
Abstract
Microbial life has been much in the news. From outbreaks of Escherichia coli to discussions of the benefits of raw and fermented foods to recent reports of life forms capable of living in extreme environments, the modest microbe has become a figure for thinking through the presents and possible futures of nature, writ large as well as small. Noting that dominant representations of microbial life have shifted from an idiom of peril to one of promise, we argue that microbes – especially when thriving as microbial communities – are being upheld as model ecosystems in a prescriptive sense, as tokens of how organisms and human ecological relations with them could, should, or might be. We do so in reference to two case studies: the regulatory politics of artisanal cheese and the speculative research of astrobiology. To think of and with microbial communities as model ecosystems offers a corrective to the scientific determinisms we detect in some recent calls to attend to the materiality of scientific objects.
Microbes were around for billions of years before humans and will likely continue to thrive long after we pass away. All of this is, in fact, some great comfort, as we could certainly not survive without them. They are the recyclers of the planet, invisibly keeping life going, and they are silent partners in our bodies … Now, reader, every bite of cheese … can have new meaning for you. Some think that extremophiles might give us insight into the forms of life that might exist on some hot, or cold, or acidic planet far off in space.
Call them millennial microbes. Across a range of scientific and popular arenas, particularly in the United States, the early 2000s saw microbes emerge as models and media for transformative food politics, new ecological and biomedical futures, and even studies of life as it might exist on other worlds. At a time when biological nature writ large is widely understood to be under threat from such anthropogenic processes as climate change, agri-industrial degradation, rain forest depletion, and overfishing, biological nature writ small – that is, microbial life – has attracted fresh attention. Ecologists have become interested in the microbial dimension of ecosystems, while microbiologists are moving beyond their Petri dishes to consider how microbial communities operate in soil, ocean, and air environments, where they mediate climate, remediate pollution, and might even provide raw material to create new biotechnologies, such as biofuel (see Pottage, 2006). As microbiologists have turned to gene sequencing to characterize the realm of the tiny, even the human body has become legible as a consortium of microbes, with the Human Microbiome Project promising to characterize the 90 percent of cells in the body that are microbial (Nerlich and Hellsten, 2009), looking toward mining what the National Institutes of Health (2013) have called ‘an area of great potential for benefits to human health’. Across diverse domains, microbes – or better, microbial ecosystems – have come into view as plenteous, promising, full of potential.
Scholars in Science and Technology Studies (STS) have taken note of this microbial moment, analyzing the emergence of novel classification systems for microorganisms (Helmreich, 2003; O’Malley and Dupré, 2007; Sommerlund, 2006), the arrival of a biopolitics that includes microbially mediated processes of infection and digestion (see Paxson, 2008 on ‘microbiopolitics’), and the coming into consequence of unexpected bacterial, fungal, and viral agencies in lab and field science (Helmreich, 2009; Hird, 2009; Ingram, 2011; Schrader, 2010). Whether manifesting as single cells or as cellular assemblages, microbial nature materializes and models a world in which boundaries – taxonomic, metabolic, ontogenetic – are breached between humans, animals, plants, fungi, protoctists, and their bacterial and archaeal familiars and unfamiliars (Sagan, 2011). The microbial turn in recent biology, we suggest, marks the advent of a newly ascendant model of ‘nature’, one swarming with organismic operations unfolding at scales below everyday human perception, simultaneously independent of, entangled with, enabling of, and sometimes unwinding of human, animal, plant, and fungal biological identity and community (see McFall-Ngai et al., 2013). 1 Microbes are not tokens, as were the late-20th-century reductionist genome and (putatively) carbon copy clone, of the ‘age of biological control’ (Franklin, 2007; Wilmut et al., 2001), but are rather pointers to a biology underdetermined and full of yet-to-be explored possibility.
In this essay, we examine such models and document how, in contemporary representations of and practices around biotic nature, the abundant microbe has moved from being a sign of peril to being also one of promise. We do so with respect to two cases: one, concerning food, in which microbes constitute a nature that may be intimate with and even incorporated into humans’ own, and another, around the search for extraterrestrial life, in which microbes offer the idea of a biotic nature quite remote from us earthlings. In each case, microbial abundance is assayed for its diversity as well as with respect to its generality – its widespread and suffusing presence, in both daily and distant domains.
Our first case takes up food politics, in which microbial life has come into a new focus for regulators, producers, retailers, and consumers (e.g. Nestle, 2003). Whereas hygienists, food safety regulators, and consumers since Louis Pasteur have largely associated food-borne microbes with pathogens, in the past decade, new attention and even popular appreciation is being showered on those microbes essential to the production, edibility, and healthiness of fermented foods (e.g. Katz, 2003, 2012; Pollan, 2013). Building on the US-based fieldwork of one of us, Heather Paxson (2013), we look particularly at the microbial life of artisanal cheese. Microbial elements of artisanal cheese have come into prominence in recent regulatory and industry debates over how best to square the potential microbial hazards of handcrafting cheese, particularly from raw (unpasteurized) milk, with the gustatory delight of eating it (e.g. Kurtz et al., 2013; Neuman, 2011; Paxson, 2008, 2013). 2 While the North American regulatory future of raw-milk cheese remains uncertain, the debate has taken the microbial constitution of cheese to center stage, and cheese has become a popular medium for thinking about how microbes contribute to the distinctive goodness, healthful as well as tasty, of a fermented food. Microbes are everywhere in and around cheese, but just as only certain ones are pathogenic to humans, only certain strains, species, or communities are held up as models that might underwrite claims to distinctive flavors or to health benefits for humans. 3 The embrace of microbial promise must still keep microbial peril at bay.
Our second case takes us off world to speculate on microbial life on other planets, in extraterrestrial settings of interest to scientists in astrobiology, the study of life as it might exist elsewhere in the universe. Drawing on the fieldwork of the other of us, Stefan Helmreich, we examine how the possible existence of microbes on other worlds has gone from a source of anxiety about interplanetary contamination to a prompt for hopeful speculation about extraterrestrial life. In astrobiological investigations of the limits of life, microbes may be everywhere around us, but only certain strains, species, or communities may be used as models that might reveal something about the general or universal properties of life.
Both our cheese and extraterrestrial examples have people enrolling microbial life in discussions of new potentials latent in organic, biotic nature. But microbes do not embody such potentials in and of themselves; rather, microbes materialize in cultural, social, political, and scientific practices, and thence reformat and reshuffle those practices in unexpected ways – a claim Bruno Latour (1988) advanced already in The Pasteurization of France. But we can modify that Latourian claim for the contemporary moment. Since the 19th-century Pasteurian period of which Latour wrote, microbes have come to be differently apprehended, no longer now as primarily frightening but also as newly appealing; microbes are also differently comprehended, no longer now only as individual strains but also, increasingly, as communities, as entities that matter within diverse ecosystems.
It is this relational angle that has made microbes newly attractive – good to think and work with – because scientists can imagine microbes to be full of unexpected, uncharacterized possibility as well as pliable to cultural projects. Microbes have become tokens of potentiality (see Taussig et al., 2013). Like model organisms, microbes are ‘both natural and artificial objects’ (Ankeny and Leonelli, 2011: 315; on the ‘artificial’: observe that the very word microbe (‘small life’, more or less) implicates artifice, because of the culturally crafted technological measures of scale and means of observation needed to make microbes manifest). Such a natural/cultural reading of microbes, of course, fits them snugly into a modernist ethos that figures nature as both born and made (cf. Rabinow, 1992). But microbes are also unlike model organisms in that they are often mobilized in ways that rely explicitly on their multiple relationalities and openness – to one another, to other organisms, and to humans. They are good candidates for forming, as some of our scientist interlocutors would say, model ecosystems.
In each of our cases, we find people sorting through microbial abundance in order to discover and model both the unique and the universal. The unique and the universal operate as figure and ground, often switching places to bring microbes into legibility. Some cheesemakers are becoming curious about microbial uniqueness (is this strain or this microbial community found only here?) to ask whether newly characterized microbes contribute to distinctive cheeses in their generation of qualities particular to a cheese’s place and method of fabrication. Astrobiologists, meanwhile, are curious about universality (is this kind of microbial life possible to find everywhere?), asking whether newly characterized microbes point to qualities general to life itself. The practice of toggling between the unique and the universal affords the possibility for microbial ecosystems to be enlisted into newly expansive projects of both naturalizing cultural practice and culturing nature.
At a moment when public intellectuals warn of the ‘end of nature’, in science writer Bill McKibben’s (2006 [1989]) phrase, such ecosystems offer to a variety of scientists and laypeople new hopes of a not yet tamed or fully known nature, a future nature. In this way, microbes and microbial communities are model biologies not only in the canonical sense documented by scholars in STS – entities, often standardized, that can be probed to describe biotic properties both enduring and surprising, or that can be tested to generalize from simpler to more complex organisms (see Ankeny and Leonelli, 2011; Creager, 2002; Kelty, 2012; Kohler, 1994; Rader, 2004; Ramsden, 2011) – but are also relational models in a more prescriptive sense, tokens of how organisms and human ecological relations with them could, should, or might be. The two reframings are linked; the repackaging of microbes as parts of ecosystems, as entities that enable and substantiate new relations, is what gives spirit and body to the promises and prescriptions that attach to claims about microbial natures.
On materiality, microbes, and models
By focusing our attention on the many lives of microbes, we do not wish to be read as making a claim that the materiality of microbes is newly consequential. We wish – somewhat programmatically – to distance ourselves from some of the new materialisms that have been percolating through recent science studies.
From Jane Bennett’s (2010) call in Vibrant Matter to recognize the vital agencies inherent in materials such as trash, metal, and oil, to the rising trend in much STS to ask after ‘ontology’ (see Van Heur et al., 2013 for a bibliometric analysis), to moves in ‘affect theory’ (Massumi, 1996; Sedgwick, 2003; Whatmore, 2002) to theorize a pre-social (sometimes brain-based) ground of feeling and action (for critiques see Hemmings, 2005; Martin, 2013), several scholars in STS have invoked notions of materiality and ontology in trying to sail between the Scylla of old-fashioned realism and the Charybdis of social construction (see also Alaimo, 2010). Taking up the puzzle of materiality usefully presented by Judith Butler (1993) in Bodies that Matter, adapting Annemarie Mol’s (2002) call to catalog the multiple ontologies precipitating from bodily experience (compare Thompson, 2005 on the ‘ontological choreography’ realized in the practices of assisted reproduction), and working with Karen Barad’s (2007) notion of agential realism, some corners of STS thus find themselves in a place that we think is analytically productive but also risky. It is productive because it can shake thinking away from the certainties of social determinism, as exampled, canonically, in the Strong Programme in the Sociology of Knowledge – and because it can show that phenomena emerge in practice (compare Law and Lien, 2013). But it is also risky, because new materialist tactics often veer toward universalizing metaphysical claims about the nature of ‘matter’ as such and also, at times, take scientific truth claims about the world at face value – a move that we consider a step backward for STS.
To offer one example, consider Jane Bennett’s (2004) claim that ‘Thing-power materialism figures materiality as a protean flow of matter-energy’ (p. 349), which posits that the material world is best described using the lexicon of physics. Bennett is clear that her attempts to invoke a world beyond human signification are speculative, so she probably would not disagree that analytic difficulties begin as soon as she puts this description forward. An STS or history of science approach would keep up the questioning, asking how ‘matter-energy’ arrives as an available turn of phrase at all, emerging as it does as a particular ontology consequent on the 19th-century rise of electromagnetic and thermodynamic science.
A framing similar to Bennett’s appears in New Materialisms: Ontology, Agency, and Politics (Coole and Frost, 2010). In that volume, editors Diana Coole and Samantha Frost exhort readers to engage in ‘an ontological reorientation that is resonant with and to some extent informed by, developments in natural science’ (pp. 6–7), a reorientation that takes on board Einstein’s account of the atomic world as a zone of ‘constant emergence, fluctuation, and the shifting of nodes of charge’ (p. 11) and that aligns with new biological views of organisms as ‘produced through complex interaction between genes and a host of other factors such as hormones, neurochemical stimuli, dietary intake, and environmental conditions’ (p. 17). Such a ‘new materialism’ accepts science-made rather than science-in-action (Latour, 1987) as the basis of materiality. 4 As Tim Ingold (2007) argues in his ‘Materials against materiality’, there is also an epistemological hazard in making general claims about ‘material’ without regard either for its variety (physical, chemical, biological, and more) or for the historically realized properties of such things as rocks, trees, or microbes (see also Ingold, 2012). He worries that the new materialism is too often ‘expounded in a language of grotesque impenetrability on the relations between materiality and a host of other, similarly unfathomable qualities, including agency, intentionality, functionality, sociality, spatiality, semiosis, spirituality and embodiment’ (Ingold, 2007: 2). To be sure, one does not want to leave abstractions out entirely, since these, too, are things. But a ‘materiality’ that is only apprehended through other abstract nouns misses a chance to follow materials in practice. Steve Woolgar and Javier Lezaun (2013) put an allied argument succinctly in a recent issue of this journal: ‘“materiality”, just as “context” and its cognate terms, needs to be understood as the contingent upshot of practices, rather than a bedrock reality to be illuminated by an ontological investigation’ (p. 326).
As nonhuman, nonanimal, biological entities, it is not surprising that microbes have become appealing materials for STS. Their oscillating detachment from and entanglement with human enterprise would seem to make them ideal actants: at once brought into view through human signification (microscopy, taxonomy) as well as oblivious to those practices.
5
Microbial being has inspired scholars to think anew about embodiment (Hird, 2009), agency (Ingram, 2011), and time (Schrader, 2010). That thinking has both prefigured and tweaked recent calls to conduct ‘multispecies ethnography’ (Kirksey and Helmreich, 2010), incorporating animal, plant, fungal, and microbial bodies and doings into social analysis. But it is important to bear in mind that microbial being does not speak for itself. So, when Myra Hird (2009), in The Origins of Sociable Life, a sociological ethnography of a microbiology lab, writes that prior to Pasteur, bacteria were ‘already fully-fledged actants’ (p. 35) and that ‘actants always exceed their mediated understanding’ (p. 36), she seems to treat microbes as obviously durable natural kinds. While Hird’s call to ‘meet … microbes halfway’ (p. 54) offers a salutary openness to phenomena not fully captured by representation, her recounting of what her microbiologist informants say often takes their accounts as truth: ‘Bacteria have further developed complex engineering capabilities as an adaptation to conflicting environmental constraints’ (p. 46). Philosopher Maureen O’Malley’s (2011) review of Hird’s book describes the difficulty:
By removing from the picture how the science was done as well as the ongoing revisability and contestability of microbiological findings, we are left with the sense that there is a fount of straightforwardly produced and accepted knowledge from which we can drink. (p. 129)
Our colleague Cristina Grasseni (2013, personal communication) suggested that this move might be called not the ‘new materialism’ but rather the ‘new reductionism’.
In what follows, we treat microbes not as bare truth-tellers, but rather – and in conversation with our ethnographic interlocutors – as model ecosystems. If ‘model organisms are always taken to represent a larger group of organisms beyond themselves’ (Ankeny and Leonelli, 2011: 318), microbial communities figured as model ecosystems are, as Hannah Landecker has put it (personal communication), ‘little stages for the irruption of microbial ways of being into human life’. Microbes are made to signify larger biological worlds and socialities, wider perils and promises, in worlds imagined yet to come. At stake for STS scholars is how to think carefully about how things and humans combine to create objects and agencies. We must continue in our accounts to resist and grapple with what Donna Haraway (1989) once named the ‘temptation’ of ‘the siren call of the scientists themselves’ (p. 8).
We mean in this essay to contribute to STS discussions of model organisms both a recognition that especially when it comes to microbes, scientists are increasingly thinking ecosystemically, as well as a suggestion that these scientists are doing so with explicitly normative aims. Centering our attention on model ecosystems, we describe models not just as representatives, standards, or experimental objects, but also as moral exemplars – models that are not simply descriptive, but that might simultaneously be prescriptive. If, as Ankeny and Leonelli (2011) write, ‘model organisms serve as models for whole, intact organisms’ (p. 318), the ‘whole’ that microbes call forth in the examples we discuss here are ‘wholes’ – ecosystems – that embrace human cultural projects and curiosities. Microbes permit artisan cheesemakers and their fans as well as astrobiologists and their publics to think and work through the ratios of the unique and the universal in gastronomical and astronomical projects. They thus offer not only model ecosystems, but also model ecologies – studies in how to frame the world.
The case of artisanal cheese
The raw, the perilous, and the promising
Microbiologically speaking, cheeses – a perishable food – are ecosystems of bacteria, yeasts, and molds functioning within lactic environments sustained by human care. Such microbial ecosystems can be hospitable to human pathogens. On 21 October 2010, federal agents locked down Estrella Family Creamery in Washington State, suspending trade of their award-winning, raw-milk cheeses. A federal judge ordered the seizure after the Estrellas refused to initiate a total recall of their cheese in light of evidence produced by US Food and Drug Administration (FDA) officials of the ‘persisting presence’ in one of their cheese-aging rooms of Listeria monocytogenes (a bacterium that can cause listeriosis, which has been linked to miscarriage and stillbirth when ingested by pregnant women). Also in Washington, in December 2010, federal officials demanded that cheesemaker Sally Jackson upgrade the equipment she had been using on her farm for 30 years. Unable to afford the renovations, Jackson retired after her raw-milk cheese was linked to eight cases of illness borne by E. coli.
For the FDA, such cases (and there have been others of late) provide a warrant for revisiting the regulation of raw-milk cheese in the United States. Since 1949, after an outbreak of typhoid was traced to cheddar infected by Salmonella and made with unpasteurized milk, the FDA has required that cheese, to be legally saleable, must either be made from pasteurized milk or be aged at least 60 days at no less than 1.7°C. US regulation presumes that pathogenic control will be accomplished one of two ways: either by pasteurizing milk to knock out potential pathogens before cheesemaking begins, or by reducing the moisture and increasing the acidity of a cheese through aging, thereby contributing to an environment inhospitable to such pathogens as E. coli and Salmonella. As a means of food safety regulation, however, the 60-day rule looks increasingly obsolete, owing in part to the virulence and lethality of pathogens linked to food-borne illness or characterized since 1949, including Listeria monocytogenes and enterohemorragic E. coli O157:H7. The legal future of the 60-day aging rule, and therefore of domestic raw-milk cheese, is uncertain. In 2011, David Acheson, a ‘former FDA associate commissioner’, told a National Public Radio (NPR) correspondent that if aging cheese made from raw milk proves insufficient to kill pathogenic bacteria, ‘[t]hen, obviously, the next step is making pasteurization an absolute requirement’ (Standen, 2011).
US regulators are turning renewed attention to raw-milk cheese just as – and likely because – the number of artisan cheesemakers has grown exponentially; among the roughly 450 producers operating today, around 50 percent make less than 10,000 pounds of cheese per year, and over half work with unpasteurized milk. 6 Pitted against an industrially based regulatory order bent on taming an unruly nature through forceful eradication of microbial contaminants (Dunn, 2007), an emergent, thoroughly modern, artisanal ethos is committed to working in selective partnership with ambient microscopic organisms to realize human values. These two regimes of hygiene may be termed the Pasteurian and post-Pasteurian (cf. Paxson, 2008, 2013). While Pasteurians view raw-milk cheese as a biohazard, potentially riddled with bad bugs, post-Pasteurians regard it as the inverse: a traditional food processed for safety by the action of good microbes – bacteria, yeast, and molds – that can outcompete bad bugs for nutrients in milk.
For much of the 20th century, the particularity of harmful germs has overdetermined popular views of the microorganismic world as a whole; under the modernist, Pasteurian regime of the FDA, microbes have been figured as perilous, requiring human control (Tomes, 1999). This peril has represented threat not only to human health, but also to market expansion. In addition to pathogens such as E. coli and Listeria, unruly food-borne microbes include bacteriophages that are harmless to human health but thwart proper fermentation and the development of desired odors and flavors in cheese and other fermented foods. Because pasteurization kills virtually all naturally occurring microorganisms, to make cheese pasteurized milk must be re-seeded, generally with laboratory-isolated, freeze-dried strains of lactic acid bacterial cultures (‘starter cultures’), to set in motion the fermentation process. The introduction of pasteurization into cheesemaking in the 1930s in the United States had as much to do with improving consistency, achieving standardization, and creating economies of scale – in other words, with market concerns – as it had to do with health and safety concerns (Latour, 1988; McMurry, 1995: 123–124). Pasteurization enabled the advent of standardized, industrial cheesemaking.
Today, the market is diversifying its concerns. Increasing numbers of consumers are looking for non-standardized, gastronomically distinctive foods whose qualities, they believe, promote health when ingested and whose trade might support family farmers and artisan entrepreneurs, rather than the agro-industrial giants partly responsible for the land’s environmental degradation. They are looking for ‘model’ foods with which they might broker a remediated agriculture. The very quality that gives food-safety officials pause about raw-milk cheese – it is teeming with an uncharacterized diversity of microbial life – makes eating it particularly desirable for some consumers. Some fans of raw-milk cheese argue that human stomachs more easily digest such ‘naturally’ probiotic foods, while others stress its gastronomic qualities (cf. McCalman and Gibbons, 2009). As one cheesemonger blogged, ‘there is a je ne sais quoi about raw milk cheese. The flavors … are more pastoral, have a touch more earthiness to them, more nuance … and [are] just … more vibrant’. 7 Such post-Pasteurian discourse portrays microbial diversity as a good in itself, as naturally overflowing with value to humans.
In countering Pasteurian dreams of microbial control, post-Pasteurian fantasies of microbial flourishing are not indiscriminate in their passion. We refer to raw-milk cheesemaking as post-Pasteurian rather than anti-Pasteurian because it recognizes risk and guards against infection through hygienic practice, while it moves beyond Pasteurianism in recruiting ‘good’ microbes as friends and allies in this effort. Joining a wider movement against the industrial standardization of food, post-Pasteurians say that pasteurization is less an obligatory passage point to cheese safety than a sledgehammer squashing the potential benefits of microbial abundance (see Paxson, 2008).
While this section began by acknowledging that food-borne microbes still represent peril, what we find fascinating about a surge of interest in raw-milk cheese and other fermented foods is that microbes are simultaneously figured as agents of a nature that is not fully objectified and that is acknowledged to be inseparable from human enterprise. For this reason, as we will show, in seeking to rescue indigenous microbial cultures from industrial homogeneity, post-Pasteurian cheesemakers may endeavor to legitimate nonindustrial foodmaking practices. Microbes embody potential not because of their brute materiality, but because they can be enrolled in modeling, and thereby shaping, new food science and politics.
The laboratory life of cheese, or cheese as a model ecosystem
Microbiologist Rachel Dutton, Bauer Fellow in Systems Biology at Harvard University, along with her postdoc, mycologist Ben Wolfe, has begun studying the surface rind of ‘naturally’ aged cheese as a model microbial ecosystem for understanding the behavior of microbes in complex communities. As Dutton (2012) explained the impetus for this work to a group of artisan cheesemakers, traders, and dairy scientists at a conference on ‘The Science of Artisan Cheese’, ‘in the real world’ microbes live in communities and interact with elements of every environment – microbial interactions are important for the ‘health’ of every ecosystem on the planet. However, she added, scientists know very little about microbial communities because they generally study microbes in isolation in laboratories. Dutton has brought aged cheeses into her lab because their rinds constitute densely packed, multispecies microbial communities, making them ‘model microbial ecosystems’. When Paxson visited Dutton’s lab, she saw cheeses represented by Petri dishes containing the cultured strains of bacteria sampled from wheels made by some of the artisans Paxson interviewed during her ethnographic fieldwork (see Figure 1). In studying, through cheese, how communities of bacteria and fungi interact with one another, Dutton and Wolfe were initially engaged in natural science research aimed at learning, as Dutton puts it, ‘who’s out there’. This is more Darwinian exploration than industrial technoscience. Dutton and Wolfe set out to understand what (or who) a cheese rind is, and how it got that way, at the microscopic level; they had little interest in (say) improving its shelf life or market stability.
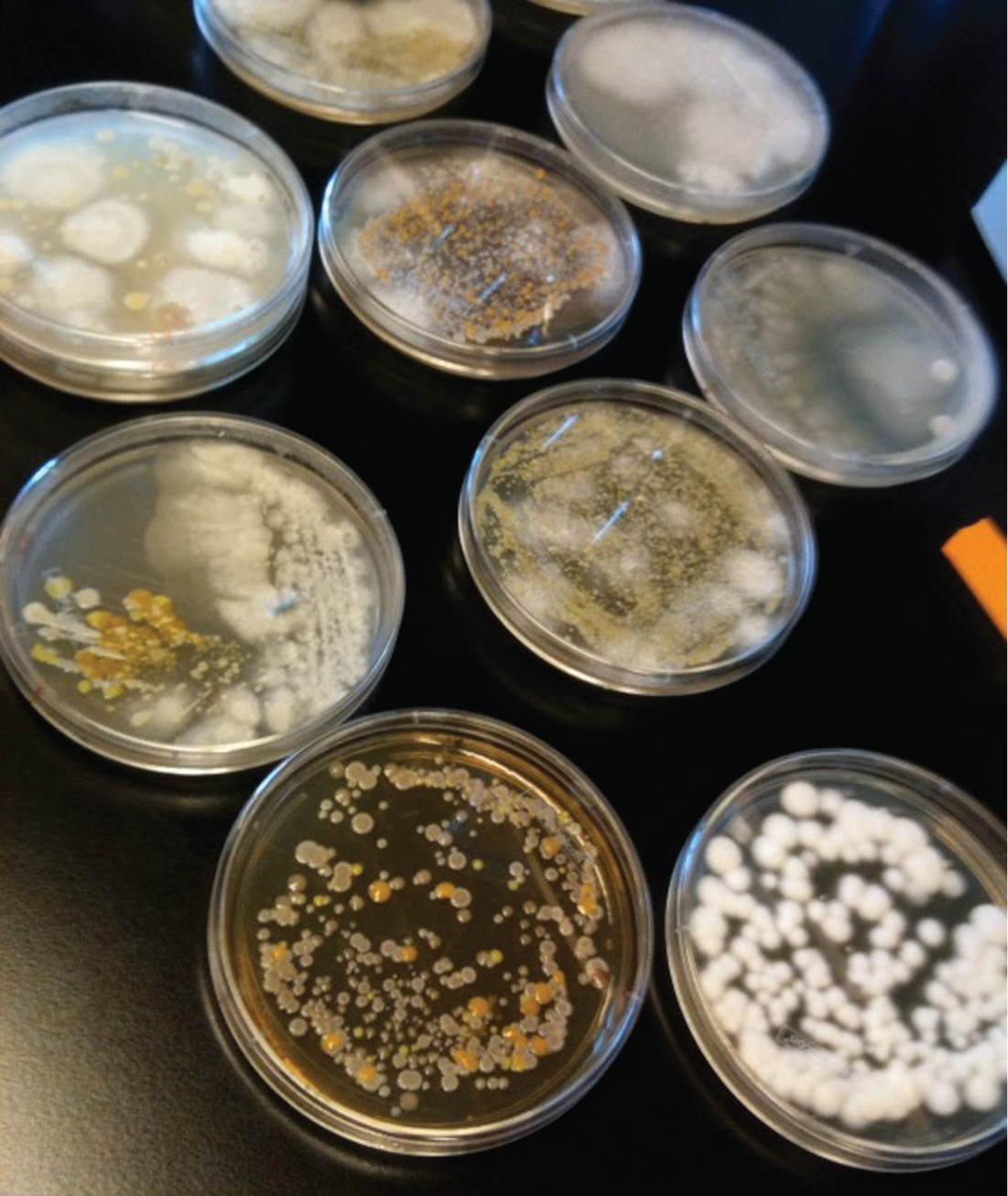
Bacterial cultures sampled from artisanal cheeses, from Dutton’s lab.
When Dutton first approached cheesemakers about participating in her laboratory study, she did so nervously. As a microbiologist, she was accustomed to encountering a public squeamish about microbes, even addicted to hand-sanitizer. Instead, she found artisans curious about ‘who’ she would find on their cheeses. Microbiologists often quip that ‘everything is everywhere’, and Dutton and Wolfe are indeed discovering that cheese rinds contain microbiota associated with an astonishing range of ecosystems and places. Their preliminary microbiological work has received considerable attention in the cheese world. The Spring 2011 issue of Culture: The Word on Cheese magazine featured an 8-page color photo spread of highly magnified, aestheticized images of bacteria and fungi that the lab had sampled from the rinds of well-known cheeses and cultured in Petri dishes (the very samples Paxson saw in the lab). A caption explains, ‘Bacteria and fungi growing together make up a microbial ecosystem, and as the microbes grow they contribute to the flavor, smell, and texture of the ripening cheese’ (Dutton, 2011). How, precisely, microbial abundance contributes to distinctive cheese flavors is a complex question that scientists, primarily in Europe, are just beginning to tease out. Dutton (2011) herself clarifies, ‘Ultimately, I hope to discover what cheese microbes can teach us about the microbial ecosystems found throughout nature’ (p. 81).
But while Dutton’s lab is interested in generalizing from cheese, cheesemakers such as Andy and Mateo Kehler, whose Jasper Hill Farm in Vermont Dutton and Paxson visited together in 2011, are more interested in specificity. Might Dutton’s lab, the Kehlers wonder, turn up a previously uncharacterized microorganism – or a distinctive microbial community – indigenous, even unique, to the ecosystems of their farm and cheese? To be sure, in the identification of microbes the Kehlers spy marketing potential for distinguishing their product in an increasingly crowded niche market. At the same time, however, the perceived value of any such identified microbes lies in their potential to trace a tangible, material connection between their cheese and the place – the environment, the land, the grasses – on which, or from which, it comes into being. The Kehlers ask whether the discovery of unique microbes or distinctive microbial communities might ground claims to artisanal cheese terroir – a French term used by winemakers to refer to a conjuncture of land, environment, climate, and customary practice said to generate distinctive taste (and thence also social distinction) in agricultural products. Could cheese represent a model ecosystem, not just in the scientific sense hoped for by Dutton, but also in the social sense hoped for by the Kehlers?
The notion that there might be a microbial basis to the potential for fermented foods to embody and convey a ‘taste of place’, anthropologist Amy Trubek’s (2008) gloss on terroir, is gaining traction in the cheese world and beyond (the tagline of The Cellars at Jasper Hill e-newsletter is ‘A Taste of Place’). In 2006, a Public Broadcasting Service (PBS) documentary, The Cheese Nun, showcased the microbiological research of Sister Noella Marcellino, whose study of the yeast-like fungus Geotrichum candidum common to certain types of cheese rind (including the Bethlehem cheese she makes at her Abbey in Connecticut) followed from concern that ‘The industry trend toward standardization of inocula and ripening conditions may lead to the loss of empirically derived biodiversity’ (Marcellino et al., 2001: 4753). In the film, Marcellino says, ‘Just as you want to save a certain kind of tree in the rain forest, you want to save the microbes that are part of a region, because they’re the ones that have contributed to the flavor and special unique character of a cheese’. In 2011, Wisconsin’s Dairy Business Innovation Center, a state-funded office that offered technical advice for on-farm, ‘value-added’ dairy processing, sponsored a seminar to explore the potential of terroir for establishing a name-protected designation for cheese fabricated in an area of southern Wisconsin known as the Driftless region, whose topography is marked by having escaped glaciation at the end of the Pleistocene. A French technical consultant, Ivan Larcher, suggested at the seminar that the identity of the scores of types of bacteria, yeasts, and molds that enter milk prior to and during the cheesemaking process
is different on different farms, even if they are only one mile apart. Each farm has its own spectrum … and the dream of every cheesemaker is to develop their own spectrum of bacteria, to make a personal signature. The spectrum is directly related to the place you live, and this is the concept of terroir, to make something different based on where you are. (Tenenbaum, 2011)
One cheesemaker in the Driftless region has done precisely this. A few years previously, Willi Lehner took a walk in the woods behind his house and ‘harvested’ soil microbes; he soaked clumps of his backyard in water to isolate the indigenous microorganisms, strained the water and added it to a brine solution. Smearing wheels of a raw-milk, Havarti-style cheese with this brine, he created Earth Schmier, a toothsome, pungent – earthy – cheese. In the form of Earth Schmier, cheese might be considered a biotechnology for localism – or a model ecosystem expressing felt connection to a place.
As more is discovered about cheese microbiology, however, claims of grounding cheese terroir in the particularity of local microbes run up against microbes’ mobility – or rather, their everything-is-everywhereness. Willi Lehner told Paxson he got the idea for harvesting microbes after visiting Irish cheesemaker Giana Ferguson, famous for inoculating her Gubbeen cheese with laboratory-cultured isolates of a previously uncharacterized bacterium first discovered on her smear-ripened cheese; the microbe is now named Microbacterium gubbeenese. Trace amounts of M. gubbeenese have since turned up in French cheeses of similar style. As more becomes known about the microbial bodies populating artisan cheeses, it seems that ubiquity rather than uniqueness is coming into view – put differently, the figure of the unique microbe is being swamped by the ubiquity of microbial presence. Attempts to characterize microbial uniqueness, singularity, or individuality end up pointing to ecosystemic conditions and connections that blur the boundaries of what people have imagined to be ‘local’ settings. After all, the force selecting for a particular microbial community in the ecosystem of a wheel of cheese is not strictly geographical location scaled to a farm or region (microbes do not obey property lines or political boundaries). Human selective agency is fundamental – for example, in choosing and adapting particular recipes. Based on DNA sequencing, Dutton and Wolfe’s initial research has confirmed Marcellino’s findings, too, that recipe or cheese type is a far greater predictor of microbial communities (of who’s there) than is geographical region or place – that is, the environmental conditions that generate particular microbial communities scale to a wheel of cheese rather than to a geographical or political region. Technique is more determining than, say, climate. This need not preclude the notion of terroir; French winemakers have long argued that the ‘place’ of terroir is not reducible to geography but emerges from conjunctions of environmental conditions and cultural practices carried out in a particular locale over the duration of successive human generations – from putting in place a durable model of human–ecological collaboration (Demossier, 2011; Teil, 2012). But it complicates the picture of ‘place-based’ foods in the United States, where artisan practice is continuously (re)invented not so much as ‘tradition’ but as novel innovation (Paxson, 2010).
American-style artisanship trades on the uniqueness of the form resulting from the impression of artisan’s hands working directly on the material. We suggest that this commitment to the unique signature of hands-on artisanship helps to frame thinking about the microbial signature of a cheese-producing farm or creamery. During a visit to the Cellars at Jasper Hill, where the Kehlers age their own and other area producers’ cheeses, Marcellino – whose own laboratory research found evidence against sui generis microbial uniqueness – scrawled a comment, graffiti-style, on a wall that bespeaks the sentimental pull – and proprietary value – of the unique signature, ‘The world of cheese awaits wonderful cheeses aged in these beautiful caves. I await learning more about the fungi grown here that are uniquely yours!’ Ecosystemic thinking about microbes remains in tension with a focus on identifying individual strains.
More recently, the idea of cultivating a ‘microbial terroir’ that might stamp a fermented food with uniquely located and therefore distinctive ‘signature flavors’ is being championed by the likes of celebrity chef David Chang. In an article in the International Journal of Gastronomy and Food Science, Chang and associates at his Momofuku restaurant’s test kitchen describe developing a fermented pork product, butabushi, as a source of intense umami (savory) flavor to be drawn off and used in cooking (Felder et al., 2012). While clearly excited about the ‘wild fermentation’ they were undertaking in the kitchen (cf. Katz, 2003), these post-Pasteurian commercial chefs were also concerned about the product’s safety for consumption, and so they sent samples of their porcine microbial ecosystem to Dutton’s Harvard lab for testing. Dutton and Wolfe not only screened for potential pathogens (none were found) but also conducted DNA sequencing and analysis to identify ambient fungi that may contribute to the sensory character of the resulting comestible. To such chefs as Chang, microbes would seem to literalize the recent culinary fetish with place-based foods; in a recent profile of Dutton’s lab in the Food & Wine section of the New York Times, Chang describes a desire to inoculate his miso, fish sauce, and fermented tenderloin with the DNA of New York City, asking ‘How can we make New York taste New York? … What makes terroir is the microbes. It’s literally what’s in the air’ (Smith, 2012).
Between artisanal cheese and Momofuku katsuobushi, Dutton’s lab has found itself thinking more in terms of applied microbiology than Dutton anticipated. What is so compelling – to a couple of research microbiologists – about the potential of microbial terroir? In an essay on ‘American microbial terroir’ published in a special 2012 issue of the journal of food writing Lucky Peach (guest-edited on this occasion by David Chang), Ben Wolfe (2012) acknowledges, ‘To a microbiologist, hearing a chef get excited about Aspergillus oryzae and Neurospora sitophila is mind-blowing’ (p. 103). In fermented foods, microbiologists such as Dutton and Wolfe find a vehicle through which they can convey to a lay public their sense of wonder at the microbial world, a world where what matters is not so much who’s out there but instead what microbes do together within certain communities and environments. The materiality that is meaningful in microbiology is not the cellular biology of microscopic organisms, but the relationality of microbial ecosystems in which microbes populate and modify other organic matter, turning dirt into soil, milk into cheese, and meat scraps into salami.
‘Signature flavors’ are not traceable to the identity of any particular microbes so much as they result from the presence of managed microbial diversity. When we eat a cheese or salami, we are not tasting the bodies of bacteria and fungi but rather, scientists such as Dutton would explain, the by-products of microbial metabolism in breaking down the enzymes, fats and carbohydrates in milk or meat to release certain odors and flavors. Microbial ‘signatures’, detectible traces of distinctive microbial presence, may therefore be left not by individual microbes but by particular constellations or communities of microbes – the ‘spectrum’ that Larcher described.
Indeed, the way research scientists such as Dutton understand microbes today – not so much as individuals under a microscope, but as communities in ecologies newly legible through high-throughput gene sequencing – also presses for a more ‘environmental’ sense of them in the wild. That ‘environmental’ sense can sometimes make surprising and sometimes revelatory links (both actual and conceptual) across what might be putatively different ecosystems; as we were doing this research, we were interested to learn that the predominant bacterium in Dutton’s model ecosystem of Winnimere, a brine-washed cheese made by Jasper Hill, was a halomonas, a kind of halophile (a salt-loving organism) most commonly identified with arctic sea ice. At the ‘Science of Artisan Cheese’ conference in 2012, Dutton and Wolfe announced preliminary findings from their microbial survey; one is that halomonas favors other washed-rind cheeses similar in style to Jasper Hill’s Winnimere. Dutton now suspects that halomonas are most likely carried to cheese by the cheesemaker via sea salt, salt being the prime ingredient in the brine with which such cheeses are ‘washed’. This is somewhat counterintuitive to cheesemakers, who view salt as an antimicrobial agent rather than a microbial vector. Viewing the microbes taxonomically as well as ecologically, Winnimere’s halophiles are a match, too, for some found in hydrothermal vent sites, which feature in Stefan Helmreich’s work on microbial oceanography. Such microbes take us far from bucolic farmlands, to the bottom of the ocean and, perhaps, to extraterrestrial worlds.
The case of astrobiology
Alien peril and promise
Astrobiology is the study of life in cosmic and planetary context, with respect to astrophysical and geological processes, whether these unfold on Earth or on other worlds. It originated under a different name, exobiology, founded around 1960 by Nobel Laureate Joshua Lederberg, a Stanford bacterial geneticist concerned that spacecraft returning to Earth might harbor extraterrestrial microbes that could contaminate the planet. Time magazine in 1961, in a profile of Lederberg’s research, warned,
The invaders most to be feared will not be little green Venusians riding in flying saucers or any of the other intelligent monsters imagined by science fictioneers. Less spectacular but more insidious, the invaders may be alien microorganisms riding unnoticed on homebound, earth-built spacecrafts. (quoted in Wolfe, 2002: 194)
Exobiology was inaugurated as the biology of invasive alien species, a defense against alien nature. Although Lederberg was also attentive to the potential contamination of other worlds – and particularly the Moon – by earth biota, public focus, and funding centered on the contamination of Earth by alien life. Historian Audra Wolfe (2002) notes that these concerns were saturated with Cold War imagery: ‘The American duty to protect freedom, through interplanetary settlement if necessary, might be challenged by invisible internal enemies’ (p. 185).
The biological defense of the planet from extraterrestrial contaminants is still a going concern at National Aeronautics and Space Administration (NASA). At a 2005 workshop on astrobiology at the Marine Biological Laboratory in Woods Hole, Massachusetts, we met a scientist who holds the title of Planetary Protection Officer – though he wears the title somewhat ironically, since most of what he oversees is the less-than-glamorous sterilization of spaceships sent to Mars. The grand American frontiering narratives that organized much space research in the 1960s have these days morphed into more modest sentiments of interplanetary ecological stewardship. Notional microbes elsewhere in the universe are now part of a nature, of a diversity, that needs protecting (one might be reminded of the science fiction show Star Trek, and the ‘prime directive’ its spacefarers were to follow, of noninterference with ‘new life and new civilizations’). NASA plans, for example, to send spacecraft to Jupiter’s icy moon, Europa, which astrobiologists consider promising destinations in need of a sort of conservationist care:
Planetary protection requirements for this mission will focus on ensuring that the spacecraft will not inadvertently crash into one of the target moons and thereby risk melting ice and possibly contaminating liquid water. Planetary protection standards recommended by the National Research Council’s Space Studies Board for missions to Europa (Preventing the Forward Contamination of Europa) would limit the probability of contaminating liquid water on Europa with a viable terrestrial organism to less than 10−4 per mission.
8
A post-Cold War environmentalism takes in the entire solar system (cf. Olson, 2010), yet-to-be-discovered microbes included.
With the official founding of the NASA Astrobiology Institute (NAI) in 1998, exobiology had a makeover. Astrobiology came to be characterized as the study of the origin and evolution of life in a cosmic context (see Des Marais et al., 2003). Microbiology came to play a central role, not only because single-celled organisms seemed a more likely extraterrestrial find than multicellular creatures, but also because late-20th-century research on extremophiles, organisms living in exceptionally hot, salty, or pressured environments, had generated new speculation about the limit capacities of living things. Extremophiles turned out to be capable of thriving in such settings as deep-sea hydrothermal vents, underwater volcanoes where temperatures and pressure rise well above the comfort zone of most known organisms. Such organisms came to be described as analogs of early one-celled earthlings, ancestors of all life, and also as possible pointers to what life might look like on remote worlds, perhaps in alien oceans on Jupiter’s moon Europa or on Saturn’s Enceladus.
Much as with discussions about cheese, then, microbes are no longer looked upon here as enemies, but are welcomed as possible companions to contemporary human cultural projects – in this case, the search for extraterrestrial life. Extraterrestrial life, these days, is no longer about the civilized interlocutors once sought by the Search for Extraterrestrial Intelligence, but about the more diminutive microbe, a sign not of extraterrestrial culture, but of extraterrestrial nature.
That nature, increasingly valued rather than feared, is – like the nature of cheese – believed to harbor a surplus of multiplicity, abundance, and potentiality humans have not yet discovered or characterized. That belief motivated a remarkable astrobiological claim in December 2010, when NAI announced that they had reason to believe that microbes might be capable of employing arsenic (in place of phosphorus) as one of the six chemical building blocks of life (Wolfe-Simon et al., 2010). Geomicrobiologist Felisa Wolfe-Simon had isolated a microbe from the sediment of Mono Lake, a body of water in California that is ‘highly alkaline and rich in carbonates, phosphorus, arsenic, and sulfur’, and taken that microbe and fed it in the lab, in a medium that included sugar, trace metals, and vitamins, but no phosphate. In the absence of any material with which to make the usual phosphorus backbone of DNA, her microbes made that backbone, she claimed, using arsenic. As Wolfe-Simon declared, ‘This microbe, if we are correct, has solved the challenge of being alive in a different way’. Wolfe-Simon and her colleagues suggested that the bug might point to possible forms that extraterrestrial life might take.
The microbe, strain GFAJ-1 of the halophilic family Halomonadaceae, became for a brief while a poster child for possible forms as life-as-it could-be. Recall that halophiles were also present on Rachel Dutton’s model cheese – though there leveraged into gastronomical rather than astronomical relevance, into claims about the specifics of human projects of food manufacture rather than into claims about the general nature of life itself. Wolfe-Simon’s arsenic research hoped to leverage the microbially unique into a claim about the biologically universal and it did so with respect to an ecosystem within which a particular kind of microbe might thrive. In the terms of Ankeny and Leonelli (2011), this was intended to become a model organism with a ‘representational scope’ that could explain a wide range of possibilities for the molecular makeup of life itself.
In the summer of 2012, Wolfe-Simon’s claim was roundly contested. Researchers trying to replicate her results found themselves unable to do so (Erb et al., 2012; Reaves et al., 2012). Tobias Erb and collaborators tried to grow GFAJ-1 in a phosphorous-depleted arsenate-containing medium but determined, using mass spectrometry, that such arsenylated compounds as happened to emerge in their samples were not formed biotically by GFAJ-1, but rather came together a priori in the organism’s growth medium. That failure speaks to the possibility that Wolfe-Simon’s lab let their sentiment outrun their science, a desired meaning outrunning the materiality they had before them. What they thought was a specific figure (GFAJ-1) that might highlight and contour a universal ground (life), became a mirage. But this undoing was not the doing of materiality as such. Rather, GFAJ-1 as a model organism had its prescriptive character undone; it collapsed back into being a description of only itself, and a description of what many scientists started to claim was a very unremarkable, even artifactual, microbe. To use a term coined by Nicole Nelson (2013) in her work on how mouse models for human biology are made and unmade in lab practice and rhetoric, the ‘epistemic scaffold’ of GFAJ-1 collapsed under the weight of Wolfe-Simon’s claims for its representativeness. It became legible as embedded within a rather exclusive ecology, one that may not generalize beyond itself and, more, one that may not even have been constituted in the way Wolfe-Simon had claimed.
The laboratory life of life, or simulated alien microbe-scapes as model ecosystems
While it is now generally agreed that Wolfe-Simon radically overstated her claims, the project of situating ‘life’ within broad planetary conditions of possibility endures as the hallmark of astrobiology, and the field continues to rely on the promise of microbes as revelatory entities that might reveal life’s universals with reference to unexpected particulars. We might think of them as ‘bio-objects’, what Vermeulen et al. (2012) describe as technical forms that acquire significance for biological work when they are framed as standing metonymically for vitality. For example, aborted fetal tissue may be treated as ‘waste matter and dead’, and hence not a bio-object, ‘yet can be re-vitalized as source material for stem cell lines’, becoming a bio-object (Vermeulen et al., 2012: 2; compare Franklin, 2006 on the ‘IVF-Stem cell Interface’). Microbial ‘bio-objects’ are almost always imagined with respect to an environment, that is, in the idiom of the model ecosystem.
Sometime after the refutation of Wolfe-Simon’s claims in Science in July 2012, we met with Massachusetts Institute of Technology (MIT) geobiologist Tanja Bosak (an early skeptic of Wolfe-Simon’s arsenic story), whose laboratory is affiliated with the NAI, a confederation of about 14 research teams working across many academic institutions, research labs, and NASA centers. Working as part of MIT’s ‘Advent of Complex Life’ team, Bosak is an Earth scientist interested in understanding how our planet came to have the geological and biochemical properties it now hosts. She wants to know, for example, when and how Earth’s atmosphere filled with oxygen. The standard story is that oxygenation unfolded some half a billion years ago – though since the responsible cyanobacteria existed as early as 2.5 billion years ago, the question raised is why it took so long. Finding out about the microbial history of Earth’s oxygen atmosphere turns out to be challenging because microbes do not fossilize; the most they do is leave bubble-like absences in rocks, traces of their once-upon-a-time metabolic activity. Much of Bosak’s work consists of trying to create model ecosystems in the lab that can produce such forms. She has to operate, she told us – and similarly to Rachel Dutton – as something of an experimental microbiologist, culturing collections of microbes with the aim of ‘translating their behaviors and metabolisms into shapes’, outlines that may be matches for billion-year-old marks left in Earth’s paleontological record. These outlines might be possible ‘biosignatures’, defined in the astrobiological literature as ‘any measurable property of a planetary object, its atmosphere, its oceans, its geologic formations, or its samples that suggests that life was or is present’. A short definition is a ‘fingerprint of life’ (McKay et al., 2002: 625).
Bosak showed us some of the work behind her search for possible biosignatures, escorting us to a refrigerator in her lab that held tens of microbial cultures, floating in large Petri dishes. She showed us a variety of cyanobacterial microbe-scapes, some of which featured submarine mini-gardens of cone-like shapes, possible model ecosystems that might shed light on the question of how ancient cyanobacteria left conical impressions in ancient rocks (see Figure 2). Think of these simulated alien ecologies as modeling the ahuman terroir of an early, just-about-to-be-oxygenated Earth.

A cyanobacterial microbe-scape from the Bosak lab.
While Bosak’s work focuses on the evolution of the Earth system, her affiliation with MIT/NASA’s ‘Advent of Complex Life’ team bespeaks the wider interest that this work may have for biology writ large, both as an object of study and as a discipline. In these studies, microbial life is valuable as a way of developing intuitions for finding signs of life more generally, for recording biosignatures, whether in fossilized impressions of microbial formations, or in measuring concentrations of sulfur that reflect microbial metabolic activity (compare Helmreich, 2006). ‘You’ll never be able to find DNA’ from early Earthly organisms, Bosak told us, ‘but you’ll be able to trace what they did’. Note the complex place of ‘materiality’ in this project: a biosignature is not a concretization of a fully present and characterized materiality, but is rather a sign of a possible chemical or biochemical world – an ‘absent presence’, to take a concept from Jacques Derrida (1982), who suggests that taking signatures as traces of authentic presence paradoxically relies upon the absence of the agent that produced the signature.
While Bosak is cautious about how much one might generalize to other possible planetary histories from microbial models (she prefers to calibrate her model system data to existing evidence in Earth’s geological history), other scientists in astrobiology move more quickly to extraterrestrial speculation – to thinking about suggestive absences (grounds) as promising pointers to possible presences (figures). The journal Astrobiology features many articles postulating possible analogs between Earthly environments and extraterrestrial ones, and microbial life is almost always in the mix (see Popa et al., 2012). Take a few examples: in ‘Life at the wedge: The activity and diversity of Arctic ice wedge microbial communities’, microbiologist Roland C. Wilhelm et al. (2012) write that ‘recurring detection of terrestrial microorganisms in subsurface ice environments, such as ice wedges found beneath tundra polygon features’ suggests the possibility that ‘polygonal terrain on Mars … may be, or may have been, a suitable habitat for extant life’ (p. 347). And in ‘Biosignature detection at an Arctic analog to Europa’, Damhnait F. Gleeson et al. (2012: 135) discuss looking for ‘potential indications of the presence of life’ in the oceans of Jupiter’s moon, Europa, by working at the ‘characterization of a range of biosignatures within potentially analogous [to Europa] sulfur deposits from the surface of an Arctic glacier at Borup Fiord Pass’. Arctic microbes on Earth stand in as potentially akin to European ones (it is not incidental that the word ‘potential’ appears in 635 articles out of the 770 that Astrobiology has published).
This ongoing discussion of the implications of Earthly microbial research for extraterrestrial worlds, Bosak suggested, emerges in part because many researchers frame their research questions and agendas to align with NASA space mission plans, which demand that knowledge of biology on Earth be employed as a particular case that may have more wide-ranging, even universal, implications. Consider the Mars Rover Curiosity, which landed on Mars on 6 August 2012, touching down in a ‘crater [that was] covered with water in the past, and that … contains minerals that can only be formed in water as well as clays that also require an H20 environment’. The Rover carries a mobile laboratory ‘assembled to look for the carbon-based building blocks of Martian life and to explore the possible habitats where life might once have existed’. 9 The properties of ‘life’ sought by NASA are those that characterize microbial vitality.
In Laboratory Life, Latour and Woolgar (1986 [1979]) concluded that scientists occupied themselves with generating inscriptions, registrations of data that might become, if defended by a robust enough sociology, facts. In the astrobiological work we have described here, microbial ‘signatures’ operate as a special kind of inscription – one that has built into it a particular set of claims about and models for ‘life’, and about life’s capacity to leave traces of itself. Such an inscription means to be a description, but it is also, at least in part, a prescription – a pointer to what one should find. A signature becomes a kind of ‘bio-object’, a formal pointer to vitality. The laboratory life of ‘life’ comes to depend greatly on how inscriptions – like Bosak’s bubble-like absences in rocks, which she seeks to investigate using model ecosystems – are interpreted, as descriptions for how we might, prescriptively, think of life.
The promising model of the microbe
In both artisanal cheesemaking and astrobiology, practitioners of microbial ecology (in cheesemaking, these are both scientific and lay practitioners) appeal to microbes to anchor their cultural projects, whether these are to valorize and stabilize local and artisanal foodmaking or to extend the possible reach of biological speculation about life elsewhere. To be sure, that deployment of microbial life – as promise rather than peril – is in part a function of a broader environmentalist vision. 10 No longer are we in a modernist Cold War world in which enemies are everywhere, even unto the microbial realm; that was the ambient ethos surrounding both the 1949 supremacy of pasteurized American cheese and the 1960 enunciation of exobiology as the defense of Earth against microbes from outer space. Rather, microbes have now become models for apprehending biotic nature, both in the sense of serving as representatives of dynamics that suffuse the organic world in general, but also in the sense of standing as promising tokens for reimagining nature as it could or should be. It is worth pointing out that this optimistic vision of microbes has been enabled precisely by the fact that the Pasteurian project has been so successful; microbes can be promising for those people who no longer have to worry about smallpox, polio, cholera, and other agents of infectious disease.
We elaborate, in conclusion, on the concept of the model ecosystem, highlighting once again the fact that such models are often not only descriptive, but also at the same time prescriptive. In itself, this is not news: ecological models have since their inception been heavily laden with moral prescription, especially since many such models, especially in the 20th century, were made with management in mind (see Anker, 2002; Toogood, 2008). As Lorraine Daston and Fernando Vidal observe in The Moral Authority of Nature (2003), nonhuman animals (think of the industrious bee or beaver) have often been held up as models for human virtue. The US setting of our cases is also important. Moral appeals to nature have a storied history in the United States, a country that historian Perry Miller (1967) called ‘nature’s nation’. As environmental historians have shown, this national narrative has inflected everything from the founding of national parks to environmental activism (Bak and Hölbling, 2003). While microbial science certainly has many transnational matrices, the location of American artisanal cheese and the science of the NASA in a post-pastoral and post–Cold War era makes the microbial story we have told here part of an American ideological and institutional history, one that could be elaborated to speak to how microbial ecosystems bump up against food regulation regimes and public health projects, national frameworks for funding, anxieties about national biologies (note, for example, that microbial nature [so far] has not been scripted into the fraught biopolitics of American race thinking). That history tells on how microbes are valued, and on how they are worked upon and by whom. 11
How shall we think about the human hand in model ecosystems? We find it instructive to think about that agent through the language of the ‘signature’ that appears in both artisanal cheese worlds and astrobiological discourse.
For some American cheesemakers, behind the dream of microbial terroir lies the hope that in the absence of valorized, generations-old artisan traditions of cultural practice (as in Europe and the United Kingdom), unique microbial diversity understood to emerge from new configurations of place and practice might give warrant to nonindustrial ways of farming and foodmaking. As Ivan Larcher put it in addressing the Wisconsin cheese conference, ‘the dream of every cheesemaker is to develop their own spectrum of bacteria, to make a personal signature. The spectrum is directly related to the place you live, and this is the concept of terroir’ (emphasis ours). Here, the signature, etched by microbes, emerges at the conjuncture of labor, environment, and terrain. The universal microbial world might yield up the unique in interaction with human practice. Although producers also draw an analogy between their own labor practices and such naturally generative forces as decomposition, which has the effect of naturalizing craft production methods, making them seem, as part of ‘nature’, not only legitimate but also moral, in the normalizing sense of, ‘that’s how food should be made’. Microbially based terroir might seem particularly compelling because microorganisms, invisible to the naked eye, would seem, in comparison to dairy animals’ breed or fodder, to be the wildest, and hence most autochthonous, element of a cheese’s ecology of production. But at the same time, distinctive communities of microbes emerge to form the rinds of cheeses in no small part owing to careful human cultivation – hence the ‘signature’, canonically a mark of the hand of the individual artisan, as a ready-made image for cheesemakers. In constituting the particular materiality of a cheese, nature and culture are fully implicated in one another; neither may be said to ground the other.
For astrobiologists, the dream of finding life elsewhere in the universe is animated by the hope that finding unique microbial diversity can model new ways of being an organism – of being alive – that can in turn amplify what counts as life more universally. As the NASA Astrobiology Roadmap has it, ‘Catalogs of biosignatures must be developed that reflect fundamental and universal characteristics of life, and are thus not restricted solely to those attributes that represent local solutions to the challenges of survival’ (Des Marais et al., 2003). Here, the signature emerges from the discovery of the ubiquitous in the particular, when the collection of microbial diversity yields up the universal.
Signatures, of course, often safeguard promises – and, as Michael Fortun (2008) suggests, promises tend to generate ideas about the future that, in their enunciation, have an uncanny way of folding back into and (re)orienting the present. Signatures also depend on a shared moral economy of meaning. And, indeed, a key feature of model organisms – and one that we wish to lean upon as we press the claim that microbes are becoming models in prescriptive as well as descriptive ways – is that they are often shared, across laboratories and research centers. 12 Microbes in cheese, or rather, the microbial activities that constitute cheese, underwrite shared families of form and flavor (e.g. washed-rind, bloomy-rind, and mold-ripened). Indeed, the microbiological research of Marcellino as well as Dutton and Wolfe suggests that recipe (and hence technique) is the strongest predictor of ‘who’ will end up on the rind of a given cheese – which explains why Microbacterium gubeneese has been found on French cheeses of a similar style. Microbes in astrobiology, meanwhile, are looked to as pointers to shared features of living things. The microbial realm, shared across scales and contexts, variously and simultaneously universal, ubiquitous and unique, has become a fresh court of appeal for those who would model new modes of living with and within biological nature. The question is not simply ‘what is life?’ but rather, ‘what forms of life do we wish to insist upon?’
This question speaks not only to what cheesemakers and astrobiologists are up to, but also, interestingly, to some of the new materialists we seek to critique. The putatively ahuman, nonhuman, inhuman, alien phenomenology forwarded in these discussions frequently comes with a moral aim in mind. Jane Bennett (2010) is explicit. She writes, ‘my hunch is that the image of dead or thoroughly instrumentalized matter feeds human hubris and our earth destroying fantasies of conquest and consumption’ (p. ix). ‘Thing-power materialism’, she writes in an earlier mission statement, ‘emphasizes the closeness, the intimacy, of humans and nonhumans. And it is here, in a heightened sense of that mutual implication, that thing-power materialism can contribute to an ecological ethos’ (Bennett, 2004: 365). Like cheese fans and astrobiologists, Bennett seeks to model a desirable ecological set of relations. Indeed, Vibrant Matter follows and adapts a tradition that reaches back to the 19th-century claims that ‘self-moving’ matter could provide a warrant for a democratic view of the world. Historians of biology Desmond and Moore (1991: 223), in their biography of Charles Darwin, argue that the ‘self-moving’ matter and ‘living atoms’ postulated by such figures as Darwin’s contemporary, botanist Robert Brown, ‘gave a scientific basis to … belief in free men controlling their own destinies … It provided the perfect political analogy – power from below, “mandating” upwards, rising from “social atoms” – the people’. 13 While the new materialism may have jettisoned the explicitly ‘social’, opting instead for the ‘ecological’, it is similarly suffused with a desire for new kinds of model ecosystems, or, to shift registers, for model ecologies, where an ecology is simultaneously a theoretical account of organic relations. Model ecologies are contemporary tools that scientists and their interlocutors (including us, as authors) use to describe desires for exemplary ways of studying human entanglements with nonhuman agencies.
As a final note, we turn to an image on the side of the Jasper Hill Farm barn (see Figure 3), a painting of the Moon made of cheese. It recalls to us A Grand Day Out, a short 1989 animated science fiction film, in which two claymation characters journey in a homemade rocket to the Moon, hoping to find cheese for a picnic they are planning. Arriving on the lunar surface, Wallace, an eccentric inventor and established tyrophile (cheese-lover), and his long-suffering and resourceful dog, Gromit, scout for cheese. Wallace samples the Moon with a cheese knife and exclaims to Gromit, ‘It’s like no cheese I’ve ever tasted’. The joke, of course, is that Wallace is taking as material truth the old European canard that as Wallace repeats it in the film, ‘the moon is made of cheese’. Wallace is, on the one hand, something like the cheesemakers and astrobiologists we have discussed, searching for new gastronomic and astronomic possibility. On the other hand, he is like some new materialists, who, in their eagerness to take on board the latest wisdom from physical science, risk taking a culturally and historically specific account as literal truth. Tellingly, however, the original fable that has it that ‘the moon is made of greene cheese’, recorded as early as 1546 by John Heywood, puts forward a slightly different epistemological tale. In the fable, a clever fox persuades a credulous wolf that a reflection of the Moon in water is a round of cheese. Taking this as an allegory for our subject, science does not produce simple reflections of the world, nor, indeed, distortions of the world, but rather interpretations or models, sometimes suffused with our desires – such as those that many of us harbor for heavenly cheese and cosmic company.

A mural, painted by Tara Goreau, on the barn of Jasper Hill Farm.
Footnotes
Acknowledgements
We thank the scientists and cheesemakers who participated in our research. Heather Paxson thanks Rachel Dutton, Ben Wolfe, Andy Kehler, and Mateo Kehler. Stefan Helmreich extends particular thanks to Tanja Bosak. This article originated as a joint lecture for the Technoscience Salon at the Jackman Humanities Institute, University of Toronto, Ontario, Canada. We thank Natasha Myers and Michelle Murphy for inviting us. From this venue, we developed the talk for presentation at the Institute for Advanced Study, University of Minnesota. Versions of this article were presented at the Department of Anthropology at City University of New York, Graduate Center; in the Program in Science, Technology and Society, University of Michigan; as a Klopsteg Lecture in the Science in Human Culture Program, Northwestern University; as a working paper for the Political Ecology Working Group at Harvard University; and as a lecture at Dipartimento di Scienze Umane e Sociali, University of Bergamo, Italy. We also thank Cristina Grasseni, Grace Kim, Nicole Labruto, Hannah Landecker, Caterina Scaramelli, and Karen-Sue Taussig for useful commentary on draft versions of this argument. Our thanks go to two anonymous reviewers for SSS and to editor Sergio Sismondo for his above-and-beyond work in shepherding this article into its final form.
Funding
This paper research that extends work initially supported, in part, by the Wenner-Gren Foundation for Anthropological Research, grants #6993 (Helmreich) and #7641 (Paxson).