
Abstract
Theories of working memory (WM) often distinguish between a central component and peripheral components for verbal and visual information. In the present study, we tested whether musicians differed from non-musicians on WM capacity and structure, with a particular focus on motor memory. We compared individuals with instrumental music training (n = 91) to those without musical training (n = 99) on seven WM tasks, measuring visual, verbal, and motor memory. The results showed that the musicians only rarely outperformed non-musicians on WM tasks. As for memory structure, a principal components analysis revealed that the seven tasks loaded onto different components for non-musicians and musicians. In musicians, scores loaded onto three components that represent motor–visual memory, verbal memory, and memory for the movements of others. In contrast, there were only two extracted components for non-musicians. These results suggest that music training leads to greater cross-modal and intermodal integration in WM, as well as specialization within motor memory.
Working memory (WM) is a memory system that temporarily processes and stores incoming information for use of in-the-moment tasks (Baddeley & Hitch, 1994; Cowan et al., 2005; cf. Brown, Neath, & Chater, 2007). Researchers have proposed several models of WM, but commonly include a central component that involves executive functioning and somewhat distinct peripheral components for verbal and visual information (Baddeley, 1996, 2001, 2012; Baddeley, Allen, & Hitch, 2011; Cowan, Donnell, & Saults, 2013; Cowan et al., 2005; Cowan, Saults, & Blume, 2014; Gray et al., 2017 Miyake & Shah, 1999; Unsworth & Engle, 2007). Neuropsychological research has often supported these distinctions (De Renzi & Nichelli, 1975; Hampson, Driesen, Skudlarski, Gore, & Constable, 2006; Moses et al., 2002). The purpose of the present study is to test if music training affects the structure of WM. We consider two possible effects of music training on WM: (1) music training enhances memory capacity and (2) music training changes the functional structure of WM, particularly motor memory.
Music training could potentially affect verbal and visual memory components, as well as central components. Through exposure and practice with music, training could enhance memory for auditory information. Indeed, there is extensive evidence that music training improves auditory processing (Magne, Schön, & Besson, 2006; Musacchia, Sams, Skoe, & Kraus, 2007; Schön, Magne, & Besson, 2004), verbal processing (Moreno et al., 2008; Musacchia et al., 2007; Schön et al., 2004; Slater et al., 2015; Thompson, Schellenberg, & Husain, 2003), verbal intelligence (Moreno et al., 2011), phonological awareness (Flaugnacco et al., 2015; Gordon, Fehd, & McCandliss, 2015), literacy (Slater et al., 2014), and can even boost second language achievement in academic settings (Yang, Ma, Gong, Hu, & Yao, 2014). Previous studies have shown that musicians outperform non-musicians on verbal memory (Chan, Ho, & Cheung, 1998; Degé, Wehrum, Stark, & Schwarzer, 2011; Hanna-Pladdy & Gajewski, 2012; Hanna-Pladdy & Mackay, 2011; Ho, Cheung, & Chan, 2003; Roden, Grube, Bongard, & Kreutz, 2014; Roden, Kreutz, & Bongard, 2012). Because the effects of music training transfer to non-musical tasks, researchers have argued that music training might increase the plasticity in the left temporal lobe (Ho et al., 2003; see also Gordon et al., 2015), thereby leading to a musician advantage in auditory tasks generally. There is also some evidence that this plasticity may extend more generally than the left temporal lobe: Schön et al. (2004) found greater bilateral involvement in pitch processing among musicians relative to non-musicians (who showed greater left hemisphere involvement in processing pitch). These results suggest that musicians show a high degree of intermodal integration with regard to auditory stimuli, that is, they can transfer their trained-through-music auditory skills to a wide variety of other auditory-related tasks (Musacchia et al., 2007).
Music training could lead to similar intermodal integration across visual/spatial tasks, as many musicians learn to read notes on a staff. In support of this prediction, one study showed small and statistically significant increases in visual attention with musical training among children (Roden, Könen, et al., 2014). Similarly, children who experience 2 years of musical training showed improved visual memory relative to control children (Degé et al., 2011). Also, older adults with 10 or more years of music training have been shown to have better visuospatial judgment than non-musicians (Hanna-Pladdy & Gajewski, 2012; Hanna-Pladdy & Mackay, 2011). However, it is important to keep in mind that several studies found no evidence for musical training leading to greater visual memory (Ho et al., 2003; Roden, Könen, et al., 2014; Roden et al., 2012). Moreover, one study found that increases in the ability to perform spatial-navigational tasks associated with musical training were not consistent across time (Mehr, Schachner, Katz, & Spelke, 2013). Not all musicians learn to read music, so some of the variability could be related to individual differences in reading music. Nevertheless, taken together, these results suggest that there are, at best, small increases in visual memory linked to musical training.
In addition to intermodal integration, music training could lead to enhanced cross-modal integration, as musicians can simultaneously read notes, listen to the sounds they make as they are played, and translate the written notes into physical movements to play an instrument. Coordinating these processes may contribute to enhanced central executive functioning in WM. Indeed, some studies have shown evidence for greater central executive involvement among musicians. For example, in an 18-month longitudinal quasi-experiment, Roden et al. (2014) found that music training led to greater improvement than training in natural sciences over the course of the study on functioning of the central executive for school-aged children. Similarly, Ramachandra, Meighan, and Gradzki (2012) found effects of musical training on central executive functioning, which they measured using a backward digit span and a reading span task.
There is extensive evidence for musical training leading to both greater intermodal and cross-modal integration through increased neural plasticity (Groussard et al., 2014; Herholz & Zatorre, 2012). As noted earlier, some researchers have argued that this increased plasticity might be observed specifically in areas of the brain related to processing auditory information (Gordon et al., 2015; Ho et al., 2003; Musacchia et al., 2007). However, there is increasing evidence that the enhanced plasticity might also be observed in the interaction between sensory modalities (see review in Herholz & Zatorre, 2012), such as enhanced interactions between auditory and motor functioning (Chen, Rae, & Watkins, 2012; Lappe, Trainor, Herholz, & Pantev, 2011) and enhanced interactions across auditory and visual function (Elmer, Meyer, & Jancke, 2012).
The research testing for musician advantages in WM has thus far focused only on visual, verbal, and central executive components. Music training could also influence motor (or kinesthetic) memory (Wu & Coulson, 2014; Zhong & Stone, 2013), the memory for body positions and movements. Motor memory is essential in coordinating hand and body movements when playing an instrument. Motor memory is thought to differ slightly from procedural memory, which accounts for implicit, habitual sequences of movements (Fuchs, 2012; Simor et al., 2018). Motor memory has often been considered a subcomponent of visual memory (Baddeley, 1996, 2001, 2012). And, indeed, some studies have shown strong relationships between visual and motor memory. For example, Seidler, Bo, and Anguera (2012) found that visuospatial WM ability predicts an individual’s rate of motor learning for sequences. Other studies have found evidence for a noted degree of independence between visual and motor memory components (Jaroslawska, Gathercole, & Holmes, 2018; Smyth & Pendleton, 1990). One study with older adults showed effects of music training on motor dexterity (Hanna-Pladdy & Gajewski, 2012), but not particularly motor memory. Another study showed that music training was associated with changes in structure and processing in motor areas of the brain (Gaser & Schlaug, 2003), but not behavioral measures of motor memory.
In the present study, we also consider the possibility that music training leads to changes in the functional structure of WM, particularly with regard to motor memory. We know of no previous studies testing the possibility of structural changes in WM due to music training directly. However, developmental studies have often shown changes in the structure of WM with age in middle childhood and adolescence (Campos, Almeida, Ferreira, & Martinez, 2013; Hale, Bronik, & Fry, 1997; Lee, Bull, & Ho, 2013; Moses et al., 2002; cf. Michalczyk, Malstädt, Worgt, Könen, & Hasselhorn, 2013). In most cases, these studies report increasing integration across memory components with age, most likely due to increasing involvement of the central executive (Campos et al., 2013; Lee et al., 2013; cf. Moses et al., 2002). In any case, it is possible that these age-related changes in WM structure are related to skill acquisition and other experiences (see discussion in Campos et al., 2013). Thus, in the present study, we may see differences between musicians and non-musicians in the functional organization of their WM, with greater integration of memory components, both intermodally (i.e., across tasks from the same sensory modalities such as auditory or visual) and cross-modally (i.e., high interrelatedness across modalities). The latter results would be consistent with the research showing that music training affects cross-modal interactions in neural processing (Herholz & Zatorre, 2012).
This study
The purpose of this study was to test whether musicians differed from non-musicians with regard to WM capacity and structure. In terms of WM capacity, we predicted that we would replicate previous studies showing musician advantages in verbal tasks and tasks involving the central executive (Ramachandra et al., 2012; Roden et al., 2014). Some (but not all) previous studies have shown musician advantages in visual memory (Degé et al., 2011). As for WM structure, we predicted that visual and verbal WM tasks would constitute separately measurable components that are not highly inter-related for non-musicians, with motor memory tasks showing connections to visual memory (Baddeley, 2012). In contrast, the functional structure of WM for musicians would differ, perhaps showing stronger integration within and between verbal, visual, and motor memory components.
Method
Design
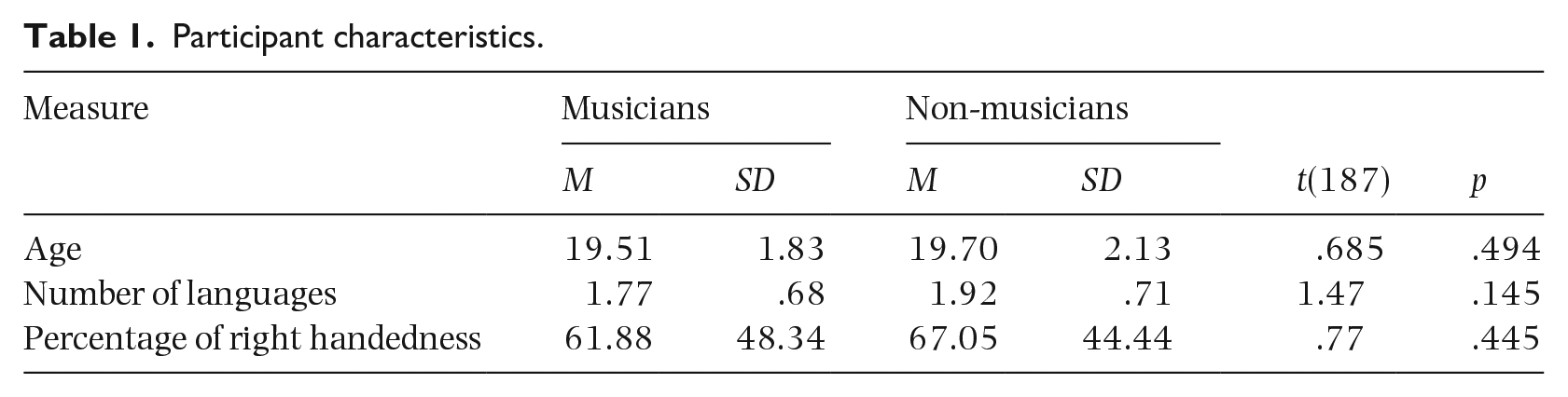
This study measured individuals’ scores on seven WM tasks. It also measured variables to account for individual differences among participants, including type and quantity of music training, handedness, 1 and languages spoken. Handedness was measured because hand dominance has been shown to affect higher order cognition, including executive function, which could implicate memory (Donoff, Madan, & Singhal, 2017). Previous studies have shown a link between bilingualism and performance on WM tasks (Bialystok, 2015) as well as music (Martínez-Montes et al., 2013). The number of languages spoken at a conversational level was measured for musicians (M = 1.77, SD = .68) and non-musicians (M = 1.92, SD = .71). There was no significant difference between the number of languages spoken between groups, t(187) = 1.47, p = .145.
Participants
One hundred ninety people participated in this study: 91 with musical experience (Mage = 19.51 years, SD = 1.83 years), and 99 without significant musical experience (Mage = 19.71 years, SD = 2.12 years). The participants with musical experience had at least 6 years of formal instrumental music training beginning before the age of 10. Participants without musical experience were those who had three or fewer years of formal instrumental music training.
One hundred eighty-two participants were recruited via the University of Alberta’s psychology research participation pool and received partial course credit for completion of the study. Snowball sampling was also used to recruit eight additional participants with music training. These participants received a Cdn$10.00 honorarium for their participation. All participants were either university students or had graduated from university within the last year and were between the ages of 17 and 29 (M = 19.61, SD = 1.98). See Table 1 for summary of participants’ background characteristics.
Participant characteristics.
Measures
Each participant completed seven memory tasks: forward and backward digit span tasks (for verbal memory), forward and backward Corsi block tapping tasks (for visual memory), a movement span task by Wu and Coulson (2014), and two versions of the test of ability in movement imagery (TAMI; Madan & Singhal, 2013). The last three tasks are measures of motor memory.
The forward digit span task is a verbal memory test in which the researcher reads a list of digits aloud to the participant at a rate of one digit per second. Participants then recall the digits to the best of their ability. The list of digits increases in length with each phase. The first phase consists of two digits, and reaches nine digits by the last phase of the task. The span increases by one digit with each successive phase. The backward version of the task begins with two digits and reaches a span of six digits. In the present study, only one version of the task was used, so all participants heard the same digits. Each participant completed every phase of the test, from spans of just two digits, to the full span of nine (or six in the backward version), regardless of their score on each phase. In the backward digit span task, participants had to repeat the digits in reverse order to what was read aloud. For both digit span tasks, each list was scored in an all-or-nothing fashion: any incorrect digit in the span resulted in a score of zero for that phase. Participants had to correctly recall all digits in the list to receive a score of one for that phase. The score represents the highest level that a participant achieved, with the highest possible score being 8 for the forward digit span and 5 for the backward digit span.
The Corsi block-tapping task was used as a measure of visual memory. Participants watched a short video in which blocks were highlighted in sequence on a computer screen. They were then required to tap the blocks on the screen back in the same sequence. There were two different trials of the same sequence length for each phase. This task had a discontinue rule, meaning participants could not move onto the next phase unless they got both trials completely correct for each phase. If participants tapped the first sequence in a phase incorrectly, the second sequence in the phase was still completed. All participants saw the same block sequences. Like the digit span task, this test was marked on an all-or-nothing basis. We included in the analyses the total score, namely the highest number of sequences correctly recalled. The highest possible score was 12. In the backward Corsi block-tapping task, the same procedure is carried out, but participants must recall the block sequence in reverse order. The backward Corsi block-tapping task also commences with two trials of sequences containing two blocks and works up to sequences containing a maximum of nine blocks, so the maximum score was 9.
Wu and Coulson’s (2014) motor memory task, the movement span task, is a psychometric measure consisting of five levels, with three trials per level. In the present study, each participant completed all levels of the task. On each trial, participants watched a short video of a camera-facing person performing an unconventional gesture on a computer screen, and then, at the signal of an auditory cue, participants turned toward the researcher to re-create this movement to the best of their ability by mirroring the video. Trials in level one contained one movement, and each successive level contained one more movement than the previous, so that trials in level five contained five movements. Following Wu and Coulson (2014), correct movements received one point, and movements that indicated recollection of the movements but were not completely accurate receive half a point. Participants could only receive a half point if they were missing only one component of the movement (such as incorrect orientation). If more than one element of the target movement was missing, a point was not awarded. The total score of the task reflects a sum of the points obtained across all trials for each level. Higher scores indicate greater motor memory capacity, particularly for the body movements of someone else. Wu and Coulson also calculated participants’ span score, which is calculated by awarding one point for every level at which they achieve at least half of the available points, plus half a point for each subsequent level that they obtain at least half of the available points after failing to obtain at least half of the points on one level. Participants were videotaped while performing this task to ensure accuracy of scoring and high interrater reliability. To reduce the effect of technical difficulties and missing trials due to camera malfunction on some participants’ scores, the percentage of total correct movements for each participant was calculated and used in all analyses. Two independent raters coded the videos from the Wu and Coulson movement span task. Each rater also coded two videos that had been coded by the other rater. Score differences between the videos coded by both raters were no greater than one point. The mean rating for Coder 1 was 15.8 and for Coder 2 was 15.6. The correlation between the two coders was high and significant, r(2) = .989, p < .02. Wu and Coulson (2014) found the movement span measure to be stable across time.
The TAMI consists of 10 multiple choice questions in which participants are shown an image of a body position and must imagine several explicit body actions, then select the appropriate final body position (Madan & Singhal, 2013). Participants were scored out of 10, with higher scores indicating better imagined motor imagery ability. Madan and Singhal (2013) found the TAMI to have good concurrent validity when it was compared with pre-existing measures of movement imagery, indicating that the test also has good construct validity. In the present study, the TAMI aims to examine motor WM, as participants must imagine multiple body movements while retaining and mentally manipulating the current body position with each additionally imagined movement to arrive at the correct final body position. Unlike the movement span task, the TAMI asks participants to imagine their own body movements.
The second version of the TAMI used in our study was the test of ability in hand movement imagery (TAHMI; Donoff et al., 2017), which is a modified version created by Christopher Donoff specifically for imagined imagery of the hand. In the present study, the TAHMI was adapted to contain five questions for each hand. The questions consisted of functionally involved questions in which participants were instructed to select the object that they could most compatibly utilize with their imagined hand position. Participants held a tennis ball in their hand while completing the TAHMI to prevent overt hand movements from occurring, since the goal of the task was to elicit imagined imagery. Four different versions of the TAHMI were used. To control for order effects, some participants saw right-handed questions first, while others saw left-handed questions first. To control for potential differences in difficulty between right-and left-handed questions, the right- and left-handed questions were all flipped to the other hand. Performance differences were not significant between TAHMI versions.
To measure handedness, each participant filled out the Edinburgh Handedness Inventory. This inventory consists of 10 questions that ask participants to rate their hand preference on common tasks such as writing, throwing, and using utensils (Caplan & Mendoza, 2011). For each named task, participants are asked to place either one or two checkmarks for the right–left hand. These checkmarks indicate the strength of their hand preference, with two checkmarks meaning that they would never use the other hand unless forced to. The Edinburgh Handedness Inventory is scored using laterality quotient: LQ = (right—left)/(right + left) × 100. The LQ scores indicate participants’ percentage of right-handedness (Donoff et al., 2017). The groups did not differ significantly in terms of their right-handedness: on average, musicians had an LQ of 61.88%, and non-musicians had an LQ of 67.05%.
Procedure
In the current study, participants participated in all seven memory tasks: forward and backward digit span, forward and backward Corsi block tapping, the movement span task, the TAMI, and the TAHMI. All seven tasks were administered by the same researcher for each participant for consistency. Tasks were counterbalanced to minimize possible order effects. Participants were randomly assigned to one of four task sequence versions. Subsequent analysis indicated that there were significant differences between task sequence versions for the forward digit span (p = .034) and the backward Corsi block task (p = .022). Since these differences were only present for two of seven tasks, we disregarded differences between task sequences in subsequent analyses.
Data analysis
T-tests were used to examine differences in memory capacity between musicians and non-musicians. Principal component analysis (PCA) was also chosen to analyze the data, since the other goal of this study was to examine the interrelatedness of memory components in musicians and non-musicians. To account for the task-difficulty-level differences, the PCA was performed on standardized scores rather than raw scores.
The factorability of seven factors (forward digit span, backward digit span, forward Corsi block, backward Corsi block, TAMI, TAHMI, movement span task) was examined separately for musician and non-musician groups. The Kaiser–Meyer–Olkin (KMO) value, which indicates how suitable a data set is for PCA, was .766 for the non-musician group, above the recommended minimum value of .5 (“KMO and Bartlett’s test,” n.d., para. 1). Values closer to 1 indicate better suitability for PCAs. Bartlett’s test of sphericity was significant for the non-musician group, χ2(21) = 82.46, p < .001, which is another indicator of a data set’s suitability for PCA. For the musician group, the KMO value for the PCA was .631, which is above the minimum recommended value of .5. Bartlett’s test of sphericity was also significant for the musician group, χ2(21) = 70.44, p < .001.
Results
Group comparisons for memory capacity
Data were analyzed using SPSS statistical software (IBM Corp. Released, 2017). To first examine the hypothesis that musicians have greater memory capacity than non-musicians, independent sample t-tests were performed for scores on all seven memory tasks to compare musician and non-musician groups. For analysis, the total scores rather than the span scores were used for both the Corsi block and movement span tasks, as those scores were more sensitive measures of memory ability than their accompanying span scores. There were significant differences in scores between musicians and non-musicians on the forward Corsi block task, t(187) = 2.06, p = .041, d = .300, and the TAHMI, t(188) = 2.42, p = .016, d = .348, with musicians performing significantly better than non-musicians in both cases (Table 2). The differences between musicians and non-musicians were non-significant for scores on the backward Corsi block, forward digit span, backward digit span, TAMI, and the movement span task (Table 2). These results demonstrate that there are a few small differences in memory capacity between musicians and non-musicians.
Means and t-test results for task scores between non-musician and musician groups.

TAMI: test of ability in movement imagery; TAHMI: test of ability in hand movement imagery.
p < .05.
Inter-relatedness of memory components
The correlations between the seven memory tasks are summarized in Table 3. As can be seen in that table, the scores for the seven variables measured were small to moderately inter-correlated for both musicians and non-musicians. Scores that were expected to be related, such as the forward and backward digit span, and several measures of visual and motor memory were correlated as expected (Table 3). The correlation between the forward and backward digit span was larger in musicians, r = .389, p < .01, than non-musicians, r = .218, p < .05. The correlation between two motor memory measures, TAMI and TAHMI, was also larger for musicians, r = .400, p < .01, than non-musicians, r = .233, p < .05.
Correlations between memory tasks for musicians and non-musicians.

TAMI: test of ability in movement imagery; TAHMI: test of ability in hand movement imagery.
Note: Top of matrix above diagonal indicates correlations for non-musicians, bottom of matrix below diagonal indicates correlations for musicians.
p < .01.
p < .05.
Non-musician group
The structure of musicians’ and non-musicians’ memory systems was examined with a PCA. The communalities in the model, represented by extracted values, indicate accounted variability within each variable (see Table 4). For the non-musician group, communalities for all seven factors were above .30, and five of seven factors were above .40, which indicates that all factors shared some variance with at least some of the other factors. For non-musicians (Table 4), the variable that played the largest role in explaining overall variability was forward digit span (.694), followed closely by backward Corsi block (.618). Three variables in the non-musician group had extraction values between .40 and .60: the forward Corsi block (.518), backward digit span (.477), and the movement span (.439). Two variables had extraction values less than .40 (TAMI; .379 and TAHMI; .386).
Communalities extraction.

TAHMI: test of ability in hand movement imagery; TAMI: test of ability in movement imagery.
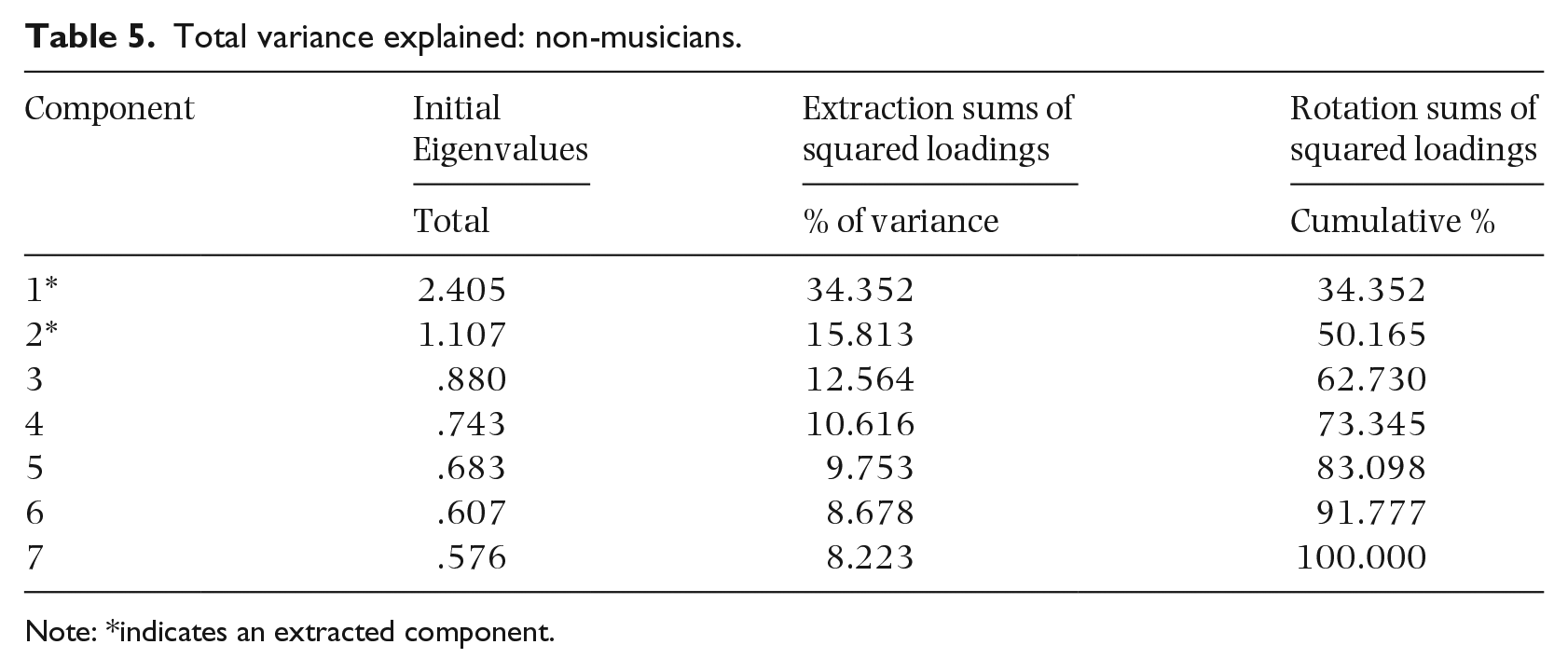
Eigenvalues are coefficients in PCA that represent covariance in the data set (Nicholson, n.d.). Larger eigenvalues represent the most significant underlying principal components in the data set. In our study, eigenvalues above one from the PCA were extracted, resulting in two extracted components for the non-musician group, which accounted for 34.35% and 15.81% of the variance, respectively (Table 5). These two components accounted for 50.17% of the variance cumulatively. The remaining five factors were not extracted, as they were not deemed to contribute sufficiently to the variability of the model. A varimax rotation with Kaiser normalization was used to examine how data from the original seven variables could be simplified by grouping them according to the factors that were extracted for musicians and non-musicians. In non-musicians, the first factor-loading component of the standardized PCA is best explained by the movement span task, backward digit span, TAHMI, forward Corsi block, and backward Corsi block (Table 7). The second component is best explained by the forward digit span and the TAMI. For the non-musician group, the two components that the seven factors load onto cannot be easily named: they do not appear to represent separate modality components, nor do they represent a central/peripheral distinction.
Total variance explained: non-musicians.
Note: *indicates an extracted component.
Musician group
Musicians differed from non-musicians in the magnitude at which the seven variables were extracted. For musicians (Table 4, rightmost column), the variable that played the largest role in explaining overall variability was the movement span (.768), followed closely by the forward digit span (.757) and backward digit span (.733). The remaining four variables had extraction values between .40 and .80 for the musician group: the TAHMI (.700), backward Corsi block (.515), TAMI (.508), and forward Corsi block (.415).
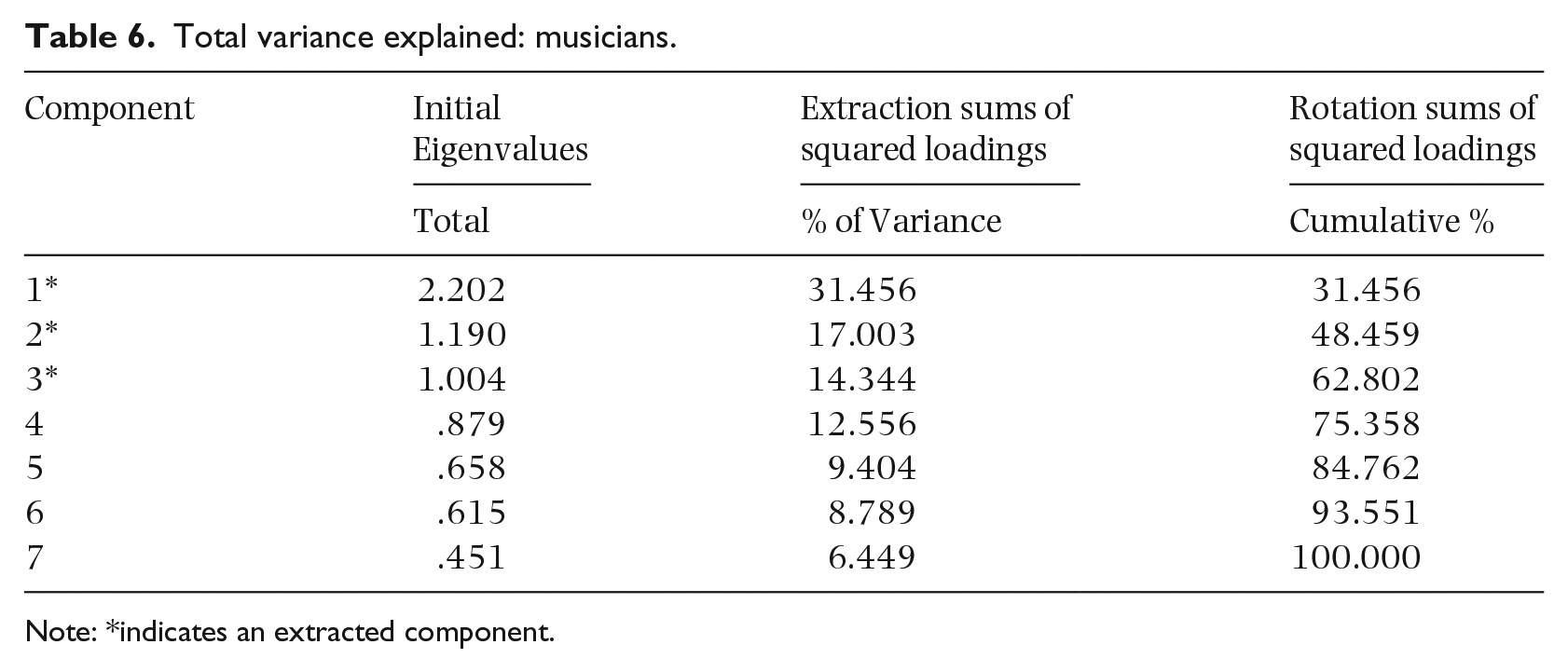
Eigenvalues above one from the PCA were extracted, resulting in three extracted components, which accounted for 31.46%, 17.00%, and 14.34% of the total variance, respectively (Table 6). These three factors cumulatively accounted for 62.80% of the variability in the musician group. Of the three components that factors loaded onto in musicians (Table 8), the first component is best explained by the TAMI, TAHMI, forward Corsi block, and backward Corsi block; the second component can be explained by the forward digit span and backward digit span; and the third component can be explained by the movement span. These three components appear to represent motor–visual memory, verbal memory, and memory of the body movements of others, respectively. Small components with coefficients smaller than .50 were suppressed in the PCA.
Total variance explained: musicians.
Note: *indicates an extracted component.
Discussion
The goal of the present study was to test if there were differences between musicians and non-musicians in terms of WM capacity and structure. Musicians and non-musicians were compared on seven memory tasks that captured visual, verbal, and motor memory domains, including forward and backward Corsi block tasks, forward and backward digit span tasks, the movement span task, the TAMI, and the TAHMI.
The results indicated that musicians had significantly greater memory capacity for the forward Corsi block task (visual memory) and the TAHMI (motor memory). Differences between musicians and non-musicians were non-significant for all other memory scores. We did not replicate the musician advantage for verbal WM and central executive functioning that has been found in previous studies (Hanna-Pladdy & Gajewski, 2012; Hanna-Pladdy & Mackay, 2011; Ramachandra et al., 2012; Roden et al., 2014). In the present study, even when we observed differences, the effect sizes were small (< .40). These results show that musicians show, at most, small differences from non-musicians in their memory capacity.
We then turn to the second hypothesis: musicians and non-musicians differ in terms of functional memory structure. We had predicted that musicians would show differences from non-musicians on WM structure, both in terms of intermodal and cross-modal integration (Herholz & Zatorre, 2012). However, the specific memory tasks did not quite load onto the components as predicted.
The non-musicians in our study did not show the distinction between verbal and visual memory, with motor tasks loading on to visual memory, as many previous studies have shown (Baddeley, 2012; Campos et al., 2013; Cowan et al., 2013; Gray et al., 2017; Michalczyk et al., 2013). In fact, the two derived components from the PCA do not correspond neatly to verbal and visual memory (with or without motor memory; see Table 7). One possible explanation of this is that the structure of memory processes is highly variable in non-musicians. In line with this explanation, some previous studies have included sample sizes well over 1000 to show a clear verbal/visual memory distinction (Michalczyk et al., 2013).
Rotated component matrix: non-musicians.

TAMI: test of ability in movement imagery; TAHMI: test of ability in hand movement imagery.
In contrast, the musicians showed a different WM structure, with factors loading onto three components, representing motor–visual memory, verbal memory, and memory for the body movements of others (Table 8). The strong connections between motor and visual memory are consistent with cross-modal integration (Herholz & Zatorre, 2012), although it was somewhat surprising to observe the relative independence of verbal memory from motor memory (cf. Chen et al., 2012; Lappe et al., 2011). Learning to play an instrument could lead to strong connections between remembering how to move in response to reading music.
Rotated component matrix: musicians.

TAMI: test of ability in movement imagery; TAHMI: test of ability in hand movement imagery.
In addition to evidence for enhanced cross-modal interactions between motor and visual memory in musicians, we also found evidence for enhanced intermodal interactions. The strong connection between the two visual memory tasks, two motor memory tasks (the TAMI and the TAHMI), and the two verbal memory tasks is consistent with previous research showing a high degree of intermodal integration and plasticity among musicians (Degé et al., 2011; Groussard et al., 2014; Musacchia et al., 2007).
One surprising finding was that, for musicians, the movement span task was independent from the other memory stores, including motor memory. Recall that the movement span task assesses participants’ memory for the movements of other people rather than their own movements (like the TAMI and the TAHMI). It is possible that musical training enhances remembering independently whether the musician is doing a movement or whether someone else (perhaps the teacher or the conductor) does a movement. To our knowledge, this degree of specialization within motor memory has not been observed in previous research. Future studies can test the replicability of this finding.
Limitations and future directions
There are a number of limitations to the present study that could guide future research. First, this study compared musician and non-musicians, inferring that any differences were due to musical training. However, it is equally possible that people with a particular memory structure are highly drawn to music (see discussion in Swaminathan, Schellenberg, & Khalil, 2017). While this interpretation seems unlikely given the existing longitudinal work showing similar effects of musical training (Roden et al., 2014), longitudinal research in which participants are assigned to music training and control conditions over several months or years should also be conducted to show causality between music training and changes in memory structure.
Another limitation of the present study is that music experience could be confounded by musicians’ efforts during lessons and practice. The present study asked about years and type of music training, but did not inquire about time spent practicing each day or week. Achievement level and the number of hours spent practicing might also influence the degree of cognitive change in WM. The present study does not allow for these types of conclusions to be drawn, but future research can include measures of practice and effort.
Yet another limitation of the present study is that we did not control which instrument(s) the musicians played. Participants with music training (n = 91) reported that 81.3% of participants played the piano, 27.5% played a string instrument (violin, viola, or cello), 25.3% played the guitar, and 52.7% of participants played some other instrument (including brass, woodwind, and cultural instruments). Many participants fell into more than one of these categories, as many musicians have training in more than one instrument. For this reason, it was not possible to separately analyze participants by instrument type, since it would be difficult to disentangle the potential influence of playing multiple instruments. The various types and styles of training that accompany different types of instruments might influence memory structure in different ways. Future research should examine whether there are additive effects of playing multiple instruments, or instruments from multiple domains (such as playing a string instrument and a woodwind instrument). Since such a large proportion of participants reported playing the piano, it is reasonable to conclude that piano training has influences on memory structure as reflected in the results of this study. Indeed, research has shown positive effects of piano lessons on verbal processing and executive functioning in both children (Piro & Ortiz, 2009) and older adults (Bugos, Perlstein, McCrae, Brophy, & Bedenbaugh, 2007).
Conclusion
The current study investigated differences between musicians and non-musicians on memory capacity and structure. We found only small differences on capacity (with musicians outperforming non-musicians). We found differences in memory structure, with musicians showing cross-modal integration (motor and visual memory), intermodal integration (within motor memory, visual memory, and verbal memory, respectively), and specialization (differences between memory of one’s own movements and memory of others’ movements). The differences in memory structure in musicians and non-musicians found in this study suggest that music training has important cognitive impacts. The experiences and practice with bodily movements, vision, and hearing that are inherent to musical training could lead to an enhanced ability to flexibly use particular modalities to do non-music tasks. Future longitudinal studies will test whether musical training does, indeed, play a causal role in the development of a different memory structure.
Footnotes
Acknowledgements
The authors would like to thank all of the research assistants who made this study possible by running participants and coding data: Maria Chow, Mariam Dar, Sunny Dhonkal, Beverly Michel Baluyot, Gurishar Dami, Mathew Gorman, Nicole Love, Anna Kuc, Habeebah Mohammad, Zaahidah Ali, and Tehseenah Zahrah. Christopher Donoff shared the most recent version of the TAHMI. Shiva Zarezadehkheibari and Christopher Westbury gave them helpful feedback regarding data analysis.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study received funding from a Discovery Grant to the second author from the Natural Sciences and Engineering Research Council of Canada.