
Abstract
The three-factor model argues that the spatial orienting benefit triggered by the cue, the spatial selection benefit of cue–target matching, and the detection cost of distinguishing the cue from the target contribute to the measured inhibition of return (IOR) effect. According to the three-factor model, the spatial selection benefit dominates the occurrence of the IOR effect in the discrimination task, while the detection cost is negligible. The present study verified the three-factor model in the discrimination task under the cue–target paradigm by manipulating the spatial location and nonspatial feature consistency of the cue and the target as well as the promotion or hindrance of attentional disengagement from the cued location with a central reorienting cue. The results indicated that the three factors of the three-factor model contributed to the measured IOR effect in the discrimination task. Interestingly, the IOR effect was stable when the cue and target were perfectly repeated and attention was maintained at the cued location, implying that detection cost was not a negligible factor. The current study supported the contribution of all three factors in the three-factor model to the measured IOR effect; however, we argue that the role of detection cost in the discrimination task under different paradigms should be further refined.
In the original study of inhibition of return (IOR), Posner and Cohen (1984) used the cue–target paradigm (Posner, 1980), in which an uninformative peripheral stimulus (the “cue”) is presented to attract transient attention, followed by a target stimulus (the “target”) at the same or opposite location of the cue. The researchers found that when the time interval between presentation of the cue and the target (stimulus onset asynchrony, SOA) was longer than 300 ms, participants showed slower response times (RTs) in the condition in which the cue and the target appeared at the same location (cued location) than in the condition in which the cue and the target appeared at different locations (uncued location); in other words, an IOR effect was observed.
The contributing factors of the IOR effect have long attracted interest in the field (Martín-Arévalo et al., 2016). Some researchers believe that the IOR effect in the original cue–target paradigm used by Posner and Cohen (1984) was caused by the difficulty of attentional reorientation; that is, difficulty in disengaging attention from the cued location and returning attention to the cued location led to the IOR effect (Klein, 2000; Prime et al., 2006); this is called the attentional reorienting hypothesis. However, a number of methods have been used to elicit the IOR effect, with some methods differing from the original method of Posner and Cohen (1984) (Dukewich & Klein, 2015). Specifically, some studies have required participants to respond to the cue in addition to the target; this is known as the target‒target paradigm (Maylor & Hockey, 1985; Pratt & Castel, 2001). In terms of task context, in addition to manipulating peripheral cueing of the cue and the target, some studies have also manipulated the consistency of nonspatial features of the cue and the target, such as color, shape, and semantic aspects (Hu et al., 2011; Pratt & Castel, 2001; Taylor & Donnelly, 2002; Zu et al., 2022). Although studies using different methods still use the umbrella term “IOR” to describe the phenomenon of a slower response to the previously cued location, the IOR effect observed in different experimental contexts may be generated by different processes or by multiple independent processes (Dukewich & Klein, 2015).
In this context, Lupiáñez and colleagues proposed a three-factor model of IOR (Lupiáñez, 2010; Lupiáñez et al., 2007), arguing that three independent processes contribute to the measured IOR effect, including the detection cost, spatial selection benefit, and spatial orienting benefit. The detection cost refers to the cost of distinguishing between the cue and the target. Specifically, when the cue and the target appear successively in the same spatial location, the visual system has a strong tendency to integrate the two stimuli. However, since the task requires participants to react to the target rather than the cue, processing resources will be consumed to distinguish between the cue and the target when presented in the same location, resulting in costs (Martín-Arévalo et al., 2013, 2019). Thus, the more similar the cue and the target are, the larger the detection cost (Hu et al., 2011; Lupiáñez et al., 2013). The spatial selection benefit refers to the benefit generated by matching the episodic representation of the cue and the target, which is based on Kahneman's object file theory. Specifically, under this hypothesis, when a stimulus is presented, the features of the stimulus (such as color, orientation, and shape) are integrated into a single episodic representation with the spatial location of the stimulus (Kahneman et al., 1992); when a latter stimulus is presented, the participants will automatically match the current stimulus with the previous stimulus if the two stimuli are presented at the same location. If the two stimuli presented successively are completely identical, the spatial selection benefit promotes the responses of the participants to the current stimulus. However, if the stimuli are not completely identical (such as appearing in the same location but with different features), a partial repetition cost will be generated due to this conflict. In this case, the responses of the participants will be slower than in conditions where the cue and the target are either completely consistent or completely inconsistent (Hommel, 2004, 2019). The spatial orienting benefit refers to the benefit of attentional orienting triggered by a peripheral cue to the cued location (Lupiáñez, 2010). This benefit is short-lived and will decay with the extension of SOA between the cue and the target. The spatial orienting benefit differs from the attentional reorienting hypothesis in that it does not suggest that attention is inhibited to return to the cued location after decaying from the cued location. The net facilitation or inhibition effect induced by the cue is the result of the sum of these three factors (Lupiáñez, 2010).
The three-factor model provides a clear expectation of the contribution of each of the three factors to the IOR effect under different experimental tasks (Lupiáñez, 2010), which is conducive to validating the model. According to the three-factor model, in a detection task, the detection cost is more pronounced (Lupiáñez, 2010; Schönhammer & Kerzel, 2017) because, in this task, the participants are required to detect whether the target is present; thus, the cost of distinguishing between cues and targets successively presented at the same location becomes the main contributing factor to the spatial IOR effect. The contribution of spatial selection benefit is smaller (Huffman et al., 2018; Tanaka & Shimojo, 2000) in the detection task because an establishment of the episodic representation of the target can enable participants to complete the task (Lupiáñez et al., 2001; Lupiáñez & Milliken, 1999); that is, participants are not required to represent the features of the target in fine detail and match the target features with the cue. The prediction of the three-factor model for the detection task has received empirical support. Specifically, in the cue–target paradigm, the IOR effect was larger when the nonspatial features of the cue and the target were consistent (Hu et al., 2011) because the similarity of the cue and the target increased the detection cost and masked the possible spatial selection benefit. In the target‒target paradigm, researchers found that there was no interaction between peripheral cueing and nonspatial feature consistency, with the IOR effect appearing regardless of whether the nonspatial features of the cue and the target were consistent (Fox & de Fockert, 2001; Tanaka & Shimojo, 1996, 2000). These results suggest that the detection cost plays a dominant role in the detection task and that the contribution of the spatial selection benefit to the IOR effect is decreased.
In discrimination tasks, the spatial selection benefit is thought to play a dominant role (Lupiáñez, 2010; Lupiáñez et al., 2013) because task-related features have a larger weight in the episodic representation of the stimulus, and whether the cue and the target share these features has a substantial impact on perceptual processing and response selection (Petruo et al., 2016; Takacs et al., 2020). The detection cost is negligible compared with the benefits and costs of matching the cue and target (Lupiáñez et al., 2013). The validation of the three-factor model in the discrimination task has mainly involved the target‒target paradigm. Such a task requires participants to respond to the cue immediately; researchers found that the spatial selection benefit determined whether the IOR effect occurred (Pratt & Castel, 2001; Taylor & Donnelly, 2002; Zu et al., 2022). That is, the spatial IOR effect occurred only when the nonspatial features differed between the cue and the target, while a target identical to the cue led to a spatial facilitation effect due to the spatial selection benefit. This is in line with the expectation of the three-factor model for the discrimination task in the target–target paradigm (Lupiáñez et al., 2013).
However, given the differences in experimental paradigms, the respective contributions of the three factors of the three-factor model in the discrimination task under the cue–target paradigm need to be further evaluated; this examination is important for refining the three-factor model. In the target‒target paradigm, an immediate response to the cue may weaken the detection cost. On the one hand, a study has shown that an immediate response to the cue reduced the IOR effect induced by the cue compared to no immediate response (Hommel & Colzato, 2004), possibly because responding to the cue reduced the costs of distinguishing between the cue and the target. On the other hand, when an immediate response to the cue was required, participants bound the response to the cue (Hommel, 2019), thus promoting the response to a repeated stimulus at the same location. This phenomenon is similar to using a stimulus-response repetition shortcut (Hilchey et al., 2020); that is, when the same stimulus reappears, the response of the participants to the previous stimulus is directly triggered to improve the response ability and shorten the current response time (Pashler & Baylis, 1991). Thus, in the cue–target paradigm, the respective contributions of the three factors may change when the participants do not need to respond to the cue immediately.
The aim of the current study was to validate the three-factor model using a discrimination task with the cue–target paradigm and to evaluate the relative contribution of the detection cost, spatial selection benefit, and spatial orienting benefit to the measured IOR effect. Based on previous studies (Chi et al., 2014; Martín-Arévalo et al., 2013; Wang et al., 2020), the present study orthogonally manipulated peripheral cueing and nonspatial feature consistency between the cue and the target. In addition, to isolate the effect of attentional orienting, in contrast to previous studies (Pratt & Castel, 2001; Taylor & Donnelly, 2002; Zu et al., 2022), the present study promoted or hindered attentional disengagement from the cued location with a central reorienting cue. In this context, a detection cost was incurred when the target appeared in the cued location and was maximized when the nonspatial features were consistent between the cue and the target. A spatial selection benefit occurred when the locations and nonspatial features were completely consistent between the cue and the target, while a partial repetition cost occurred when the cue and the target were partially consistent. A spatial orienting benefit was generated when the central reorienting cue pointed to the cued location (i.e., hindering attentional disengagement from the cued location) but disappeared when the central reorienting cue pointed to the uncued location (i.e., promoting attentional disengagement from the cued location). The relative contribution of the three factors in the cue–target paradigm was evaluated by observing the facilitation or inhibition of the cued location under different experimental conditions. We expected that if the detection cost remained negligible in the discrimination task under the cue–target paradigm, the spatial facilitation effect would occur when the cue and the target were completely consistent and attention was maintained at the cued location.
Methods
Subjects
Twenty-eight participants (5 men, 23 women; mean age = 20.21 years) were recruited to participate in exchange for monetary compensation. One participant was left-handed, while the rest were right-handed. None of the participants had mental illness, color blindness, or color weakness, and they had normal or corrected-to-normal vision. This study was approved by the Academic Committee of the Department of Psychology, Soochow. The study conformed to the principles outlined in the Declaration of Helsinki, and informed consent was obtained from the participants.
A power analysis was performed in G*Power 3.1.9.2. (Faul et al., 2009), and the results were analyzed by repeated-measures ANOVA. The sample size was estimated on the basis of our previous study, which used a within-subject design and procedure similar to that of the present study, who reported ηp2 of 0.60 and 0.50 for the interaction between peripheral cue validity and nonspatial feature consistency in Experiments 1 and 2, respectively (Zu et al., 2022). Thus, the projected ηp2 of the expected interaction was set at 0.50 in the present study. Given a power (1–β err prob) of 0.80 and an alpha level of 0.05, the power analysis ultimately yielded a recommended sample size of 10. Thus, the sample size of the present experiment was sufficient.
Apparatus and Stimulus
The participants were in a dimly lit, soundproof chamber environment to minimize ambient noise and external distraction. The experiment was conducted on a Dell 3020 MT computer. The participants were seated 60 cm from a 23-inch Dell E2316Hf PC monitor, operating at a resolution of 1,280 × 1,024 pixels and a 60 Hz refresh rate. Stimulus presentation was controlled using the Psychophysics Toolbox (Brainard, 1997; Pelli, 1997) in MATLAB. All stimulus materials were drawn using black lines (RGB: 0, 0, 0) on a gray background (1 cd/m2). In each trial, three square boxes (2.5° × 2.5°) were arranged horizontally on the screen, with one square in the center of the screen and the other two on the left and right sides. The center-to-center distance between two adjacent squares was 5°. The central fixation point (1° × 1°), cue, target, and neutral stimuli were displayed inside the squares. Following the stimuli commonly used in previous nonspatial priming paradigms (Chi et al., 2014; Snodgrass & Vanderwart, 1980; Wang et al., 2020), the cue and the target stimuli were drawn outlines of a cat or a dog (2 ° × 2°), and the central reorienting cue was an arrow (1° × 1°) pointing left or right.
Procedure and Design
The experimental procedure is shown in Figure 1. Each trial began with the presentation of a central fixation cross (“+”). After a duration of 750−1,250 ms, the cue stimulus (a picture of a dog or a cat), was randomly placed in a square to the left or right of the fixation point. The cue stimulus was presented for 300 ms, and its features and presentation location did not predict the target stimulus. After the cue stimulus, an interstimulus interval (ISI) occurred, during which only the fixation cross and squares were displayed on the screen. After the 300 ms ISI, a central reorienting cue was presented for 300 ms in the central square. The central reorienting cue can hinder or promote attentional disengagement from the cued location (Martín-Arévalo et al., 2013). Another ISI occurred for 400 ms, followed by the presentation of a visual target for 300 ms. The target stimulus was the drawing of a dog or a cat, randomly placed in a square to the left or right of the fixation point. After the target stimulus was presented, participants were asked to perform a discrimination task regarding the identity of the target stimulus by pressing one of two buttons (“F” or “J” key) as fast and accurately as possible, with one button assigned to the cat and the other assigned to the dog. The response buttons were counterbalanced across participants, and the reaction times of the participants were limited to 1,000 ms. To promote the processing of the cue features, participants were asked to report the identity of the cue after identifying the target. Specifically, after providing their response to the target, a report screen was presented for 2,000 ms. Participants were required to respond by pressing the left or right key (i.e., “F” or “J” key) for the image shown on the corresponding side of the screen to indicate which of the pictures was the previously presented cue stimulus. On the report screen, the spatial locations of dog and cat images were randomized between trials, preventing participants from forming action plans after the cue stimulus was presented. This experimental setup ensured the processing and maintenance of cues. The intertrial interval (blank screen) was a random duration ranging from 1,300 to 2,100 ms (with 200 ms increments: 1,300, 1,500, 1,700, 1,900, or 2,100 ms).
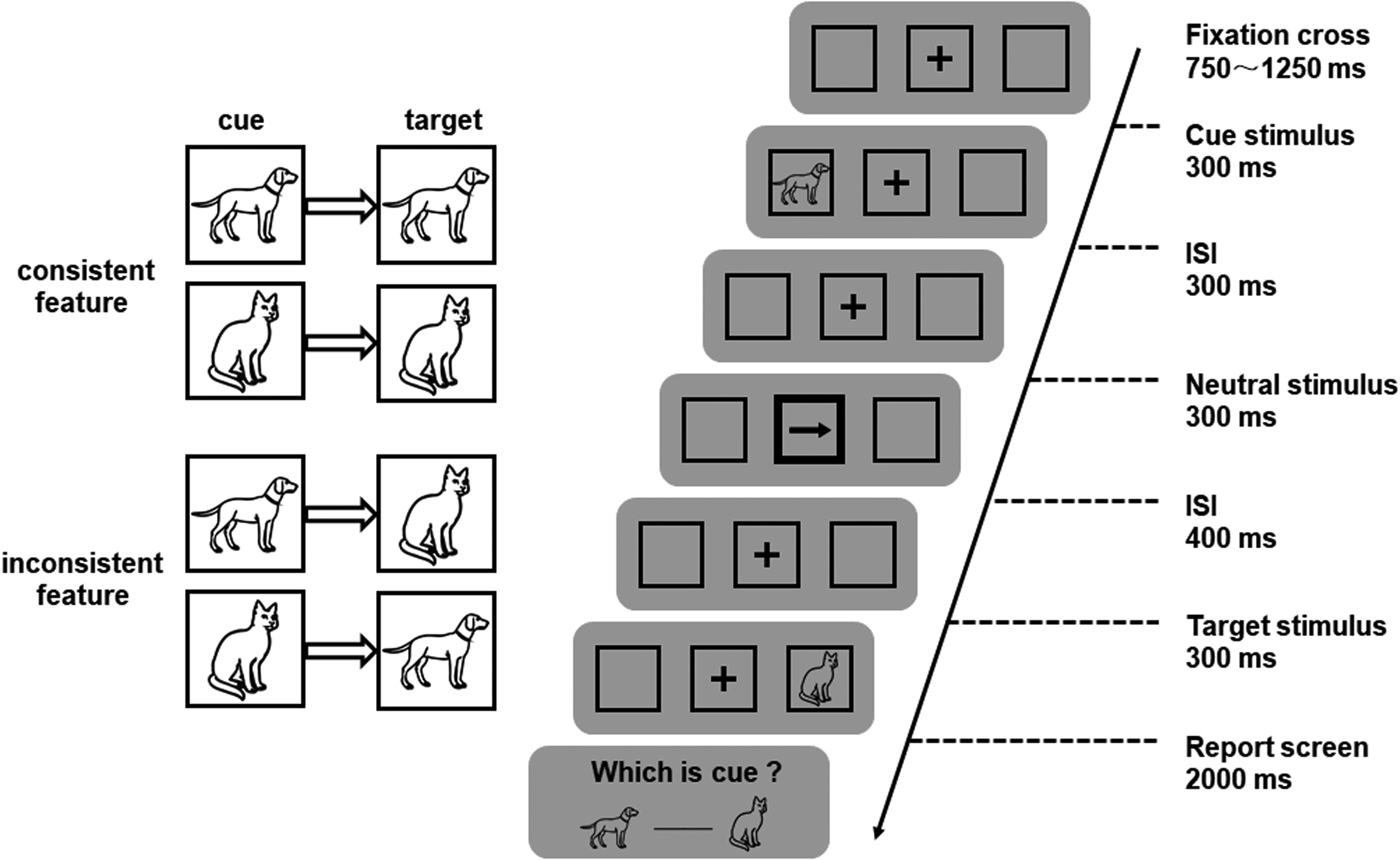
Experimental procedure. The cue and the target stimuli were pictures of dogs or cats and were randomly placed in a square to the left or right of the fixation point. The central reorienting cue was an arrow in the central square that pointed to the left or right. The participants were asked to perform a discrimination task regarding the identity of the target and to identify the previously presented cue stimulus after providing their response regarding the target stimulus.
Therefore, the present study had a 2 (peripheral cue validity: valid vs. invalid) × 2 (nonspatial feature consistency: consistent vs. inconsistent) × 2 (central cue orienting congruency: congruent vs. incongruent) within-subjects experimental design. In the consistent feature condition, the cue and the target were identical, and in the inconsistent feature condition, the cue and the target differed. In the condition of congruent orienting of the central cue, the arrow pointed to the same location as the cued location, and in the condition of incongruent orienting of the central cue, the arrow pointed to the opposite location as the cued location. The formal experiment consisted of eight blocks with 64 trials in each block and 512 trials in total. The participants completed 40 practice trials before the formal experiment. The entire experiment took a total of 70 min.
Results
To ensure participant processing of the cues, only the trials on which participants provided correct responses to the cues were included in further data analyses. We also excluded trials with no responses, incorrect responses, or extreme responses (trials with mean RTs exceeding three SDs from the mean across all experimental conditions for each participant) in the target discrimination task from the subsequent RT analysis. These criteria resulted in the exclusion of 7.9% of the trials.
With regard to the accuracy (ACC), a 2 (peripheral cue validity: valid vs. invalid) × 2 (nonspatial feature consistency: consistent vs. inconsistent) × 2 (central cue orienting congruency: congruent vs. incongruent) repeated-measures ANOVA was conducted. The results showed no main or interaction effects, Fs < 3.65, ps > .067.
We focused on the RTs (see Table 1 and Figure 2). The three-factor repeated-measures ANOVA showed a main effect of peripheral cue validity, F(1, 27) = 58.54, p < .001, η p 2 = 0.68, with RTs in the invalid cue condition (567 ms) faster than those in the valid cue condition (601 ms), suggesting that a spatial IOR effect occurred. The main effect of nonspatial feature consistency was significant, F(1,27) = 9.20, p = .005, η p 2 = 0.25, with RTs in the consistent feature condition (576 ms) faster than those in the inconsistent feature condition (591 ms), suggesting that feature-based facilitation occurred. The main effect of central-cue orienting congruency was not significant, F(1,27) = 4.04, p = .055. The interaction between peripheral cue validity and nonspatial feature consistency was significant, F(1, 27) = 48.97, p < .001, η p 2 = 0.65. Further simple effect analysis showed that when features were repeated between the cue and the target (i.e., in the consistent feature condition), the RTs in the invalid cue condition (566 ms) were significantly faster than the RTs in the valid cue condition (587 ms), F(1,27) = 22.50, p < .001, η p 2 = 0.46, and a spatial IOR effect occurred. When features were not repeated between the cue and the target (i.e., in the inconsistent feature condition), the RTs in the invalid cue condition (567 ms) were significantly faster than the RTs in the valid cue condition (615 ms), F(1,27) = 81.46, p < .001, η p 2 = 0.75, and a spatial IOR effect also occurred. The moderating effect of nonspatial feature consistency on the measured IOR effect was reflected in the size of the effect; the magnitude of the IOR effect (21 ms) under the consistent feature condition was significantly smaller than that of the IOR effect (48 ms) under the inconsistent feature condition, t(27) = 7.00, p < .001, Cohen's d = 1.32, 95% CI = [18.87, 34.52].
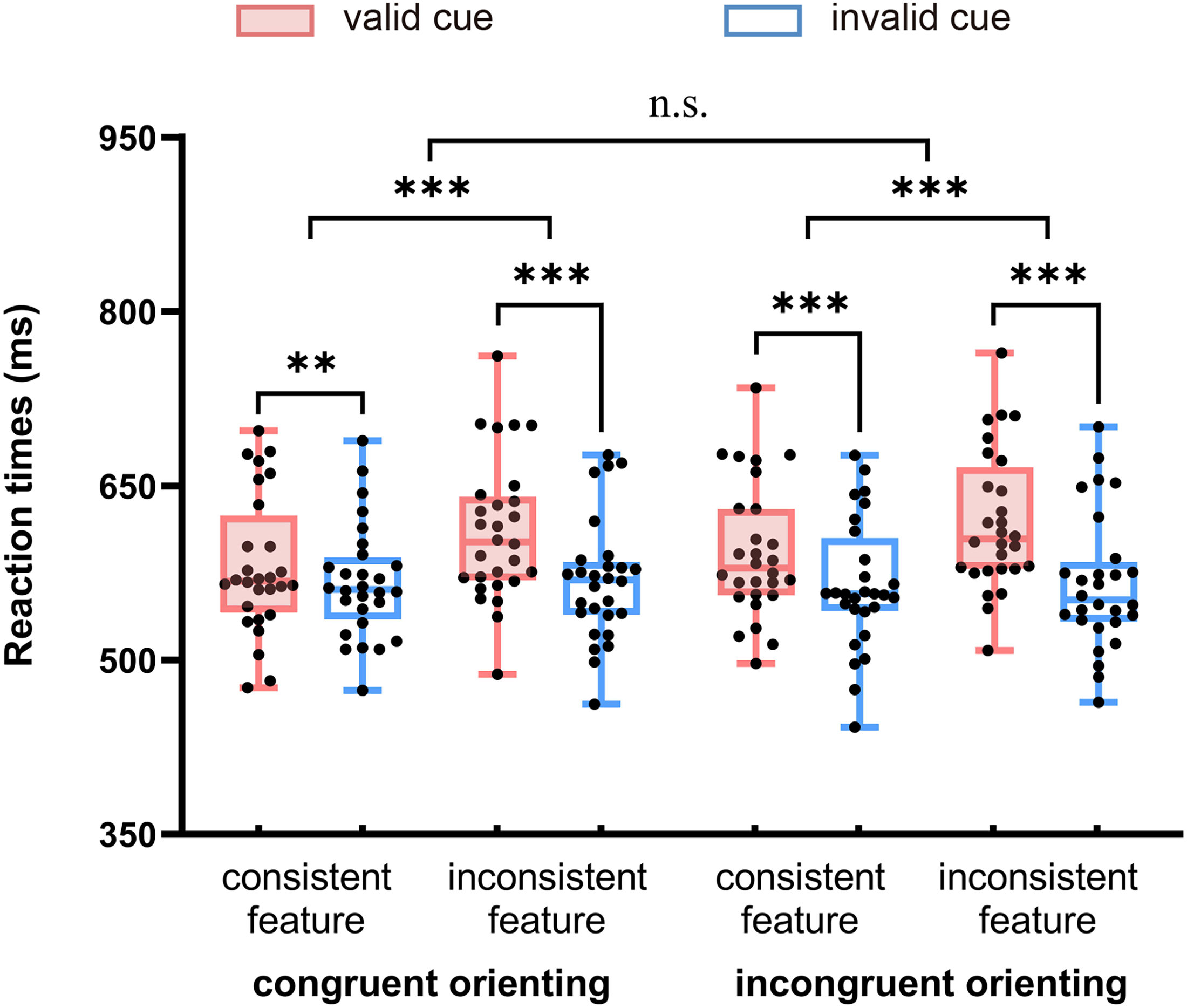
Mean response times (RTs) (ms) in all experimental conditions. In the boxplot, the central line is the median, the edges of the box indicate the 25th and 75th percentiles, and the whiskers extend to the most extreme data points not considered outliers. The points indicate individuals’ mean RTs within each condition. “n.s.” represents no statistically significant differences, and asterisks represent statistically significant differences (**p < .01, ***p < .001).
Mean response times (RTs) (ms) and standard errors of the mean under different experimental conditions.

In terms of the effect of attentional orienting, the interaction between peripheral cue validity and central-cue orienting congruency was significant, F(1, 27) = 7.12, p = .013, η p 2 = 0.21. Further simple effect analysis showed that when the central reorienting cue pointed to the peripheral cued location, the RTs in the invalid cue condition (568 ms) were significantly faster than the RTs in the valid cue condition (596 ms), F(1,27) = 43.06, p < .001, η p 2 = 0.62, and a spatial IOR effect occurred. When the central reorienting cue pointed to the peripheral uncued location, the RTs in the invalid cue condition (565 ms) were significantly faster than the RTs in the valid cue condition (612 ms), F(1, 27) = 62.11, p < .001, η p 2 = 0.70, and a spatial IOR effect also occurred. The moderating effect of attentional orienting on the measured IOR effect was reflected in the magnitude of the IOR effect: the magnitude of the IOR effect (28 ms) under the congruent orienting condition was significantly smaller than that of the IOR effect (41 ms) under the incongruent orienting condition, t(27) = 2.67, p = .013, Cohen's d = 0.50, 95% CI = [3.02, 23.13]. The three-way interaction was not significant, F < 1, and there was no evidence that central cue orienting congruency influenced the interaction between peripheral cue validity and nonspatial feature consistency.
A paired-sample t-test was performed for the peripheral cue-valid condition and peripheral cue-invalid condition under different nonspatial feature consistency and attentional orienting congruency conditions. The results showed that the RTs in the peripheral cue-valid condition were slower than those in the peripheral cue-invalid condition in all experimental conditions, ts > 2.91, ps < .007, and the IOR effect was stable across conditions.
Discussion
The aim of the current study was to validate the three-factor model using a discrimination task under the cue–target paradigm and to assess the relative contribution of the detection cost, spatial selection benefit, and spatial orienting benefit to the measured IOR effect. To this end, we manipulated peripheral cue validity and nonspatial feature consistency between the cue and the target to control the spatial selection benefit and manipulated the central-cue orienting congruency to control the spatial orienting benefit. On the one hand, the results indicated that the spatial selection benefit, spatial orienting benefit, and detection cost contributed to the measured IOR effect, further supporting the three-factor model (Lupiáñez, 2010). On the other hand, the IOR effect appeared stable when the cue and the target were identical (i.e., the consistent nonspatial feature condition) and attention was maintained at the cued location (i.e., the peripheral cue-valid condition), implying that the detection cost is not a negligible factor in a discrimination task under the cue–target paradigm.
The attentional orienting process triggered by the cue is widely considered the cause of the IOR effect (Pierce et al., 2018; Prime et al., 2003, 2006). As reflected in the term “IOR” (Posner & Cohen, 1984), researchers believe that the difficulty of returning attention to the cued location after disengaging attention from the cued location produced the IOR effect (Taylor & Klein, 1998). In contrast to the attentional reorienting hypothesis, the three-factor model takes into account not only the effect of attentional orienting triggered by the cue (the spatial orienting benefit) but also the effect of the cue presentation itself on the processing of subsequent stimuli at the cued location (the detection cost and spatial selection benefit) (Lupiáñez, 2010). In terms of the spatial orienting benefit, the current study used the central reorienting cue to hinder or promote attentional disengagement from the cued location. When the central reorienting cue hindered the disengagement of attention from the cued location, the IOR effect was smaller but still present; this was still the case when the cue and the target were completely identical, meaning that the process of attentional orienting was not necessary for the occurrence of the IOR effect in the current study and was only a contributing factor for this effect. This result is in line with previous studies, which have shown that the IOR effect occurred either when the peripheral cue at least partially predicted the target location (Chica et al., 2006), allowing attention to be endogenously maintained at the cued location, or when attention was automatically directed to the cued location via the central reorienting cue (Martín-Arévalo et al., 2013). Of course, in some contexts, the process of attentional orienting may still be the main factor in the occurrence of the IOR effect, such as in the detection task under cross-modal conditions (i.e., the cue and target stimuli are presented in different sensory modalities), in which the detection cost is reduced because the cue and the target belong to different modalities, and the spatial selection benefit is weakened because of the detection task; thus, the role of attentional orienting is prominent. This situation is essentially consistent with the three-factor model.
In terms of the spatial selection benefit, the current results showed that when the features of the cue and the target were consistent, the measured IOR effect was smaller in magnitude than that when the features of the cue and the target were inconsistent. These results correspond to the spatial selection benefit in the three-factor model (Lupiáñez, 2010). That is, when the target appears at the cued location, it is automatically matched with the episodic representation of the cue (Hommel, 2004; Kahneman et al., 1992). When the features of the cue and the target are consistent, the spatial selection benefit is generated, which reduces the measured IOR effect. When the features of the cue and the target are inconsistent, a partial repetition cost is generated, which increases the measured IOR effect. In fact, the similarity of two consecutively presented stimuli (in terms of location or task-related features), which may facilitate or impair behavioral performance, has been widely studied in the field of feature integration (Hommel, 2004; Takacs et al., 2020), as it reflects rapid learning of humans in response to previously presented stimuli or events (Henson et al., 2014). In the present study, the three-way interaction was not significant, and there was no evidence that attentional orienting moderated the interaction between peripheral cue validity and nonspatial feature consistency, consistent with previous research findings that feature integration was largely independent of attention (Hommel, 2005).
Interestingly, in the present study, the IOR effect was still present when the cue and the target were identical and attention was maintained at the cued location. The results imply that the detection cost imposed by the difficulty in detecting the target due to the integration of the target and the episodic representation of the cue played an important role in the present study; this cost counterbalanced the spatial orienting benefit and spatial selection benefit to yield the IOR effect. This observation is counterintuitive. Researchers believe that the detection cost is prominent in the detection task and negligible in the discrimination task compared to the spatial selection benefit generated by cue–target matching (Lupiáñez et al., 2013). The detection cost represents a mechanism that prevents the current task from being constantly interrupted by a novel stimulus; after one stimulus is presented, a subsequent stimulus presented in the same location is less able to capture attention and is more difficult to perceive as a new stimulus (Lupiáñez, 2010). The detection task emphasizes the detection of new stimuli (Lupiáñez & Milliken, 1999); thus, the detection cost plays a more important role. However, in the discrimination task, detection of the new stimulus is not enough to complete the task, and participants need to further identify the target. In this process, the completion of the task is affected to a greater extent by the similarity of the target and the episodic representation of the cue; thus, the spatial selection benefit plays a prominent role (Huffman et al., 2020; Lupiáñez et al., 2013). In fact, previous studies using the target‒target paradigm have found that in the discrimination task, the spatial IOR effect disappeared, and the spatial facilitation effect occurred if the target presented at the cued location was identical to the cue (Pratt & Castel, 2001; Taylor & Donnelly, 2002; Zu et al., 2022). Therefore, the findings from the present study differ from those of previous studies, possibly due to paradigm differences.
The prominent detection cost in the present study may have arisen because the current task did not require an immediate response to the cue. First, the detection cost originates from difficulty in detecting the target stimulus; that is, the attentional system has difficulty distinguishing between the cue and the target (Lupiáñez et al., 2013). However, if the response is completed for the cue, the cue may be “marked” by the response or the integration of the cue and the target may be interrupted, making the cue and the target easier to distinguish and reducing the detection cost. Second, in reference to feature integration studies (Hommel, 1998; Hommel & Colzato, 2004), responses to the cue may have bound the corresponding response, and the presentation of a target identical to the cue may have directly triggered the response (Henson et al., 2014), thus generating additional benefits and offsetting the detection cost. For the above reasons, it is possible that the contribution of the detection cost in the target‒target paradigm is obscured and emerges in the cue–target paradigm, in which an immediate response to the cue is not needed. Another important factor contributing to the current findings may be that participants were instructed to report the cue after responding to the target. This requirement allows the cue identity to be maintained in working memory (WM) (Baddeley, 2000). When responding to the target, this continuous activation of the cue representation may increase refractoriness to the same stimulus compared with contexts in which the cue is not maintained in WM. In addition, there was only one picture in each category selected in the current study, which may have caused the participants’ expectation for the presentation of the target picture. The presentation with only two possibilities was difficult to capture the participants’ attention. The similarity in basic features of cat and dog pictures may also increase the detection cost, consistent with the content of habituation (Dukewich, 2009). And it is possible that greater benefit will occur when the number of pictures is increased.
The present results imply that the contribution of the detection cost in the discrimination task under the cue–target paradigm should be further evaluated; that is, the detection cost is not a negligible factor in such experimental situations. In fact, researchers have attempted to combine the detection cost with the concept of habituation (Martín-Arévalo et al., 2013), which refers to the insensitivity of the visual system to stimuli presented at a repeated location (Klein et al., 2015). The process of habituation is related to the refractoriness of neural activity (DuKewich, 2009), which reduces incoming activity in response to repeatedly presented stimuli (Desimone, 1996). The researchers found that the IOR effect increased with the repetitions of cue (Dukewich & Boehnke, 2008), which was considered evidence that habituation plays a role in the IOR effect; that is, with increased repetitions of stimulus, the visual system became increasingly insensitive (DuKewich, 2009). Habituation lasts longer than the original content of the detection cost (Turatto & Pascucci, 2016), and there is no evidence that habituation is attenuated by performing the discrimination task. In the current study, the complete repetition of the cue and the target (i.e., using an identical cue and target) may have caused the attenuation of the perceptual salience of the target due to habituation and impaired target processing. Notably, the maintenance of cue representation in WM may be an important explanation for the detection cost highlighted in the current study; thus, the representation of cue in WM may be a boundary condition for the magnitude of the detection cost. Further studies should compare the IOR effect in conditions that vary in the maintenance of cue representation in WM using different tasks to expand the three-factor model.
Our experimental settings are similar to that of Fuentes et al. (1999), in which researchers manipulated both the location consistency and semantic relevance of prime and target, and found that semantic-based inhibition occurred in the uncued location when neutral stimulus between cue and target was presented, and when neutral stimulus was not presented, there was a semantic-based facilitation. And no facilitation or inhibition effect was observed in the cued location. In this study, although our study focused on the occurrence of spatial IOR, the analysis of another dimension of our results revealed that feature-based facilitation occurred when the spatial location of cue and target was consistent, t(27) = 5.36, p < .001, Cohen's d = 1.01, 95% CI = [17.45, 39.14], while no facilitation or inhibition effect occurred when the location of cue and target was inconsistent, t < 1. The similarities in paradigms but some differences in results make it necessary to discuss these two studies. We believe that the lack of semantic-based facilitation in the cued location in the study of Fuentes et al. (1999) may be due to the fact that although the prime (cue) and the target were semantically related, the stimuli themselves had different surface features (because the researchers used different words), so the same location might have incurred a partial repetition cost that offset the semantic-based facilitation (Hommel, 2004). As discussed by Fuentes et al. (1999), for the uncued location, when the neutral cue was not presented, the object file established by the cue might not have been closed, resulting in a facilitation effect. However, when a neutral cue was presented, the object file was closed, resulting in a partial repetition cost of the same feature at different locations when the target retrieved the cue, causing a semantic-based inhibition. In our study, since both the cue and the target were picture stimuli, the exact same feature could easily generate a benefit when the location between the cue and the target was the same. However, when the locations between the cue and the target were different but the feature was the same, on the one hand, partial repetition cost would be incurred, leading to feature-based inhibition. On the other hand, there were only two picture materials (a picture of a cat and a picture of a dog) selected in the current study, and the basic features of cats and dogs are highly similar, which might make the different pictures also produce partial repetition cost in a sense when the spatial locations and features of cue and target were different. Therefore, in the uncued location, partial repetition cost may be incurred between the cue and target for both the same and different features, resulting in no significant difference in behavioral performance between the same and different features, which needs to be further explored.
Given that many studies (Taylor & Klein, 1998), including the current study, have employed variations of the cueing paradigm, it is possible that the inhibitory effect observed in many cases may differ from those observed in the original studies of IOR (Posner & Cohen, 1984). Since the term “IOR” implies not only an effect but also a cause, we endorse the term “IOR” in some variant studies as an “inhibitory cueing effect,” particularly in variants that introduce additional effects on the outcome.
In conclusion, the present study validated the three-factor model in the discrimination task under the cue–target paradigm; that is, the cue-triggered spatial orienting benefit, detection cost, and spatial selection benefit generated by cue presentation contributed to the measured IOR effect. Moreover, the current study indicated that the role of the detection cost in the discrimination task should be further explored; that is, compared with the target‒target paradigm, the cue–target paradigm has a more prominent contribution to the detection cost, especially in cases where the representation of the cue is maintained.
Supplemental Material
sj-docx-1-pec-10.1177_03010066231190216 - Supplemental material for Detection cost: A nonnegligible factor contributing to inhibition of return in the discrimination task under the cue–target paradigm
Supplemental material, sj-docx-1-pec-10.1177_03010066231190216 for Detection cost: A nonnegligible factor contributing to inhibition of return in the discrimination task under the cue–target paradigm by Ming Zhang, Guangyao Zu and Aijun Wang in Perception
Footnotes
Author Contribution(s)
Author’s Note
Ming Zhang and Guangyao Zu contributed equally to this study and should be considered co-first authors.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article: This research was funded by the Suzhou Science and Technology Development Plan [People's Livelihood Science and Technology: SKY2022113], the 14th five-year plan of Jiangsu Province Education Science (B/2021/01/87), the Humanities and Social Sciences Research Project of Soochow University (22XM0017) and Interdisciplinary Research Team of Humanities and Social Sciences of Soochow University (2022). Ming Zhang was supported by the National Natural Science Foundation of China (31871092) and the Japan Society for the Promotion of Science KAKENHI (20K04381).
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.