected (i.e., dance step), it was thought to be depicted better at the peak of the action, which happens to be the end posture (see also Dobrez, 2013, on the role of asymmetry in still images depicting movement). Our choice is in accordance with other studies that have also utilized the end posture and have reported an effect on timing estimates (e.g., Bueno & Nather, 2012; Nather & Bueno, 2011, 2012a; Nather et al., 2011). Third, the naturalistic form of our displays, presenting real biological motion as compared to images of ballerinas-statues or computer-generated images (e.g., Nather et al., 2011; Yamamoto & Miura, 2012; see also Watanabe, 2008, for different timing effects when comparing biological, scrambled biological, and object motion) could have led to the obtained null effect. That is, the latter type of stimuli, even though complex and indicative of biological motion, lack the naturalistic context that the former stimuli contain (see also Moscatelli et al., 2011). In addition, the utilization of actual biological motion is critical, as it has been considered as a special kind of stimulus with social relevance (e.g., Benton, Thirkettle, & Scott-Samuel, 2016). Furthermore, in terms of the notion of independent temporal processing mechanisms for animate as compared to inanimate movement (e.g., Carrozzo, Moscatelli, & Lacquaniti, 2010; Orgs & Haggard, 2011; Wang & Jiang, 2012), one could argue that implied motion pictures of a dancer (i.e., animate)—the stimuli used in the present study—in comparison to ballerinas-statues (i.e., inanimate)—the stimuli used in past studies—could modulate our timing percepts, thus explaining our contradiction to the findings of previous studies (e.g., Bueno & Nather, 2012; Nather & Bueno, 2011, 2012a; Nather et al., 2011).
To conclude, directly comparing, for the first time in the timing domain, the effect of real and analogous implied motion resulted in an overestimation of the intervals tested for the former as compared to the latter ballet steps. This finding implies potential differential processing mechanisms or differential involvement of the same brain areas (e.g., V1 and V5) for real and implied motion in timing. Furthermore, the amount of movement contained in a stimulus was found to influence participants’ performance, but only when real motion was presented. These results corroborate the potential real and implied motion distinction in timing. More studies are needed to further verify this distinction. These studies should explicitly test static body postures of both the starting and end point of an event, include static images without implied motion as an additional condition, and further explore the previously reported presentation order effects. The efficient depiction of a dance act via a static image and the thorough investigation of the correspondent dynamics involved for both implied and real dance versions, apart from the timing domain could also promote the emerging field of neuroaesthetics (e.g., Calvo-Merino, Urgesi, Orgs, Aglioti, & Haggard, 2010; Di Dio et al., 2016; Zeki, 1999a; Zeki & Ishizu, 2013).
Footnotes
Acknowledgements
This research was part of HS’s doctoral thesis work, which was funded by the Onassis Foundation. We would like to thank all the participants that took part in the experiments. We express our gratitude to Ms. Tatiana Myrkou and Mr. Haris Farros for their assistance in the recording of the stimuli and Ms. Foteini Pavlopoulou for allowing us to use the dance studio where all the recordings took place. Part of this project was presented as a talk at the International Multisensory Research Forum, 13–16 June 2015, Pisa, Italy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.