
Abstract
In the present study, we examined the effects of the other’s triadic attention to objects on visual search performances in chimpanzees. We found the search-asymmetry-like effect of the other’s attentional state; the chimpanzees searched a target object not attended by the other individual more efficiently than that attended (Experiment 1). Additional experiments explored the possibility that the other individual “holding an object but not looking at it” led to expectancy violation (Experiment 2) or the role of nonsocial cues such as the proximity relation between the head and the object (Experiment 3). Still, these accounts alone did not explain this effect. It was also shown that the other’s attentional state affected the chimpanzees’ performances more readily as the interference effect than the facilitation effect (Experiment 4). Furthermore, the same effect was observed in the visual search for the gaze (head direction) of others (Experiment 5). We obtained the same results using photographs of chimpanzees (Experiment 6). Contrary to the chimpanzees, humans detected the object to which attention was directed more efficiently than vice versa (Experiment 7). The present results may reflect species differences between chimpanzees and humans in processing triadic social attention.
We humans can recognize the attentional state of others based on the direction of their gaze, head, body, pointing, and other actions and utilize this social information in various ways (Moore & Dunham, 1995; Stephenson et al., 2021). Comparative studies on understanding of the other’s attentional state have also been conducted in nonhuman primates (Emery et al., 1997; Hare et al., 1998; Johnson & Karin-D’Aarcy, 2006; Okamoto et al., 2002, 2004; Tomasello, 2022; Tomasello & Call,1997; Tomonaga, 2010) and other nonhuman animals such as dogs, cats, dolphins, and so on (Koyasu et al., 2020; Pack & Herman, 2006; Siposova & Carpenter, 2019).
There are two main types of interactions in which social attention operates. The first is called dyadic interaction, based on the relationship between the self and the other. The second is triadic interaction, based on the interexchange of attention between the self, the other, and the object (cf., Striano & Stahl, 2005). Dyadic social attention has been studied in relation to the sensitivity to eye contact or direct gaze. For example, humans prefer facial stimuli from birth and can discriminate between faces with direct and averted gaze by 4 months of age (Vecera & Johnson, 1995; see also Farroni et al., 2002). This direct-gaze preference has also been reported in infants of chimpanzees, gibbons, and macaques (Myowa-Yamakoshi & Tomonaga, 2001; Myowa-Yamakoshi et al., 2003; Simpson et al., 2019). In the studies using a cueing task (or dot-probe task) in which a front face was presented as a precue in human adults, detection of the target stimulus at the cued location was facilitated (Bindemann et al., 2007). The same results were reported in chimpanzees (Tomonaga & Imura, 2009a; Wilson et al., 2018). In addition, humans detected direct-gaze faces more efficiently than averted-gaze faces in visual search tasks (stare-in-the-crowd effect; von Grünau & Anston, 1995; Senju & Hasegawa, 2005). Again, chimpanzees also searched for direct-gaze faces more efficiently than averted-gaze ones (Tomonaga & Imura, 2010, 2015). Dyadic interactions based on mutual gaze develop in chimpanzee mothers and infants around 2 months of age (Bard et al., 2005; Tomonaga et al., 2004). Taken together, dyadic social attention is processed similarly in humans and chimpanzees.
On the other hand, there are critical differences in triadic social attention between humans and chimpanzees (Tomasello et al., 2005; Tomonaga et al., 2004). As mentioned earlier, nonhuman primates, including chimpanzees, routinely follow the gaze direction of others (e.g., Itakura, 2004; Tomasello et al., 1998). However, they do not show “joint attention” as shown by human infants from 9 months, in which infants make eye contact with others, follow the gaze of others, and jointly attend to the objects (Okamoto et al., 2004; Striano & Stahl, 2005; Tomonaga et al., 2004). Chimpanzees also reach for objects manipulated by their mothers (joint engagement), observe tool-using behaviors of mothers and siblings close, and attend to their mothers eating novel foods (Biro et al., 2006; Hirata & Celli, 2003; Kosugi et al., 2003; Ueno & Matsuzawa, 2005). Although attention to these goal-directed behaviors of others is a component of joint attention, the patterns of attention are different from humans. Myowa-Yamakoshi et al. (2012) used an eye tracker to observe where chimpanzees and humans pay attention to the goal-directed behaviors of others. Both humans and chimpanzees showed predictive looking at the goal of the behavior, but in humans (especially 1-year-olds), the gaze moved back and forth between the goal object and the actor’s face. In contrast, chimpanzees looked at the face of the actor only very briefly and paid more attention to the goal object or the action itself. This result is consistent with the lack of joint attention during joint engagements with objects between mothers and infants of chimpanzees (Kosugi et al., 2003).
Among the effects of others’ attentional states on behavior, there have been many empirical studies on attentional shifts triggered by social cues in humans. Nonsocial peripheral cues cause reflexive shifts of attention (fast activation and tolerance to low predictability), whereas central cues such as arrows cause voluntary shifts (relatively slow activation and high sensitivity to cue predictability; Muller & Rabbitt, 1989; Posner, 1980). In addition, discrimination and identification of attended objects are facilitated (Downing, 1988; Handy et al., 1996; Kravitz & Behrmann, 2008; Yeshurun & Carrasco, 1999). Social cues, such as eye gaze, face orientation, and pointing, trigger reflexive shifts of attention in humans (Driver et al., 1999; Frischen et al., 2007; Friesen & Kingstone, 1998; Langton & Bruce, 1999). On the other hand, in chimpanzees, the attentional shift triggered by eye gaze, head direction, and reaching gesture occurs slowly and is influenced by cue predictability, regarded as voluntary orienting (Tomonaga, 2007b, 2010a, 2010b; Tomonaga & Imura, 2009b). Thus, their social attention may operate differently from humans.
In the present study, we investigated how the attentional states of others (social attention) affected the observer’s searching behavior using visual search tasks. When chimpanzees and humans search for an environment, they are influenced by the attentional states and interaction patterns of others, such as eye gaze. However, there have not been many studies examining this issue in humans in laboratory settings, and none from a comparative cognitive perspective. Humans showed faster search response times when the congruent gaze cue was presented before the search display (Kuhn & Tipples, 2011; Kuhn et al., 2015) and better change detection by the gaze cues (Langton et al., 2006). In contrast, the facilitation effect of the gaze cue was weakened when embedded in the search display (Hermens, 2017).
Compared to these studies on the effects of dyadic gaze, there are very few studies on those of triadic gaze (e.g., Seow & Fleming, 2019). In the present study, therefore, we conducted visual search tasks in which several photographs of a person (or chimpanzee) holding an object and looking at or away from it were presented as a search display. Chimpanzee and human participants were trained or instructed to select a target photograph in the display. In all experiments, except for Experiment 5, the target stimulus was a specific object (a red can), regardless of the direction of the person’s gaze in the photograph. By combining two attentional states of the persons (Directed and Averted states) in the photographs, we prepared two target/distractor conditions in these experiments. The first was the Directed condition in which the target was the red can held and looked at by the person, and distractors were the other objects held but looked away. The second was the Averted condition in which the target was the red can held but looked away by the person, and distractors were the other objects held and looked at (see Figures 1 and 2).
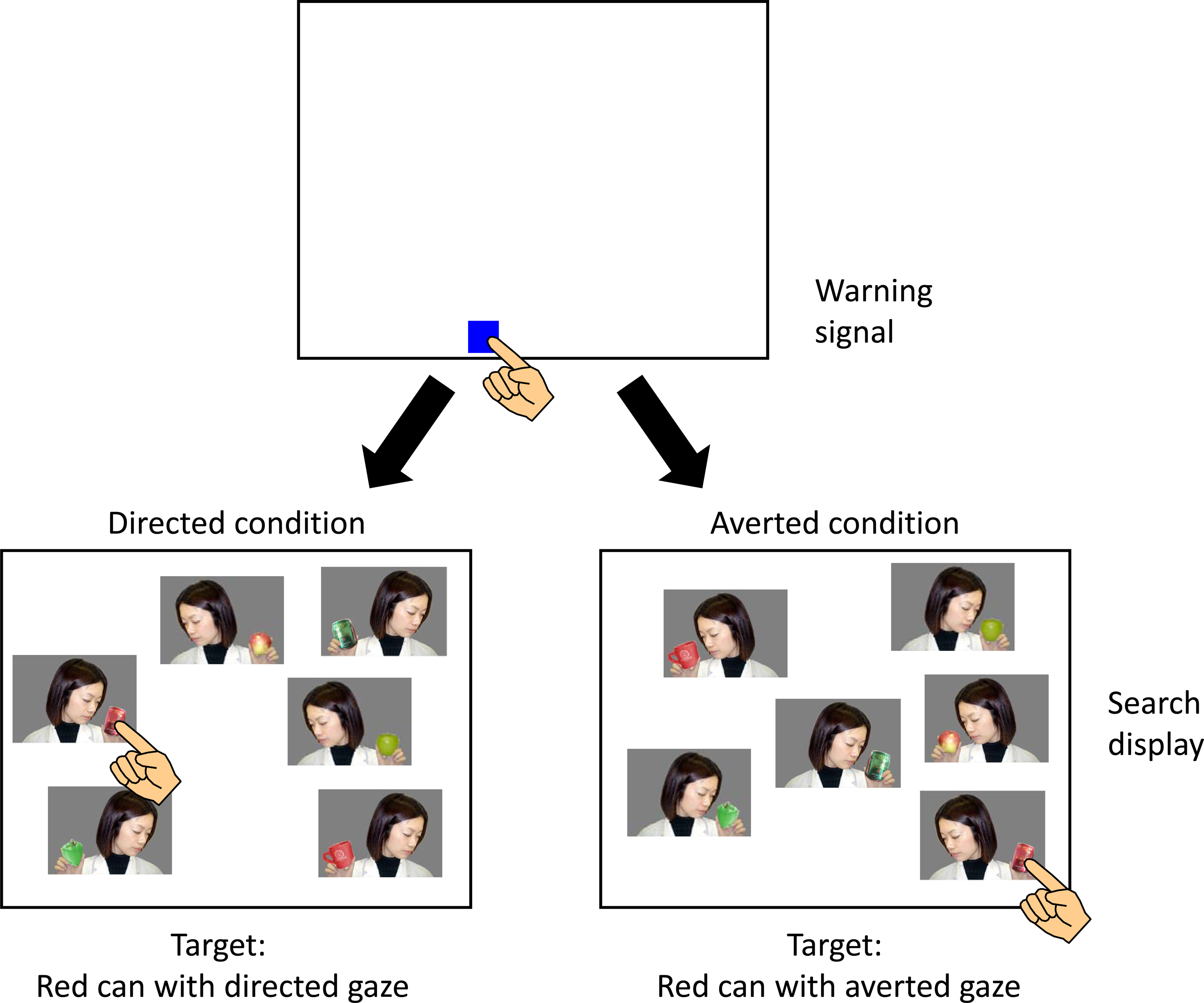
Schematic illustration of the flow of the trial. Note that the search displays are not to scale.
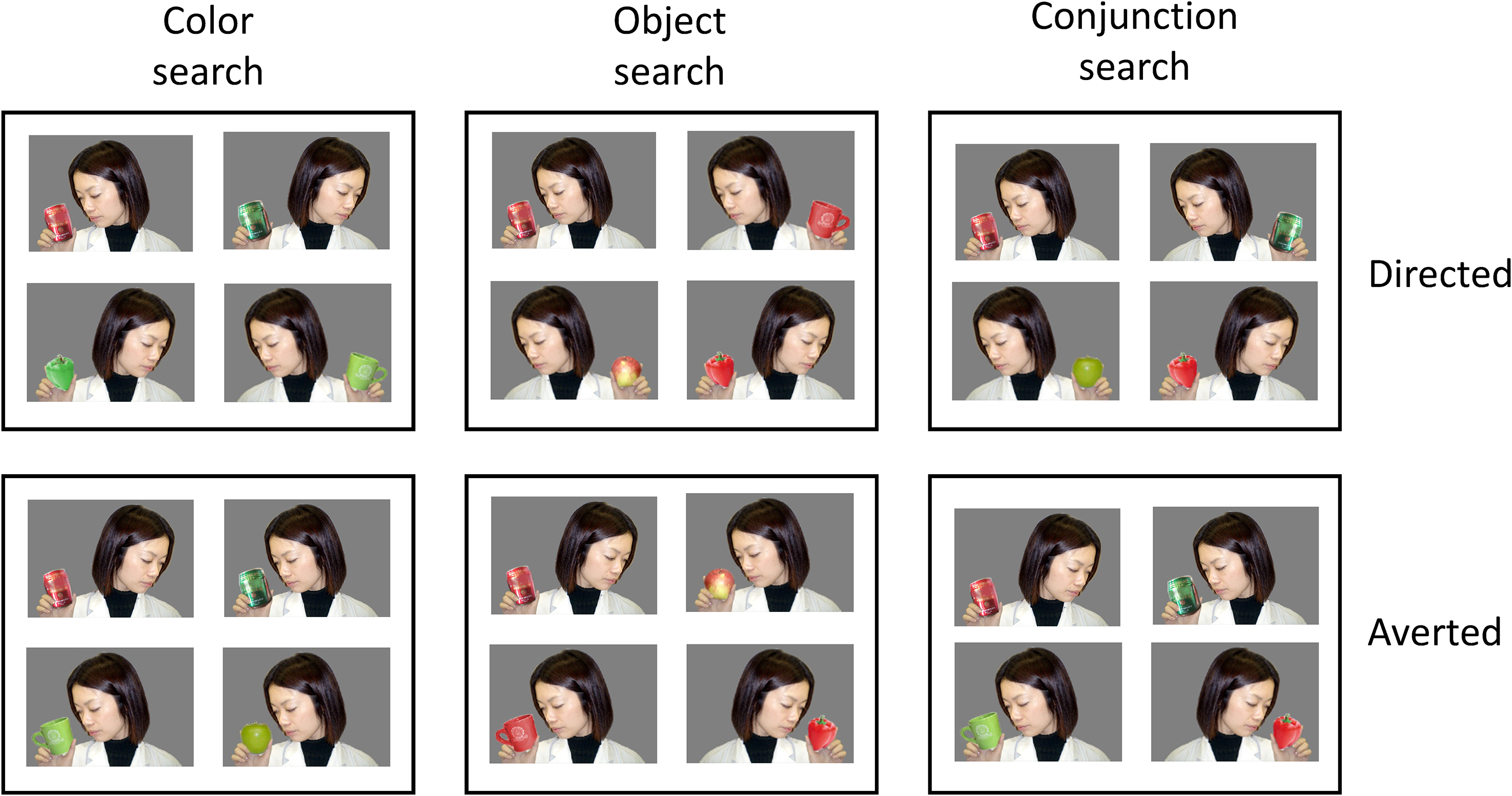
Search types and target/distractor conditions used in Experiment 1 (search displays not to scale).
Based on the previous studies, two types of competing results would be predicted. Firstly, the participants would search more efficiently for the object in the Directed condition than in the Averted condition. This prediction is based on evidence from the studies on joint attention/gaze-following, attentional shift by gaze cues, and attention to goal-directed actions. Since the observer’s attention is initially directed to the object to which the other person’s attention is directed, its processing would be facilitated. As a result, if the object is a target stimulus, it would be efficiently detected.
On the other hand, the second prediction is that the Averted condition is more efficient than the Directed condition. Holding an object but not looking at it is a non-goal-directed behavior. If the observer has prior knowledge or belief that “if an agent has an object, he/she should pay attention to it,” then “holding an object but not looking at it” would violate the observer’s expectancy (cf., Myowa-Yamakoshi et al., 2015; Woodward, 1999). Violation of expectancy attracts an observer’s attention (Stahl & Kibbe, 2022). In human infants, looking times increase when the goal of the repeatedly presented action changes (Biro & Leslie, 2007; Woodward, 1998). Attentional capture caused by such violation of expectancy may facilitate the detection of objects in the scene.
In the present study, we tested chimpanzees and humans on visual search performances under the abovementioned conditions. Based on the species differences in triadic social attention in humans and chimpanzees, species differences may also be observed in this study. First, we examined the effect of target/distractor conditions in Experiment 1. We additionally tested the expectancy-violation account in Experiment 2 and the role of nonsocial cues, for example, spatial proximity between the head and the object, in Experiment 3. In Experiment 4, we compared the facilitation and interference effects of the attentional states of the others in the photographs. In Experiment 5, we further examined the effects of the attentional states of the others on the visual search for gaze directions (head directions) instead of the search for objects (cf., Tomonaga & Imura, 2010, 2015). Experiment 6 tried indirect replications with new stimuli, including new chimpanzee participants, 9 years after the first experiment to verify the replicability of the results. Finally, in Experiment 7, we tested human participants in identical settings to chimpanzees.
Experiment 1
In Experiment 1, we conducted visual search tasks for an object using photographs in which a person held an object and looked at it and those in which a person held an object but did not look at it. We compared search performances by chimpanzees in two conditions: one in which the object at which the person looked was the target stimulus, and vice versa.
Methods
Participants
Three juvenile chimpanzees (Pan troglodytes; for age-class categorization, see Goodall (1986), Matsuzawa (2006), and Nishida et al. (2003)) participated in Experiments 1 through 5; Ayumu (Great Ape Information Network (GAIN) ID#0608, https://shigen.nig.ac.jp/gain/), Cleo (GAIN ID#0609), and Pal (GAIN ID#0611). At the onset of the present study, they were 6 years old. They had been participating in the research project on cognitive development in chimpanzees from birth (Matsuzawa et al., 2006; Tomonaga et al., 2003, 2004). They had started a computer-controlled cognitive task at very early infancy, that is, less than 1-year-old (Sousa et al., 2003; Tanaka et al., 2003). Before the onset of the present study, they experienced visual search tasks. Thus, they were familiar with the current experimental setting. They lived in a social group of 14 individuals in an indoor and environmentally enriched outdoor compound (770 m2) at the Primate Research Institute, Kyoto University (Matsuzawa, 2006). No food or water deprivation was used in the present study.
Ethics Statements
The care and use of the chimpanzees adhered to the 2nd edition of Guide for the Care and Use of Laboratory Primates of the institute. The experimental designs of the present study with chimpanzees were approved by the institute’s Animal Welfare and Animal Care Committee (06-1433). All procedures also adhered to Guideline of the Animal Experimentation of the Japanese Society of Animal Psychology, Guideline for the Care and Experimental Use of Captive Primates of the Primate Society of Japan, Code of Ethics and Conduct of the Japanese Psychological Association, and Japanese Act on Welfare and Management of Animals.
Apparatus and Experimental Setting
All the experiments were conducted in a booth (1.8 × 2.15 × 1.75 m) in a laboratory adjacent to the chimpanzee facility. The mother–offspring pair of chimpanzees came to the booth through the overhead pathway connecting the facility to the booth. Two sets of 17-inch LCD monitors (I-O Data LCD-AD172F2-T, 1280 × 1024 pixels, pixel size: 0.264 mm × 0.264 mm) with touch panels were installed on the booth wall. The viewing distance was approximately 40 cm. A universal feeder (Biomedica BUF-310) outside the booth supplied food rewards (small pieces of apple). The computer controlled all equipment and experimental events.
Stimuli
We used color photographs in the present experiment. Each photograph contained a person and an object. We prepared four objects in two colors; a can, a mug, an apple, and a bell pepper, colored red or green. Two persons familiar to the chimpanzees were also prepared. As shown in Figure 1, the stimuli were categorized into two conditions based on the attentional state of the person in the photograph: the “Directed” state in which the person held an object and looked at it and the “Averted” state in which the person held an object but did not look at it. The object was placed at either left or right of the photograph. These photographs were retouched on a gray background using Adobe Photoshop®. The size of each photograph was 213 pixels in height and 300 pixels in width. The target stimuli were photographs containing a red can.
We also prepared stimuli for the preliminary training in which an object alone was placed on a gray or random-dot background without a person.
Procedure
A typical trial flow for each experiment with chimpanzees was as follows (Figure 1). After 2 s of the intertrial interval, a warning signal (blue square, 90 × 90 pixels) was presented randomly at the bottom of the monitor on a white background with a sound. When the chimpanzee touched it, it disappeared, followed by the presentation of the search display. The search display contained one target and several distractors. These stimuli appeared randomly at 3 × 3 possible locations. The number of stimuli in the search display (called set size) was randomly changed from trial to trial. When the chimpanzee touched one of the stimuli, all stimuli on the screen disappeared. If the chimpanzee touched the target, a chime was sounded, and a food reward was delivered. On the other hand, if a distractor was selected, a buzzer was sounded. If no responses occurred until 8 s passed, a click sound was given, and the screen was turned off, then shifted to the next trial. These trials were treated as incorrect trials. Generally, one session was conducted per day for a maximum of five days per week.
Preliminary Training
Before the main experiment, the chimpanzees were given preliminary training for learning a target stimulus defined by the experimenter. In the first pretraining phase, one session consisted of 96 trials, and all sessions used photographs of objects without a person placed on a gray background. The set size was fixed at 4. The positions of the target and distractors were randomized from trial to trial. Four search types were prepared to control the task difficulty: two color search types (one in which only the green cans were presented as the distractors, and the other in which all the distractors were green but the objects were different from each other), the object search in which all the distractors were red but the objects were different, and the conjunction search in which the colors and objects of the distractors were different from each other. These search types were given to the chimpanzees in random order. Each chimpanzee performed 15 sessions of this pretraining. Each chimpanzee showed 97.9% of correct trials in the last session of the conjunction search condition, and the mean response time for correct trials was 778 ms, 743 ms, and 956 ms for Ayumu, Cleo, and Pal, respectively.
In the second phase, the background was changed to a random-dot pattern. The three search types appeared randomly within one session of 72 trials: color search with different object distractors, object search, and conjunction search. Set sizes of 4, 6, and 8 were presented in random order. The chimpanzees received 6–9 sessions of this pretraining (7 sessions for Ayumu, 9 for Cleo, and 6 for Pal, respectively). The accuracies for the last session were 95.8% (mean response time for correct trials = 857 ms) for Ayumu, 95.8% (740 ms) for Cleo, and 98.6% (1003 ms) for Pal, respectively. After these preliminary training sessions, the chimpanzees were shifted to the main experiment.
Main Experiment
The main experiment used two types of photographic stimuli in which a person held an object. Two target/distractor conditions were prepared as described in the introduction (Figure 2); the Directed and the Averted conditions. Three search types were prepared as in the second phase of the preliminary training (Figure 2). The target/distractor conditions varied from session to session, and the search types and set sizes (4, 6, and 8) changed within sessions. The persons in the photographs in the search display were the same person (see Figure 2), but two different persons appeared randomly in each trial. The left-right position of the object in each photograph and the spatial arrangement of the stimuli also changed randomly for each trial. Each session consisted of 72 trials, and four sessions per target/distractor condition were given to each chimpanzee.
Data Analysis
In the present study, we mainly used the response times for data analyses. The response times were defined as the duration between the onset of the search display and the participant’s response. We used only the data from correct trials. The response times shorter than 200msec were also excluded from analyses (however, only one trial was correct among these excluded trials throughout the experiments). We applied Generalized Linear Mixed Model (GLMM) analyses. We selected a model with the smallest Akaike Information Criterion (AIC) among all possible models. We also additionally evaluated whether parameter estimates (or coefficients) of the selected models differed significantly from zero or not, using p-values (alpha level was set at .05) and confidence intervals based on Wald test statistics. We applied inverse Gaussian distribution with the identity link function for each model. We initially prepared all possible random intercept models with participant and session (nested with participant) as random effects. Fixed effects were target/distractor condition, search type, and set size. These analyses were performed using lmerTest (Kuznetsova et al., 2017) and MuMIn (Bartoń, 2022) packages in R version 4.2.0 (R Core Team, 2022).
The Supplementary File shows the raw data for each participant in all experiments. The results for all GLMMs in each experiment are shown in Supplementary Table S1.
Results and Discussion
Chimpanzees’ performances were accurate and stable throughout the present study (Table 1). The mean percentage of error trials throughout the present study was 2.7% (the standard error of means (SEM) across experiments was 0.7). No further detailed analysis of the errors was conducted in this study.
Mean percentage of errors for each experiment.
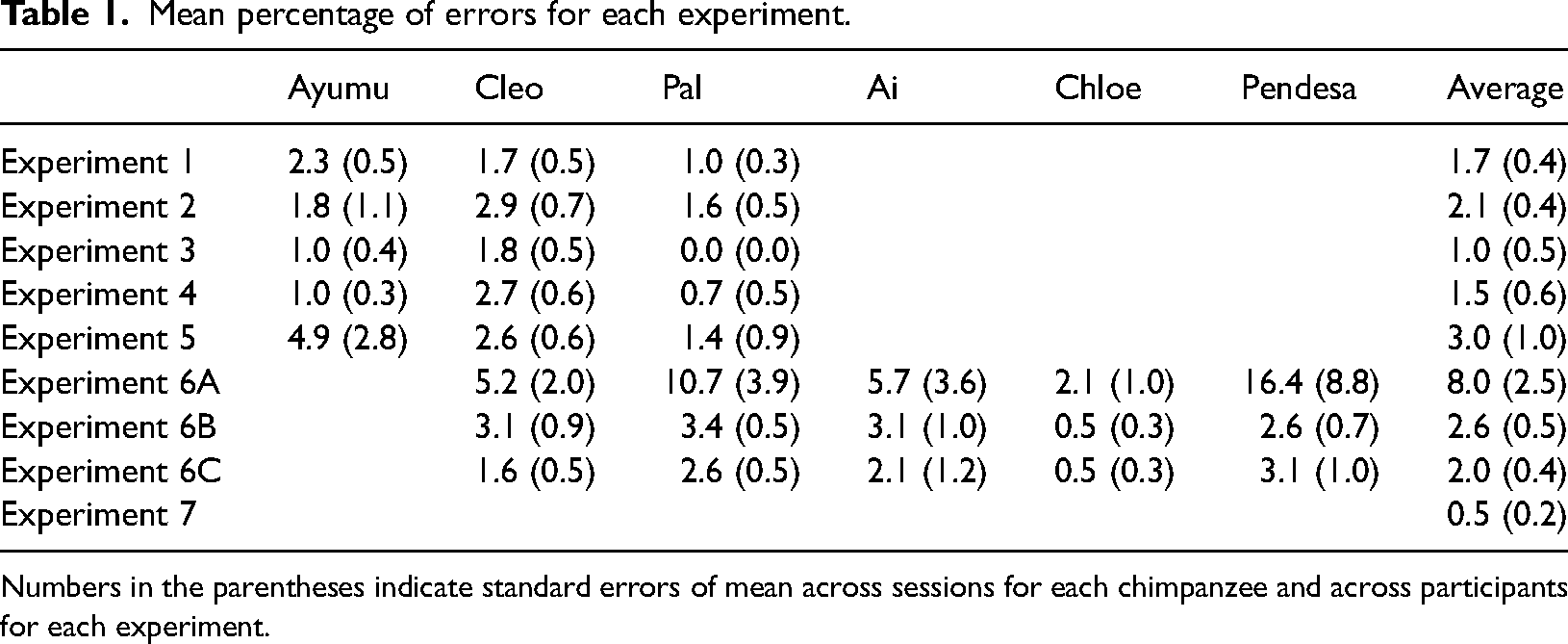
Numbers in the parentheses indicate standard errors of mean across sessions for each chimpanzee and across participants for each experiment.
Figure 3 shows the mean response times for correct trials for each search type and target/distractor condition. Table 2 presents a summary of the selected model. The model selection process is shown in Table S1. As shown in Figure 3, the Averted condition was more efficient than the Directed condition in all search types, indicating the search-asymmetry-like phenomena. The selected model included set size (β = 39.76, 95% confidence interval [24.69, 54.82], p < .001), target/distractor condition (β = −86.03, [−122.27, −49.78], p < .001), search type, and the interaction between set size and search type as fixed effects. Note, however, that the Wald confidence intervals for the coefficients of the search type include 0 (Table 2). For the interaction between set size and search type, the search slope (=effect of set size) in the color search was shallower than in object and conjunction search types.
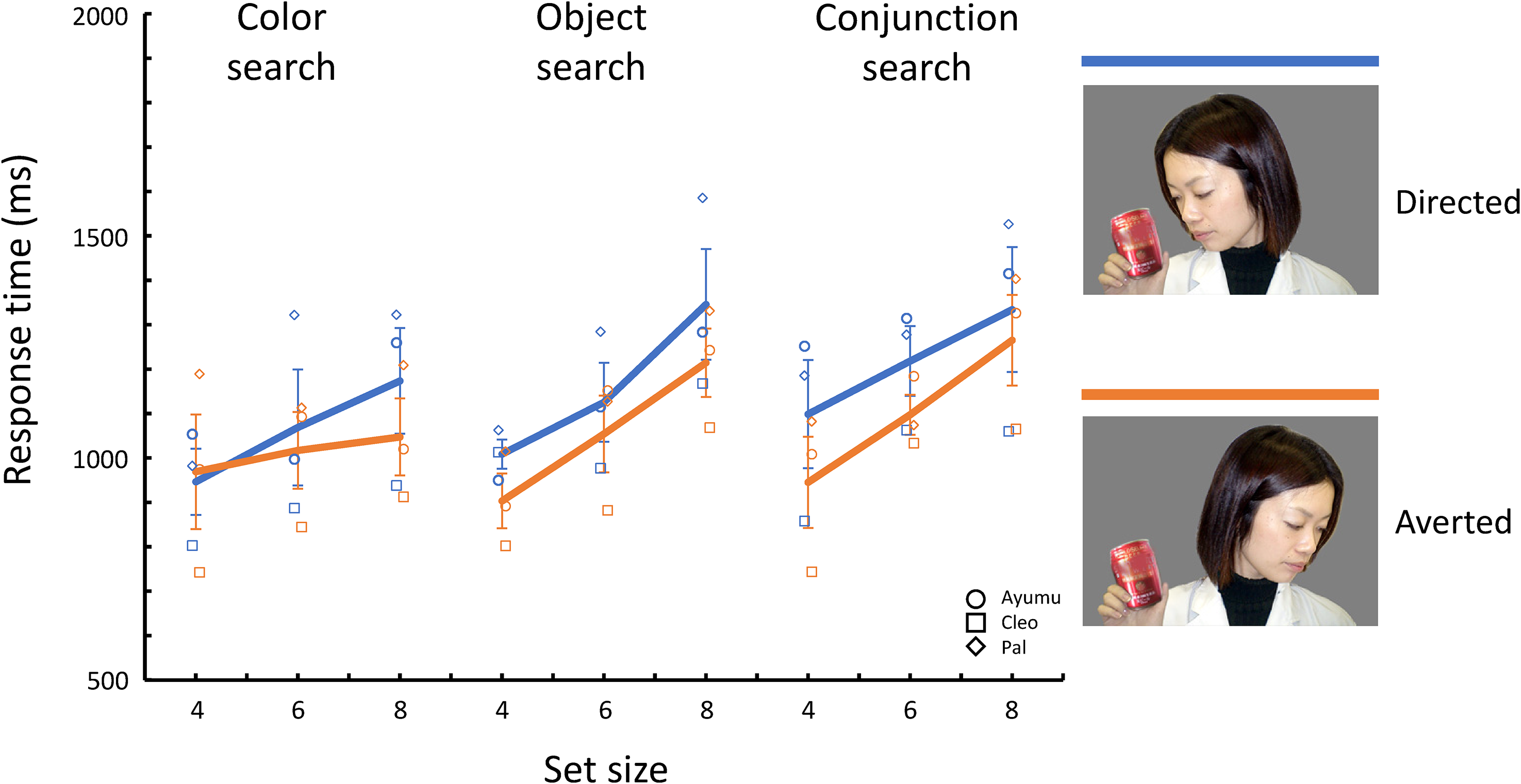
Mean response times for correct trials for each search type and target/distractor condition in Experiment 1. Symbols indicate each chimpanzee’s data. Solid lines indicate averages across chimpanzees. Error bars indicate the standard error of the mean across chimpanzees.
A summary of the selected models for each experiment.
For each categorical variables (TGT/DST and Search type), the base and the other levels are shown.
TGT/DST: target/distractor condition.
D: directed gaze to the object, A: averted gaze to the object, F: face in front.
“:” denotes interaction.
SE: standard error, CI: confidence interval, SD: standard deviation.
Experiment 1 indicated that “objects in hand but not attended to” were detected more efficiently in the chimpanzees than “objects in hand and attended to.” These results are not consistent with the prediction based on joint attention and gaze-triggered attentional shifts but consistent with the prediction based on the violation of expectancy: the state “holding an object but not looking at it” may be more likely to capture chimpanzees’ attention.
Experiment 2
If “holding an object but not looking at it” violated the chimpanzee’s expectancy or prediction, then there might be no difference in search performance between target/distractor conditions when the objects were also placed on both sides (see Figure 4). Experiment 2 was conducted under these conditions to examine the effects of expectancy violation.
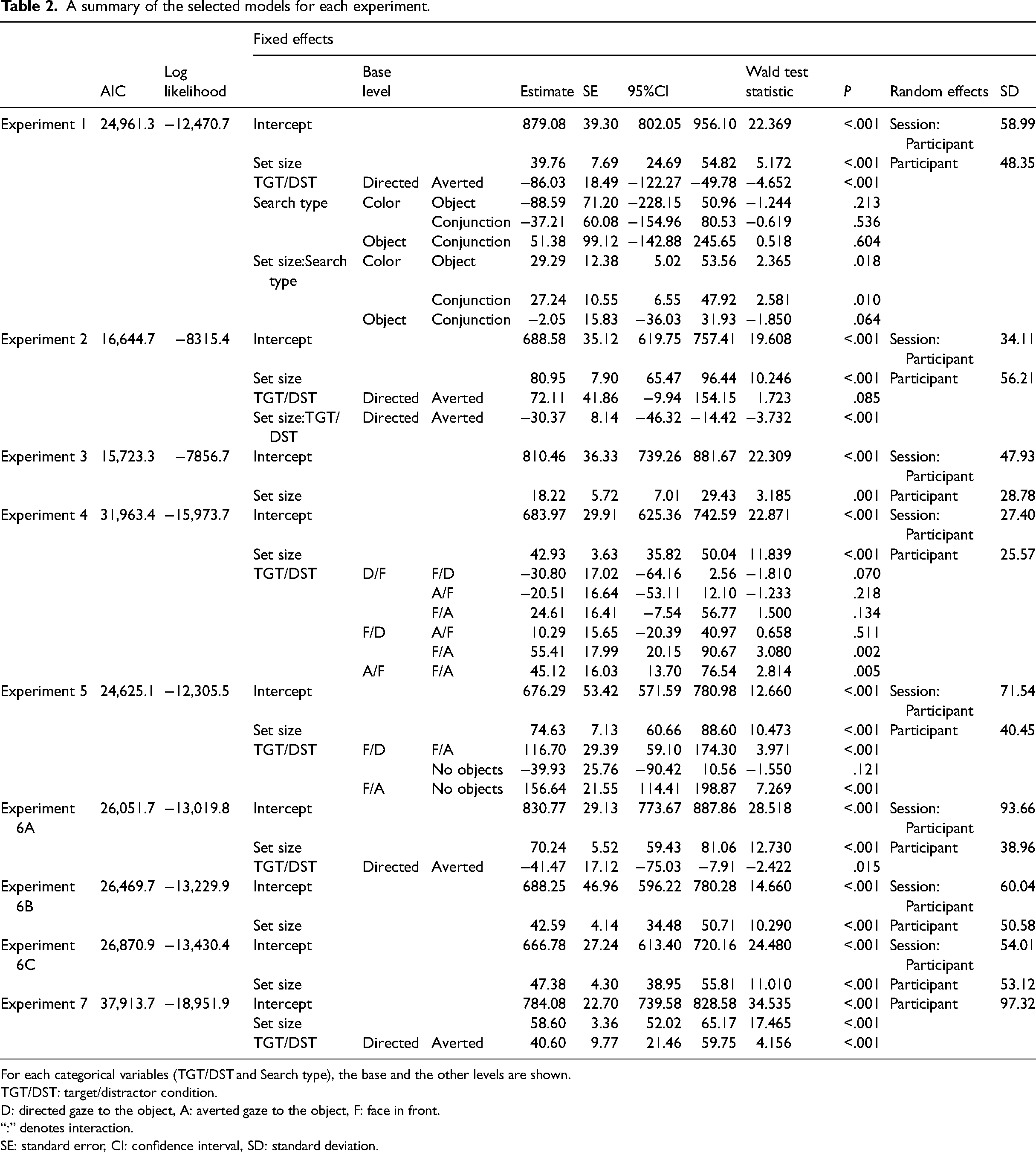
Mean response times for correct trials for each target/distractor condition in Experiment 2.
Methods
In Experiment 2, as shown in the right part of Figure 4, the person in the photograph held objects in both hands. As in Experiment 1, the person held one of the eight objects with one hand, while the person had a ball (common to all photographs) with the other hand. Therefore, even in the Averted photographs, the person “ looked at the object she held.” As in Experiment 1, the photographs were created by retouching using Adobe Photoshop®.
As in Experiment 1, the same three chimpanzees participated in the experiment. Only the conjunction search type was conducted in the present and subsequent experiments. The number of trials per session was 48. The other conditions were the same as in Experiment 1. Four sessions were conducted for each target/distractor condition.
Results and Discussion
Figure 4 shows the mean response times for correct trials for each target/distractor condition. Table 2 presents a summary of the selected model, and the model selection process is shown in Table S1. The selected model included set size (β = 80.95, [65.47, 96.44], p < .001), target/distractors condition, and their interaction as fixed effects. Although the Wald confidence interval for the parameter estimates of the target/distractor condition included 0 (β = 72.11, [−9.94, 154.15], p = .085), the search slope was shallower in the Averted condition than in the Directed condition (β = −30.37, [−46.32, −14.42], p < .001), indicating the search was more efficient in the Averted condition than in the Directed condition as in Experiment 1.
In Experiment 2, the search performances were examined using photographs in which the person held an object in each hand, and whichever way she turned, she was “looking” at one of the objects. It was to eliminate the violation of expectancy caused by the state of “holding an object but not looking at it.” The results reconfirmed the search-asymmetry-like effect that the Averted condition was more efficient than Directed as in Experiment 1 and suggested that a simple expectation violation account alone is unlikely to explain the present results. However, we cannot rule out the possibility that the chimpanzees paid attention only to the person’s attentional state toward the target object, and thus the violation of limited expectancy, that is, “holding a target object but not looking at it,” might capture the chimpanzee’s attention. If so, the “interference effect” of the other’s attentional state in the distractors may be less critical than the “facilitation effect” of the target. We address this issue in Experiment 4.
Experiment 3
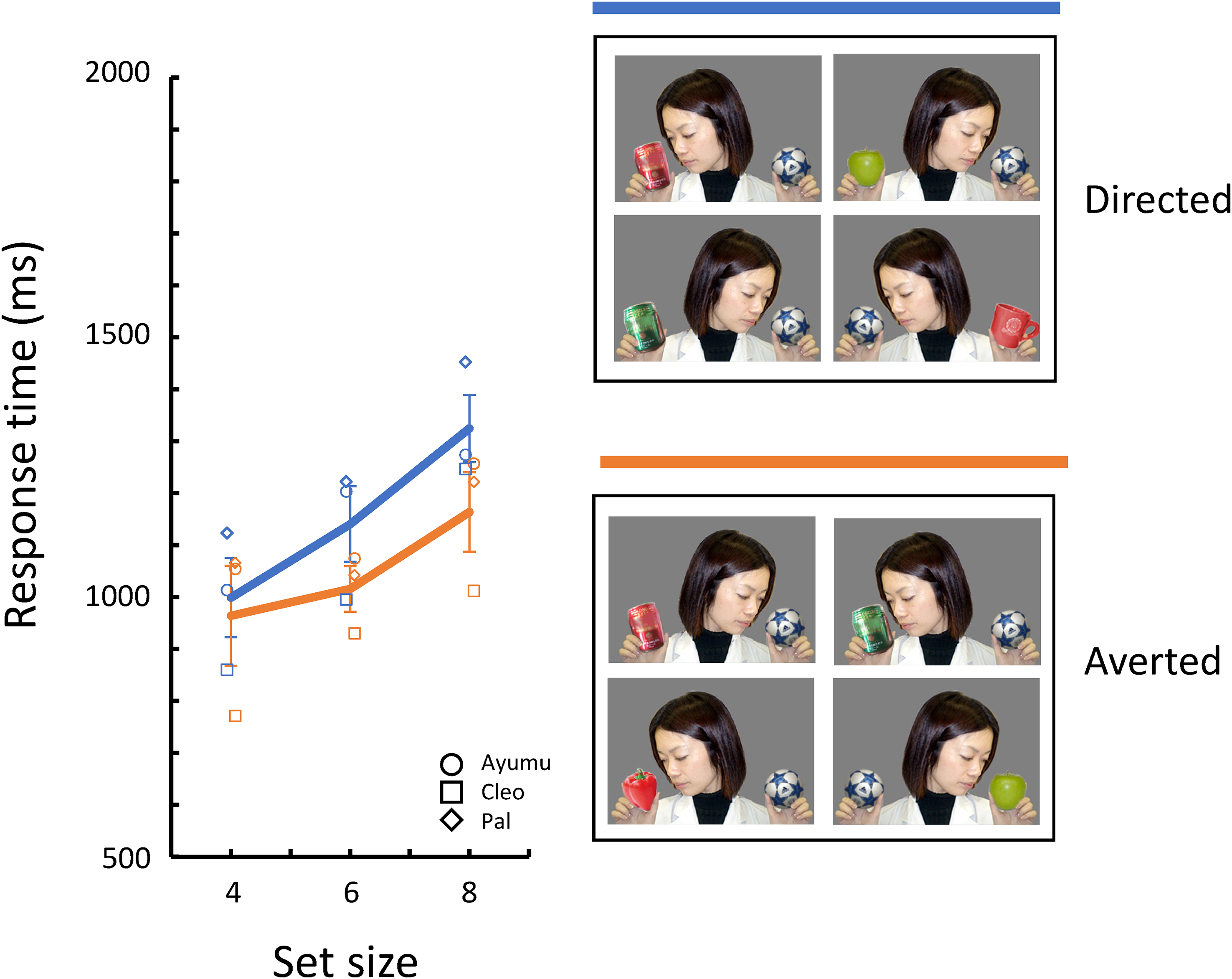
In the Directed photographs used in this study, the person tilted her head toward the object. In the Averted photographs, on the other hand, the head was tilted toward the opposite direction of the object. Nonsocial cues such as the proximity of the head to the object, rather than the attentional state of the other person, might have influenced the chimpanzees’ performance. In Experiment 3, we silhouetted the person in the photographs to remove information about the eye-gaze direction and examined the effect of the silhouette on search performance.
Methods
In Experiment 3, we silhouetted the person in the photographs used in Experiment 1 (see Figure 5). All the other procedures were the same as in Experiment 2.
Mean response times for correct trials for each target/distractor condition in Experiment 3.
Results and Discussion
Figure 5 shows the mean response times for correct trials for each target/distractor condition. The figure shows that, unlike in the previous experiments, there was no difference in the target/disturbance conditions, and the search slope was very shallow The model including only set size as a fixed effect was selected (β = 18.22, [7.01, 29.43], p = .001).
When the stimuli with a silhouetted person were used, the “attentional state” of the person did not affect the object search. The results suggest that nonsocial cues such as head tilt and proximity to the object did not explain the results of the previous experiments. It is known that chimpanzees perceive and recognize various body information from silhouettes (Gao et al., 2018; Tomonaga & Imura, 2023; Tomonaga, 2010b). However, under the conditions of the present experiment, the attentional state signaled by the orientation of the body and head by the silhouette did not affect the chimpanzee’s visual search performances. These results suggest that the information from the inner features of the head removed by silhouetting, namely, the position of the eyes in the head area, may be the most critical for the present task (cf., Perrett et al., 1992).
Although the present experiments indicate that it is difficult to explain the previous results solely in terms of specific nonsocial cues, we still cannot rule out the possibility that other nonsocial cues may still have some effects. For example, the profile of the persons in the photographs used in the present study was brighter in the front (exposed skin) than in the back (covered by hair). The effect of this brightness contrast between the front and rear parts and its spatial relation to the object is further examined in Experiment 6.
Experiment 4
Experiment 2 suggested that if chimpanzees’ search performances were affected by the violation of expectancy, its effect might be more enhanced in the targets rather than in the distractors. However, because the target and distractors in each target/distractor condition (Directed and Averted) were different, it was not possible to examine differences in the effect of the other’s attentional state only for the target or for the distractors. Therefore, Experiment 4 introduced the third set of photographs in which a person looked at the front to fix the target or distractor between the target/distractor conditions.
Methods
In Experiment 4, as shown in Figure 6, three new stimulus sets were created using photographs of two persons (one of whom appeared in the stimuli of the previous experiments). The eight objects remained the same as before. The first set was the “Front” photographs (F), in which a person faced in front, lifting the object to eye level and holding it in her hand. The second set was the “Directed” photographs (D), in which a person looked at an object held in her hand. Unlike the stimuli in the previous experiments, the person in the photograph did not tilt her head toward the object. Finally, in the “Averted” photographs (A), a person looked at the opposite side of an object in her hand. In the previous stimuli, the gap between the head and the object differed from the Directed and Averted photographs since the head was tilted. But in these three sets of photographs, the gap between the head and the object was almost equal. Of the six possible target/distractor conditions, four conditions were prepared. The F/D and F/A conditions had the same stimulus set of targets and different sets of distractors, while the D/F and A/F conditions had the same set of distractors and different sets of targets. These four conditions varied across sessions. The Directed and Averted conditions did not appear in the present experiment. All other conditions were the same as in Experiments 2 and 3.
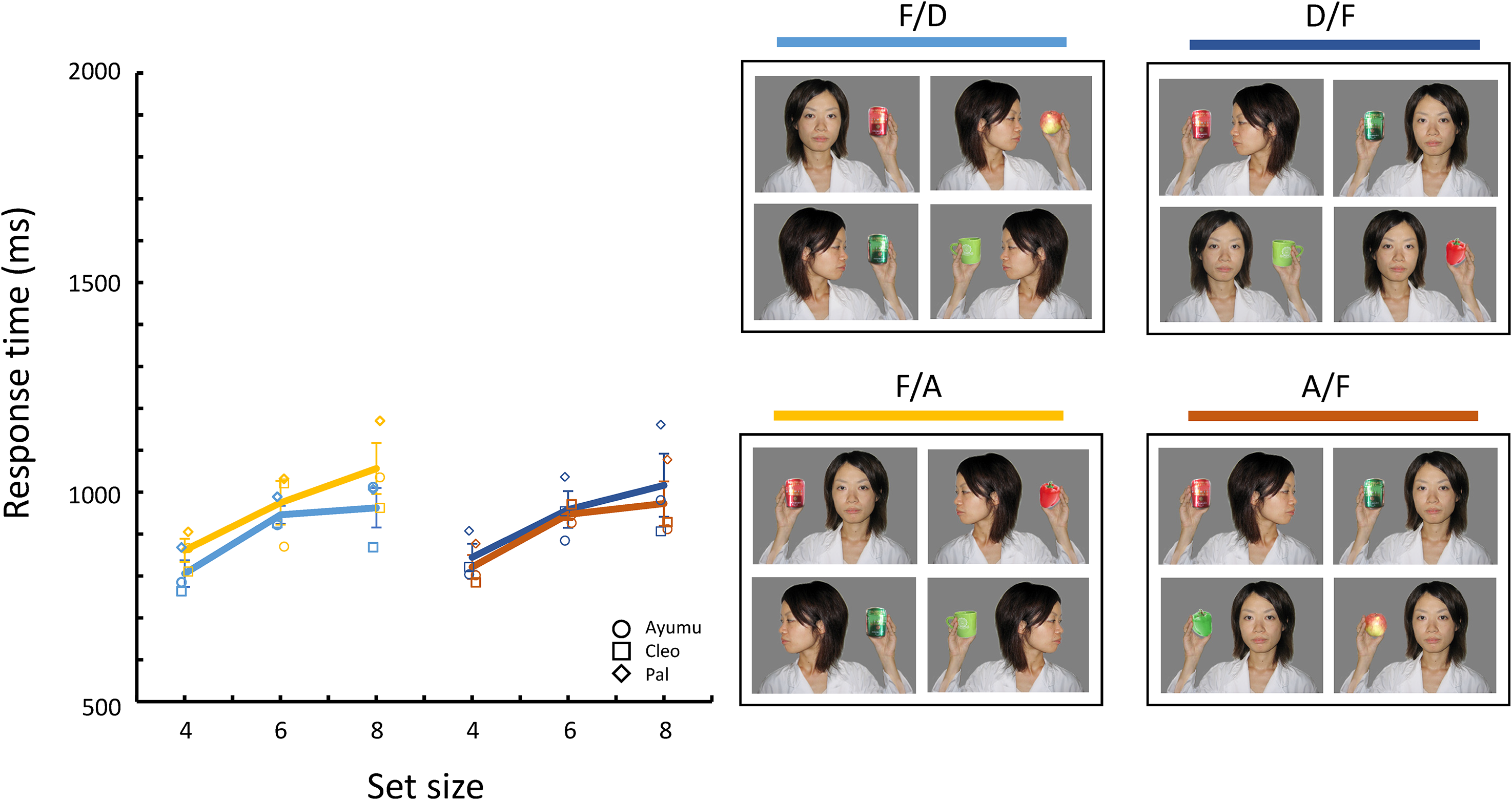
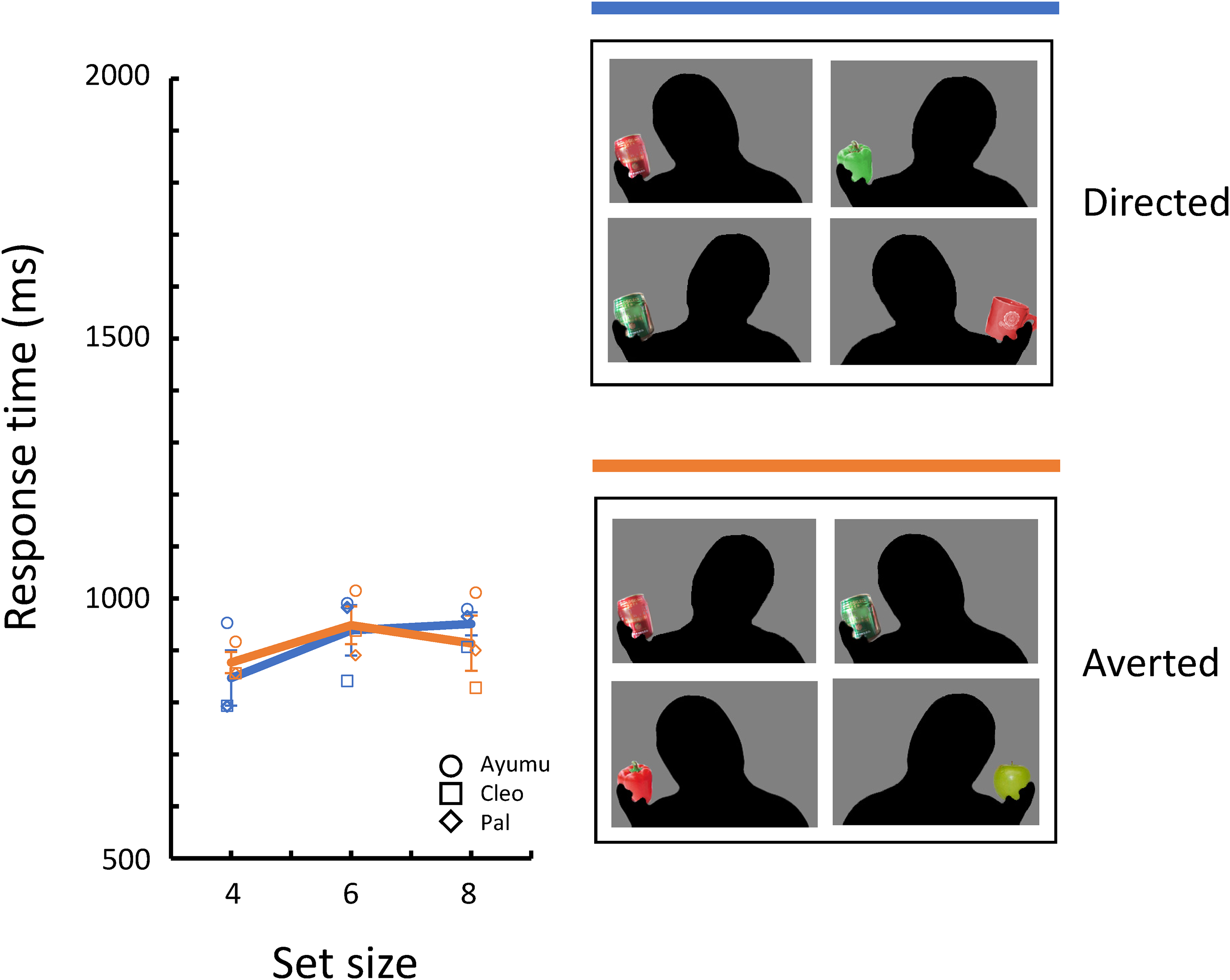
Mean response times for correct trials for each target/distractor condition in Experiment 4.
Results and Discussion
Figure 6 shows the mean response times for correct trials for each target/distractor condition. The two conditions with the Front target (F/D and F/A) are shown on the left side, and the two with Front distractors (D/F and A/F) are shown on the right side. Table 2 also shows a summary of the selected models. The model selection process is shown in Table S1. The selected model included set size (β = 42.93, [35.82, 50.04], p < .001). and target/distractor condition as fixed effects. Detailed inspections based on the Wald statistics (see Table 2) showed clear differences between symmetrical conditions with Averted photographs (F/A and A/F; β = 45.12, [13.70, 76.54[, p = .005) and between conditions in which the target sets were the same (F/D and F/A; β = 55.41, [20.15, 90.67], p = .002), but no differences between the other symmetrical conditions with Directed photographs (F/D and D/F) and between conditions in which the distractor sets were the same (D/F and A/F).
In Experiment 4, we introduced the third set of photographs, the Front photographs including a dyadic gaze, and compared the facilitation and interference effects of the triadic gaze on search performances. The results from the symmetrical target/distractor conditions indicate that the effect of the directed gaze toward the object on search asymmetry was weaker than that of the averted gaze away from it. Furthermore, the effect of the triadic gaze, especially the averted gaze away from the object, was more substantial when it was present as the distractor than as the target. These results may be explained by the difference in the number of the target and distractors in the search display. If the averted gaze captures the chimpanzee’s attention more readily than the directed gaze, then each stimulus with the averted gaze in the search display would capture attention one at a time, resulting in increased search response time. This cumulative interference effect of the Averted photographs may explain the results of the previous experiments.
Experiment 5
In the previous experiments, the chimpanzees searched for a specific object (a red can). We found that the attention to the object held but not looked at by the other person affected the chimpanzees’ visual search performances. If the chimpanzees paid attention not only to the object alone but also to the entire interaction between the person and the object (cf., Myowa-Yamakoshi et al., 2012, 2015), the differences in the other's attentional state would also affect the visual search for head directions instead of search for the objects. To address this question, we trained the chimpanzees to search for a front-facing person with an object among distractors in which a person looked at or away from an object in Experiment 5.
Methods
As shown in Figure 7, the four different stimulus sets were prepared in Experiment 5. Two were pictures of a person facing front and side without an object. The remaining two sets were picked up from the Directed and Averted photographs used in Experiment 4, which were photographs of a person holding a green mug. The two persons appearing in the photographs were the same in Experiment 4. Only the green mug appeared as an object. The target stimulus was always a photograph of a person facing front (Front photographs, F). Three target/distractor conditions were prepared. The first was the No Objects condition as a control condition, in which photographs of a person without an object appeared. The second was the Front/Directed condition (F/D), and the third was the Front/Averted condition (F/A). Previous experiments suggested that attention was more likely to be captured in the Averted photographs than in the Directed photographs. Thus, the chimpanzees would show a less efficient search for the head direction in the F/A than the F/D condition due to the more substantial cumulative interference effect by the distractors.
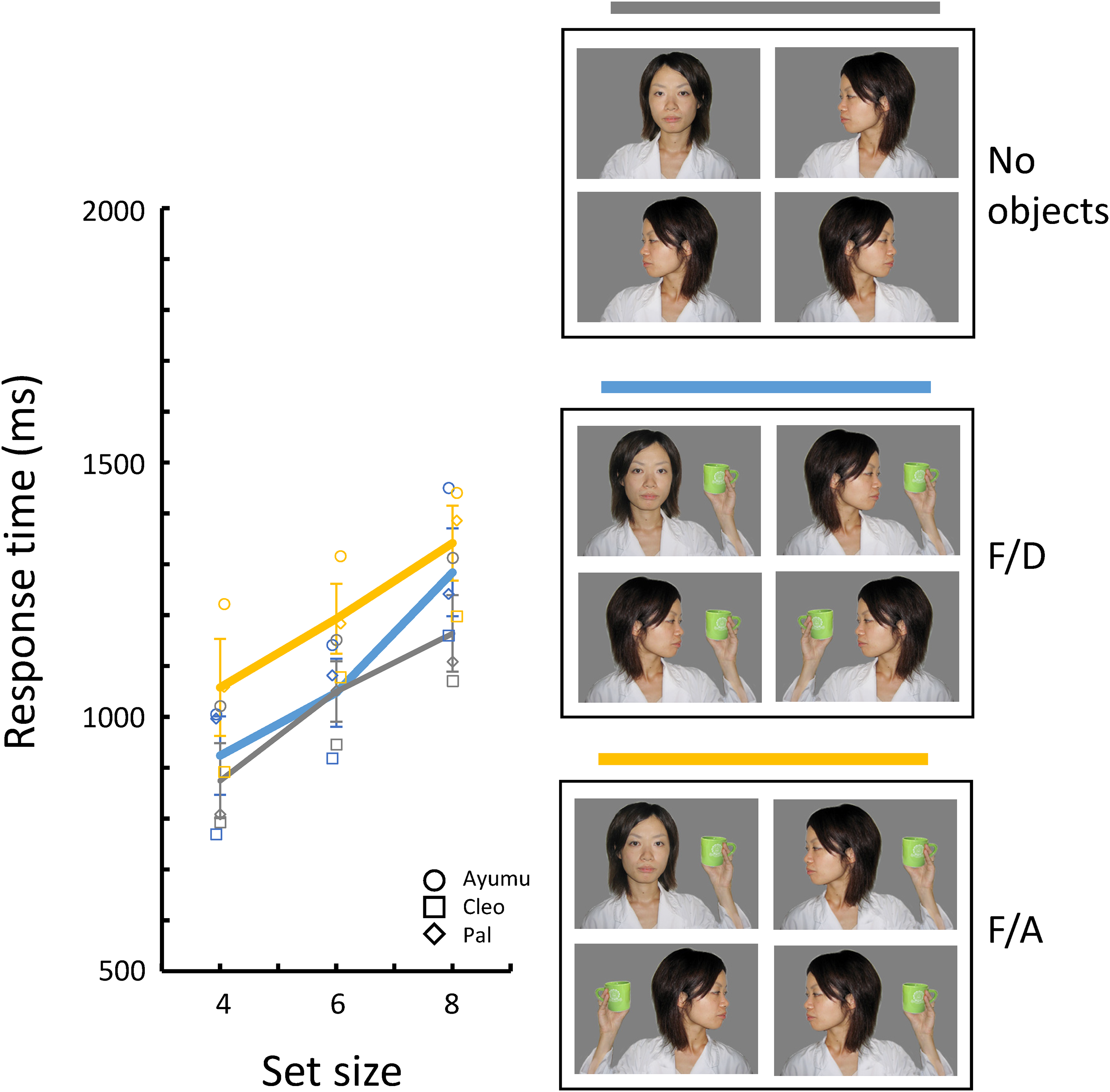
Mean response times for correct trials for each target/distractor condition in Experiment 5.
Since, in the present experiment, the chimpanzees were required to search for head directions, unlike in the previous experiments where the target was the object, we initially introduced preliminary training with photographs in which only the person appeared. Each session consisted of 32 trials, and the set size was 4. The target was a frontal face, and the distractor was a profile. The spatial arrangement of the stimuli in the search display was randomly changed for each trial. We introduced a correction method for incorrect trials, in which the same trial was presented again. If the chimpanzees made a further error on the correction trial, only the target was presented on the next correction trial. Ayumu received 11 sessions of this pretraining, Cleo received 6, and Pal received 5, respectively. Ayumu showed 40.6% of correct trials in the first session but 87.5% in the last session (mean response time for correct trials was 951 ms), Cleo showed 12.5% in the first session, and 81.3% in the last session (1334 ms), and Pal showed 43.8% in the first session and 88.5% (1096 ms) in the last session.
After the preliminary training, the chimpanzees were shifted to the main experiment. As in previous experiments, the target/distractor conditions varied across sessions, while the other conditions varied within sessions. Each session consisted of 48 trials, and the chimpanzees were given four sessions per target/distractor condition. As in the preliminary training, the correction method was applied to the main experiment, but data from correction trials were not used for the data analysis.
Results and Discussion
Figure 7 shows the mean response times for correct trials for each target/distractor condition. As can be seen from the figure, the F/A condition was more inefficient than the F/D condition. The results for the No Objects condition were almost identical to those for the F/D condition. Table 2 shows a summary of the selected model. The model selection process is shown in Table S1. The selected model included set size (β = 74.63, [60.66, 88.60], p < .001) and target/distractor condition as fixed effects. The detailed inspection of the parameter estimates for the target/distractor condition revealed differences between the F/D and F/A conditions (β = 116.70, [59.10, 174.30], p < .001) and between the No Objects and F/A conditions (β = 156.64, [114.41, 198.87], p < .001).
As predicted by results from the previous experiments, the search for the front face was hampered by the distractors in which a person held an object but did not look at it. These results suggest that the presence or absence of interaction between the person and object in the scene, rather than the attentional shift triggered by the other’s gaze, affected the visual search performances in chimpanzees. In other words, the chimpanzees might initially direct their attention to the agent–object interaction as a whole (cf., configural processing of social interaction in humans; Papeo & Abassi, 2019).
Experiment 6
In the previous experiments, it has been repeatedly shown under several conditions that the attentional state of others toward an object affected the search for the object and the gaze direction in chimpanzees. It might be concluded that, in chimpanzees, attention is more likely to be captured by the state in which the other individual holds an object but does not pay attention to it and that this attentional capture causes an interference effect on visual search performances. However, this conclusion is based on the results from the experiments using photographs of humans (not conspecifics) and artificial objects by three juvenile chimpanzees as young as 6 years old. One should be cautious about concluding based on a small sample, even if the same results are consistently shown. Furthermore, the present results are limited to juvenile chimpanzees and thus may be different for adult chimpanzees. It has been suggested that the mode of face recognition may differ between young and adult chimpanzees (Dahl et al., 2013). Gao and colleagues (2020) also indicated that chimpanzees’ perceptual processing of the body might differ between the bodies of chimpanzees and humans. Therefore, approximately nine years after Experiment 5, we conducted follow-up experiments with five adult chimpanzees (including those who had participated in the previous experiments) using chimpanzee photographs.
Methods
Participants
Of the three chimpanzees who had participated in the previous experiments, Cleo and Pal participated again in Experiment 6. Since the present experiment was conducted 9 years after Experiment 5, they were 15 years old (adult) at the onset of Experiment 6. In addition to these two chimpanzees, Ai (GAIN ID#0434, 40 years old at the beginning of the experiment), Chloe (GAIN ID#0441, 35 years old), and Pendesa (GAIN ID#0095, 39 years old) participated in the experiment. Ai was Ayumu’s mother, and Chloe was Cleo’s mother. The new individuals were members of the same group as Cleo and Pal and had participated in various kinds of computer-controlled perceptual and cognitive experiments (Matsuzawa et al., 2006; Tomonaga, 2001a; Tomonaga et al., 2003).
Ethics Statements
The care and use of the chimpanzees adhered to the 3rd edition of the Guide for the Care and Use of Laboratory Primates of the institute. The experimental designs of the present study with chimpanzees were approved by the Animal Welfare and Animal Care Committee of the institute (2016-064). All procedures also adhered to the Guideline of the Animal Experimentation of the Japanese Society of Animal Psychology, Guideline for the Care and Experimental Use of Captive Primates of the Primate Society of Japan, Code of Ethics and Conduct of the Japanese Psychological Association, and Japanese Act on Welfare and Management of Animals.
Stimuli
Photographs of a chimpanzee, not a human, were used for the present experiment. The original was a royalty-free photograph from Getty Images (Credit: Anup Shah, Creative#:200325811-001), and the stimuli were created by retouching this photograph on gray backgrounds using Photoshop® (see Figure 8). The objects in the photographs were the same as those in previous experiments. The size of the photograph was the same as the previous one. As in Experiment 3, we also prepared stimuli with the silhouetted chimpanzee. In addition, we prepared stimuli in which the silhouette area corresponding to the face was brightened schematically to examine the effect of brightness contrast, as discussed in Experiment 3.
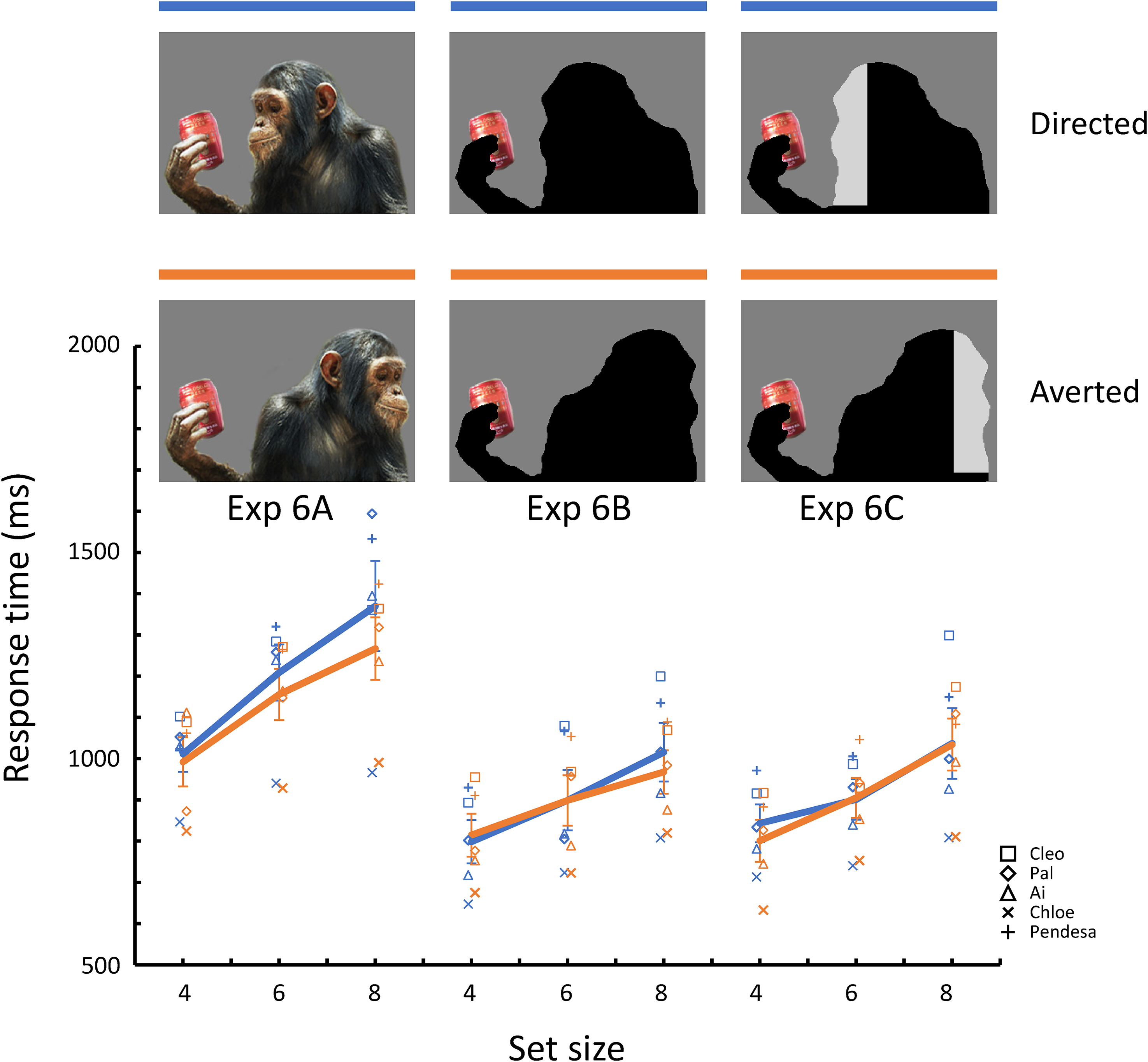
Mean response times for correct trials for each target/distractor condition in Experiment 6.
Procedure
Experiment 6 was conducted under the same settings as the previous experiments, and was separated into three sub-experiments using the three stimulus sets described above. Before the main experiment, all chimpanzees received preliminary training in which only objects were presented on a gray background. The mean percentage of correct trials in the last session of pretraining for the conjunction search was 96.9% (mean response time was 935 ms) averaged across participants
Following the preliminary training, the main experiments were conducted. All procedures were the same as in Experiment 2 except for the stimuli used in each experiment. Photographs of chimpanzees, silhouettes, and silhouettes with bright sides were used in Experiments 6A, 6B, and 6C, respectively. Each session consisted of 48 trials. The correction method was not applied in the present experiments. Four sessions were conducted for each target/distractor condition in each experiment. Data analysis was performed separately for each experiment.
Results and Discussion
Figure 8 shows the mean response times for correct trials per target/distractor condition for each sub-experiment. Table 2 presents a summary of the models selected. The model selection processes are shown in Table S1. In Experiment 6A, in which photographs of chimpanzees were used, the Averted condition was more efficient than the Directed condition. In contrast, there were no differences between the target/distractor conditions in Experiments 6B and 6C, in which the two types of silhouettes were presented, respectively. The model including set size (β = 70.24, [59.43, 81.06], p < .001) and target/distractor condition (β = −41.47, [−75.03, −7.91], p = .015) as fixed effects was selected for the data of Experiment 6A, whereas the models including set size alone were selected for those of Experiments 6B (β = 42.59, [34.48, 50.71], p < .001) and 6C (β = 47.38, [38.95, 55.81], p < .001).
Experiment 6A, where the adult chimpanzees participated and photographs of conspecifics were used, well replicated the results of the previous experiments. However, as in Experiment 3, when internal features such as eyes were removed, they showed no differences in search performances between the target/distractor conditions, confirming again that this search-asymmetry-like effect was caused by differences in the state of social attention of the other individuals. Furthermore, the effect of the other nonsocial cue, brightness contrast around the head part, was not observed. Taken together, these results suggest that the effect of Averted photographs on visual search performances is relatively robust in captive chimpanzees (at least from the same facility).
Experiment 7
In the final experiment, we tested human participants in the same settings as chimpanzees to confirm the validity of the current experimental settings and to examine species differences.
Methods
Fourteen human adults (undergraduate students of the university, nine females and five males) participated in the present experiment. All participants had a normal or corrected-to-normal vision. The experiment was conducted in the same booth as the chimpanzees. Each participant wore a laboratory coat and mask and entered the booth that had been thoroughly cleaned and sanitized. The stimuli were the same as those used in Experiment 1. As in Experiment 2, only the conjunction search type was given to the participants. The procedure was the same as for the chimpanzees, except that the intertrial interval was reduced to 1 s, and a food reward was not delivered for each correct choice. Each participant was verbally instructed to find the “red can” in the search display and was given several practice trials before the experimental session. Each session consisted of 96 trials, and each participant received one session for each target/distractor condition. For the data analysis, GLMM was used, but only participant was set for a random effect.
Ethics Statements
The experimental design was approved by the Human Research Ethics Committee of the institute (2016-05) and adhered to the Guideline of the Human Research Ethics of the institute and the Code of Ethics and Conduct of the Japanese Psychological Association. Informed consent was obtained from participants.
Results and Discussion
The mean percentage of error trials for human participants was 0.5% (SEM = 0.2%; Table 1). Figure 9 shows the mean response times for correct trials for each target/distractor condition. Table 2 shows a summary of the selected model. The model selection process is shown in Table S1. Contrary to the chimpanzees, humans showed more efficient search in the Directed condition than in the Averted condition. The selected model included set size (β = 58.60, [52.02, 65.17], p < .001) and target/distractor condition (β = 40.60, [21.46, 59.75], p < .001) as fixed effects.
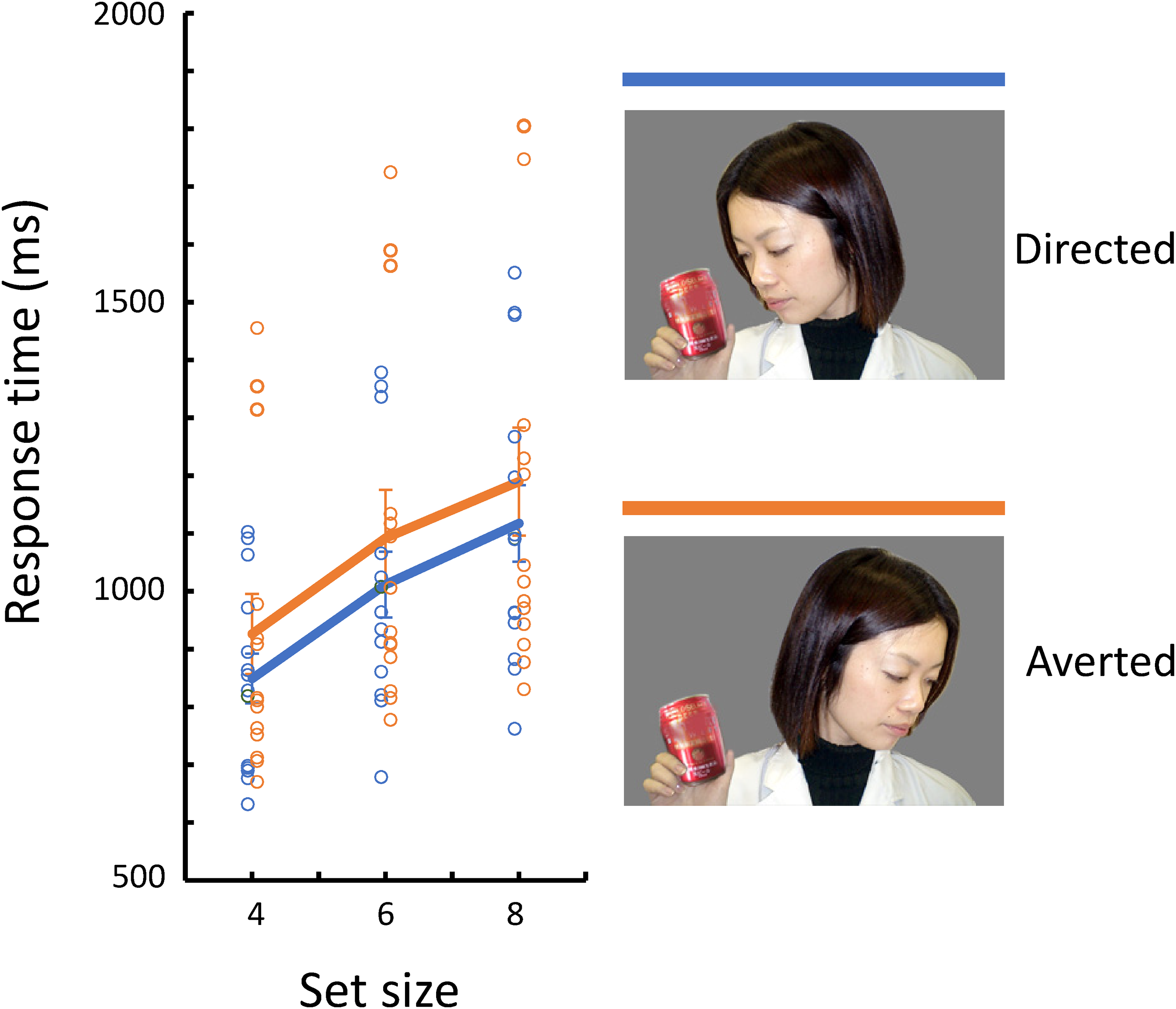
Mean response times for correct trials for each target/distractor condition in Experiment 7. Symbols indicate each participant’s data. Solid lines indicate averages across participants.
Visual search performances by the human participants were also affected by the attentional state of the person in the photograph. In this respect, the validity of the stimuli and experimental procedures used in the chimpanzees was verified. However, the effect was opposite to the chimpanzees. In humans, the object held in hand and attended to by the other person was detected more efficiently than the object held but not attended to. These results are consistent with the previous studies on joint attention, the shift of attention by gaze cue, and the recognition of goal-directed actions in humans. However, it remains why the results differed for chimpanzees and humans. This most critical issue is discussed further in the General Discussion.
General Discussion
In the present study, we examined the effects of the attentional state of others on visual search in chimpanzees and humans. We found that both chimpanzees and humans showed differences in the search for the object held and looked at by the person and for it held but not looked at. Interestingly, however, there were clear species differences. The chimpanzees searched for the object held but not looked at by the person more efficiently (Experiment 1). These results were not due to a simple violation of expectancy, such as “it is strange to hold an object but look away toward empty side” (Experiment 2). Furthermore, nonsocial cues such as the proximity relationship between the head and object or the brightness contrast between the front (face part) and back of the head also did not explain the results (Experiments 3, 4, 6B, and 6C). In addition, the objects held but not attended to caused cumulative interference effects when they were presented as the distractors (Experiment 4). Furthermore, even though the chimpanzees were required to search for the head direction instead of the object, the same effect was observed: “a person holding an object but not looking at it” caused a more substantial interference effect than the person holding an object and looking at it (Experiment 5). In Experiment 6A, these results were replicated with adult chimpanzees using photographs of chimpanzees. On the other hand, an experiment with humans in the same settings (Experiment 7) showed that, conversely, objects held and looked at by the person were more efficiently detected, suggesting species differences.
The results from humans in Experiment 7 are consistent with the first prediction described in the introduction. That is, they are consistent with previous studies of joint attention, the shift of attention by gaze, or the recognition of goal-directed actions (Frischen et al., 2007; Myowa-Yamakoshi et al., 2012; Tomasello, 1995). These studies indicated that the observer’s attention shifted to objects in the direction of the other’s attention, which might initiate triadic social interaction with the others (e.g., Tomasello, 1995) and enhance the perceptual sensitivity to the object (e.g., Seow & Fleming, 2019). The perceptual sensitivity is critical to the present study. Nonsocial attentional precues are known to increase perceptual sensitivity in the attended area and decrease the sensitivity in the unattended (Downing, 1988; Handy et al., 1996; Kravitz & Behrmann, 2008; Yeshurun & Carrasco, 1999). Studies of the attentional shift by the social cues such as gaze showed that these cues also trigger the attentional shift as nonsocial cues do (Driver et al., 1999; Frischen et al., 2007; Friesen & Kingstone, 1998; Langton & Bruce, 1999). In Experiment 7, the search may have been more efficient when the target was an object to which perceptual sensitivity was enhanced by the triadic social attention and more inefficient when such objects were presented as the distractors. In addition, the target object located in areas where social attention was not directed and thus the perceptual sensitivity was reduced would be less accurately identified and discriminated, contributing to the inefficient search (cf., Downing, 1988).
The question then arises as to how to interpret the results for chimpanzees. Of course, we cannot deny the possibility that the results of the present study were some kinds of false positives because of the small sample. However, given that the same results were repeatedly obtained in the chimpanzees in the same settings, they might not be simply dismissed as false positives. Chimpanzees may be more likely to pay attention to the state where “others are not paying attention.” Then why?
One of the possible accounts may be the violation of expectancy. From infancy, like humans, chimpanzees pay attention to what their mothers or other close individuals manipulate with their hands (Hirata & Celli, 2003; Kosugi et al., 2003; Ueno & Matsuzawa, 2005). This has also been confirmed in the eye-tracking study (Myowa-Yamakoshi et al., 2012). Through such experiences, they may form knowledge or belief of the relationship between agents and objects, that “agents hold objects in their hands to manipulate and pay attention to them.” And if the state in which “the agents are not paying attention to the object held by them” causes a violation of expectancy, then their attention would be captured by such scene as a result.
Human infants are sensitive to violations of expectancy formed by short-term experiences (such as the familiarization phase in experimental settings) as well as long-term experiences during everyday life. For example, Woodward (1998) showed that after infants repeatedly looked at a goal-directed action (grasping one of two objects), their looking time increased when they saw an action directed to a different goal (i.e., a different object). Furthermore, the predictive looking toward the goal before the action is known to occur both in human infants and in chimpanzees (Biro & Leslie, 2007; Cannon & Woodward, 2012; Kano & Call, 2014; Kano & Hirata, 2015). The prolonged looking time while observing the expectancy-violated scene might activate attentional processing of the other objects in that scene. In particular, chimpanzees have shorter fixation durations to social stimuli such as faces than humans, and fixations quickly move among objects in the scene one after another (Kano & Tomonaga, 2011). These species-specific eye-movement (attentional shift) patterns in chimpanzees might be led to the attentional capture of objects not attended by the agent in the scene more quickly than in humans.
However, Experiment 2 indicates that expectancy violation, as discussed here, may not be sufficient to explain the chimpanzees’ results. In Experiment 2, to eliminate the state of “holding but not looking,” the persons in the photographs held objects in both hands. That is, all of the persons in the search display directed their attention to some object. Even under these conditions, the chimpanzees efficiently searched for the target object held but not attended to by the person. Therefore, the violation of simple expectancy of “holding but not looking” alone cannot explain the present results.
Here we propose another hypothesis, although it is speculative in that it has not been empirically examined: attention is captured not only to objects but also to social interactions as a whole. Whether for object search or head direction, the chimpanzees’ attention was more readily captured by the state of “holding but not looking.” The attentional state of the person in the scene enhanced not only perceptual sensitivity to objects but also sensitivity to the person herself. If the person and object are integrated into “interaction” by social attention, the perceptual sensitivity to the interaction would be simultaneously enhanced as a whole. It is also well known that even if spatially separated, several objects are perceived as some kind of unit, called perceptual grouping. And then object-based attention (e.g., Egly et al., 1994), in which attention is directed to the whole of such perceptual groupings, may be activated (Scholl, 2001). It has recently been reported that both gestalt groupings and groupings based on social interactions (“social binding”) could occur in humans. For example, Papeo et al. (2017) reported that the inversion effect was stronger when two persons were facing each other (with interaction) than when they were not (without interaction) in a categorization task. These results suggest that dyads grouped by social binding may be processed configurally. If perceptual grouping based on the interaction described above also occurs in chimpanzees, object-based attention to the interaction would occur (cf., Ushitani et al., 2010; Yin et al., 2018). If “interaction-based” attention as proposed here had been activated in the Directed and Averted conditions of Experiment 1, a scene in which a person looked at an object held in hand was one unit for attention. In contrast, a scene in which she held but did not look at the object was divided into two units (the person and the object were separated). In such contexts, the number of units in each photograph may affect search performances. Reijnen et al. (2013) found a search asymmetry in humans that search for more dots in multiple arrays was more efficient than vice versa (cf., Tomonaga, 2001a). If the scene with more objects captures attention more readily, then object search in that scene would be facilitated. Specifically, in the present study, the scene in which an object is held but not attended to by the agent (two units for attention) may capture the chimpanzees’ attention more readily than the scene in which it is held and attended to by her (one unit).
Papeo (2020) further pointed out that the object-superiority effect (Homa et al., 1976) and the object-inferiority effect (Suzuki & Cavanagh, 1995) may occur at different processing levels in dyads that are perceptually grouped by social binding (Papeo & Abassi, 2019; Papeo et al., 2019; Vestner et al., 2019, 2021a, 2021b). Papeo et al. (2019) found search asymmetry between interacting and non-interacting dyads: efficient search for a facing (i.e., interacting) dyad among non-facing (non-interacting) dyads than vice versa, similar to the results of Experiment 7 with humans in the present study. On the other hand, when participants were instructed to search for the pose of a single person comprising a dyad, search performances were better for the pose of the person in the no-interactive dyad than in the interactive dyad, indicating the object-inferiority effect. Conversely, Vestner et al. (2019) found that a single body was individuated better in a short-term memory task when first seen in a facing dyad than in a non-facing dyad (i.e., object-superiority effect).
Although Papeo et al. (2019) studied the effect of the interaction-based grouping of two persons, Green and Hummel (2006) showed that a functionally interacting object pair, such as a pitcher and glass, was also perceptually grouped, and this grouping had a facilitating effect on the identification of objects in the pair. Given these findings, similar effects would have occurred in human–object pairs in goal-directed actions in the present study: the configural processing and efficient detection of the entire pair, as well as the resulting facilitation and interference effects on the identification of individual objects within the pair.
While not mentioned by Papeo and colleagues, triadic social attention within a socially-bound dyad may affect the perception of dyads. The social attention of others increases the perceptual sensitivity of the objects (Seow & Fleming, 2019), and object-based attention is activated within the socially-bound dyad by shaking hands (Yin et al., 2018). Such effects of the attentional shift within the unit could compete with the object-inferiority effect. But it may be evident that how these effects interact with each other depends on the task demand (Papeo, 2020; Papeo et al., 2019; Scholl, 2001). In the present study, the interactions (goal-directed actions) between the other persons and objects would have formed the socially-bound units. On the other hand, the scene containing more units would have been efficiently detected. Furthermore, the socially-bound unit would more likely capture the observer’s attention in that scene. If the other’s social attention enhances the perceptual sensitivity of the object in the socially-bound unit in humans, then the object held and looked at by the person would result in a more efficient search (or inefficient search if those scenes are the distractors). Conversely, if the object-inferiority effect through social binding and the rapid shifts of attention between objects in the scene (Kano & Tomonaga, 2011) influence the visual search performances in chimpanzees, the object held but not looked at by the person would be more efficiently detected resulting in facilitation effect as a target and cumulative interference effect as distractors.
However, this hypothesis is highly speculative as of yet. At the very least, empirical verification of several points would be necessary. The first is the demonstration that socially-bound interaction is processed as a whole. It is already reported that chimpanzees show inversion effects not only in the face but also in the body perception (Dahl et al., 2013; Gao et al., 2020; Gao et al., 2018, 2020a, 2020b; Parr et al., 1998; Tomonaga, 1999, 2007a; Tomonaga & Imura, 2015). We could explore the configural processing of social interactions by examining the inversion effect using stimuli such as the two agents used in Papeo et al. (2019) and the goal-directed actions with objects used in Myowa-Yamakoshi et al. (2012) and the present study. The second question is how perceptual sensitivity changes in the object or spatial area to which the other’s social attention is directed in both species. In addition, not only the comparison between species but also between social and nonsocial cues would be necessary. And third, the visual search for numerosity should be examined from the comparative perspective. Previous studies demonstrated search asymmetries in various stimuli in chimpanzees (Tomonaga, 1993, 2001a, 2001b; Tomonaga & Imura, 2010; Wilson & Tomonaga, 2022), including search asymmetries related to quantity (e.g., more is easier to find; Tomonaga, 2001a). Do chimpanzees show search asymmetry for numerosity as observed in humans?
Finally, and most importantly, it should be examined whether “interactions” as whole capture attention in chimpanzees. Object-based attention has been reported in chimpanzees (Ushitani et al., 2010). Therefore, it may be possible to investigate interaction-based attention using object-based precueing tasks used in humans (Egly et al., 1994; Yin et al., 2018) and chimpanzees. Note that it would be essential to separate the effect of the shift of the observer’s attention triggered by the agent’s attention from the effect of interaction-based attention (cf., Yin et al., 2018). Recently, it was reported that efficient search occurred not only for facing dyads but also with facing arrows, fans, and so on (Flavell et al., 2022; Vestner et al., 2021b), pointing out that this effect may not be unique to social interactions. Through these investigations, it is necessary to examine further the species differences observed in the present study.
In the present study, we focused on the relationship between agents and objects as the attentional state of others. However, we can also study attention to the social interaction between agents. Do chimpanzees discriminate the states of social attention (looking at each other, not looking at each other, etc.) between the two individuals? If so, which attentional state captures the observer’s attention more readily? At the onset of the present study, few studies on attention to social interactions had been conducted. Among them, Beier and Spelke (2012) reported that human infants at 10 months old who had habituated to mutual gaze dishabituated to averted gaze, indicating they are capable of discriminating attentional states between agents. And as noted above, in recent years, many studies have been conducted on how humans perceive and recognize social interactions in various ways (Papeo et al., 2019; Vestner et al., 2019; Yin et al., 2018). These studies have shown that social interactions cause perceptual grouping (social binding), capture the observer’s attention during visual search, are processed configurally similar to faces and bodies resulting in object-inferiority and object-superiority effects, and are activated by interaction-based attention like object-based attention. Do similar effects also observe in nonhuman animals (cf., Matsuno, 2021)? Based on the present results, attention to social dyads would cause different effects in humans and chimpanzees. In humans, interactions such as eye contact are easier to find as targets and cause interference effects as distractors. In chimpanzees, dyads looking away from each other may capture attention more readily (cf., Vestner et al., 2021b).
Although further studies are needed, this is the first study to systematically examine the effects of the attentional state of others in goal-directed actions on visual search performances in chimpanzees. The species differences between humans and chimpanzees found in the present study are quite suggestive for examining species differences at a more fundamental level, such as the perception of social attention and social interactions and operating characteristics of spatial attention. It is necessary to investigate further the perception and cognition of social interactions in chimpanzees from the perspective of comparative cognition.
In closing, we should discuss the limitations of the present study. Due to the limitations of research resources, we had to discuss the results from the chimpanzees with extensive experimental experiences. Inevitably, not only daily experience but also various experimental histories could influence the results in question. In addition, due to the small number of individuals, confounding factors that should normally be adjusted for could bias the results. In Experiment 6, however, we were able to show, albeit indirectly, that the present results were observed not only in the juvenile chimpanzees but also in adults. In the future, it will be necessary to further verify the generality and specificity of the results by increasing the number of individuals and species by using tasks that require less training, such as eye-tracking tasks (e.g., Hopper et al., 2021; Kano & Tomonaga, 2009) through the effective use of a multi-institutional collaborative replication study platform that has attracted much attention in recent years (ManyPrimates, 2019a, 2019b).
Supplemental Material
sj-pdf-1-pec-10.1177_03010066231158255 - Supplemental material for Attention to inattention: Effect of the other’s attentional state on visual search performance in chimpanzees
Supplemental material, sj-pdf-1-pec-10.1177_03010066231158255 for Attention to inattention: Effect of the other’s attentional state on visual search performance in chimpanzees by Masaki Tomonaga and Tomoko Imura in Perception
Supplemental Material
sj-xlsx-2-pec-10.1177_03010066231158255 - Supplemental material for Attention to inattention: Effect of the other’s attentional state on visual search performance in chimpanzees
Supplemental material, sj-xlsx-2-pec-10.1177_03010066231158255 for Attention to inattention: Effect of the other’s attentional state on visual search performance in chimpanzees by Masaki Tomonaga and Tomoko Imura in Perception
Footnotes
Acknowledgments
We are very grateful to Drs. T. Matsuzawa, M. Tanaka, M. Hayashi, I. Adachi, Y. Hattori, and the staff at the Language and Intelligence Section, and the Center for Human Evolution Modeling Research of the Primate Research Institute, Kyoto University for their technical advice, support, and care of the chimpanzees.
Author Contributions
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Japan Society for the Promotion of Science, (grant number 12002009, 13610086, 16002001, 16300084, 19300091, 20002001, 15H05709).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.