
Abstract
This article foregrounds comparison as a key practice in science by discussing the case of chronological comparability in paleoclimatology. Based on an ethnographic study of a paleoclimate research project, I illustrate how paleoclimatologists are able to produce comparative data on and images of past climates through the use of ‘proxies’. I focus on the calibration of a type of algae as a proxy for climate variables. Such comparability is one illustration of the myriad ways in which relatively standardized forms of comparison underlie conceptions of ‘climate change’ and of ‘climate’ itself. The work of comparison discussed here has relevance for a variety of practices of qualification, quantification, monitoring, and evaluation.
Climate change as matter of comparison
Any assessment of ‘climate change’ involves an aspect of comparison, without which no ‘change’ can be observed. But how is such comparison done? This article foregrounds practices of comparison and, specifically, of comparability as pivotal in science by discussing the case of chronological comparability in paleoclimatology. Climate change is obviously a major preoccupation in contemporary climate science, yet until now its comparative aspect has not been a significant focus of attention (one analysis of the role of comparison on a related issue is Sundberg, 2011). Comparison plays various roles in meteorological observation, in climate modeling, and in the drafting of Intergovernmental Panel on Climate Change (IPCC) assessments and scenarios, all of which are based on forms of interoperability. Comparison plays an important role in the harmonization and standardization of climate monitoring itself, giving rise, for instance, to forms of ‘climate model intercomparison’ (e.g. Lamarque et al., 2013).
Building on studies of standardization and on laboratory studies, this article highlights the practice of comparability by illustrating how paleoclimatologists are able to produce data on and images of past climates through the use of ‘proxies’. Proxy measurement involves the use of proxies as stand-ins for the climate variables of the geological past, and in doing so it relies heavily on comparability: it involves comparability of a proxy and that for which it acts as stand-in, and on the basis of this, it involves the comparability of objects called ‘climates’, which existed along a chronological time scale of millions of years. On the basis of an ethnographic study of a paleoclimate research project, I illustrate how comparability is achieved through what is called ‘proxy calibration’. Specifically, I focus on the calibration of a type of algae as a proxy for climate variables. Such comparability can be considered as one illustration of the myriad ways in which relatively standardized forms of comparison underlie conceptions of ‘climate change’ and of ‘climate’ itself, which is defined as a comparative average across at least 30 years (Barry and Hall-McKim, 2014: 2; Mitchell et al., 1966). Although my case is drawn from paleoclimatology, the work of establishing comparability discussed here has relevance for a variety of practices of qualification, quantification, monitoring, and evaluation. 1
Paleoclimatology has particular relevance for the study of comparison and comparability in that it foregrounds chronological comparison. It not only compares climate variables across time scales of different sizes usually larger than a human lifespan (Bradley, 1999: 32; Ruddiman, 2008: 10–16; Saltzman, 2002: 3) but also has relevance for assessments of contemporary climate, for instance, by providing arguments for the ‘tuning’ and parameterization of climate models. The case discussed here concerns a multi-layered comparison. It involves the comparison of carbon concentrations across different time scales and in different places on earth. This is done by developing a ‘proxy’ for these concentrations, which is a type of algae (dinoflagellates) that takes in CO2 from the sea to produce organic carbon. Comparing the stable carbon isotopic composition (δ13C) found in organic fossil remains of algae from different times enables paleoclimatologists to compare carbon cycling and, therefore, (aspects of) climate and climate change. Isotopes are chemical elements with varying mass numbers; that is, they are chemical elements occurring in lighter and heavier forms. CO2 occurs, among others, as 12C and as 13C. The dinoflagellate algae studied by paleoclimatologists take in CO2 from the sea. The type of CO2 they take in (their carbon composition; that is, the relative presence of 12C and 13C) is thought to reflect the carbon composition of the ocean and the atmosphere, which is heavily correlated with temperature. In this way, algae fossils become proxies for crucial climate variables. Analyzing their carbon composition eventually enables analyzing climates, and thus, such proxies are also called ‘paleothermometers’. But in order for the comparisons to become possible, scientists need to study the ways in which the algae take in CO2 from the sea, in order to learn in what way the algae fossils actually ‘mirror’ the carbon composition of the sea. This requires work on ‘proxy calibration’ that involves biological experiments in which carbon intake of certain algae under various controlled conditions is compared.
Conceptualizing comparison and comparability
Comparison enacts the idea that reality is not wholly made of or actualized in singularities, by which I mean singular, non-recurring, self-organized entities that constitute continuous and complete novelty. Events that are wholly unique share no basis for meaningful – that is, informative – comparison, and in a domain made up entirely of such events comparative systems would be redundant, as would memory systems. At the same time, comparison entails that there is no complete lack of novelty or, less strongly put, of change. Comparison would not be of much interest if objects were compared that constituted a series issuing from a single model. Comparison, rather, relies on combinations of sameness, or similarity, and difference. The question this article deals with is how things get to be rendered comparable. That is, how are objects of science – in this case a proxy that stands in for past climates – stabilized in such a way that they exhibit simultaneously meaningful similarities and differences?
Practices of comparability center on the establishment of what has been called the tertium comparationis, the common element that allows similarity and difference to be established in the first place. This common element is often discussed in terms of commensurability, which implies a degree of similarity of, for instance, object type, scale, and standard of comparison (Espeland and Stevens, 1998; Owen-Smith, 2005). Difference in comparison refers, among other things, to measured or otherwise observed object properties and quantities. In line with prevailing definitions as well as etymology (the middle of the term derives from ‘measurement’), Espeland and Stevens (1998) consider commensurability exclusively in contexts of quantification. For this reason, commensurability is not quite what is meant by the tertium comparationis. In fact, judgments of commensurability and incommensurability are always already comparative statements, based on comparative practices. This becomes clear when a broader concept of commensurability is deployed, such as in Kuhn’s (1996) philosophy of science or in Povinelli’s (2001) account of cultures. Incommensurable paradigms in Kuhn, for instance, are nonetheless comparable and can be judged as incommensurable precisely because they are compared. Likewise, when commensurability does include the possibility of measurement, it follows from an already comparative evaluation. Such practices of comparative evaluation require first an act of positioning, which establishes a parallel between things that, once thus positioned, appear as comparata. That positioning entails a situation of what I shall call comparity.
I take comparity, rather than commensurability, to denote the shared characteristic of occurring in a single scale-defined space of comparison. As such, comparity is what allows comparison to be abstractly qualified; it specifies the quality of what is being compared. Difference, or differential occurrence within a scaled space of comparison, involves a more concrete qualification – for instance, through quantification and the establishment of commensurability – of the relation between the entities compared. Thus, I consider commensurability only as a special case of comparity, one which is merely concerned with the establishment of quantitative comparison and which always already relies on a previous establishment of comparity. Differences only appear once objects or events have been first ‘lumped’ together in an object space (Zerubavel, 1996) and have been ‘frozen’ in formats allowing coordination and, hence, comparison (Bowker and Star, 1999: 135). In other words, differences emerge once a practice of rendering objects comparable as ‘objects’, and hence to some degree of rendering them similar, has occurred. Creating and sustaining comparity, like creating commensurability, is a social process (Espeland and Stevens, 1998) or a practice (Povinelli, 2001). It brings disparate things together as ‘objects for comparative scrutiny’ by stabilizing a space in which these objects, along with potential others, can occur alongside each other in the first place. Depending on their type of specification or their level of measurement, difference in comparison may then involve a quantification of the relation between the entities compared. Often, this involves quantification on the basis of some common metric (Espeland and Stevens, 1998), but it need not. More fundamental, I want to argue, is the creation of an ontological object space, a comparity space, in which an initial qualification is at stake: the qualification of the tertium comparationis or shared basis of comparison. Finally, difference is not yet meaningful if there is not some significance attached to it. This is a qualification that may itself be performed through quantification, as in statistical significance, but, analogous to Bateson’s definition of information, it is crucial in constituting ‘a difference that makes a difference’. Comparison, then, does not involve scales that exist prior to practices of comparison. Rather, it involves a practice of forging what Marilyn Strathern (2004) has called ‘partial connections’ (pp. 51–55) and in which some selection of elements provides a relation between similarity and difference specified as significant for purposes at hand. ‘Doing’ similarity and difference involves a back-and-forth between objects of comparison that involves a constant form of what in Actor–Network Theory has been called translation. The point of calling attention to translation in the most general sense of the concept is to note that the ‘objects of comparison’ do not pre-exist in comparative practices – although they are not ‘socially constructed’ and they can be considered as actively participating in comparison (Stengers, 2011: 51).
Standardization is subject to an everyday work, albeit a work that is most often ‘invisible’ (Lampland and Star, 2009). In a study of ‘intercomparison projects’ of astrophysicists, meteorologists, and oceanographers, Mikaela Sundberg (2011) finds that a main effect of such projects is the standardization of research problems. That comparison is ‘work’, something ‘done’ in practice and requiring skill and connectivity to or enrollment of various actors, is also given in the etymology of the verb ‘to compare’. From Latin comparare, it entails ‘making equal’, ‘bringing together’, and ‘coupling together in pairs’. This ‘making equal’, or what can be called comparity work, precedes difference since it provides the basis for differentiation. It involves an active equalization or bringing together in a shared domain or object space. As M’Charek (2008) writes in an analysis of DNA comparisons, ‘comparability is not inherent in the entities being compared. Comparability is rather a technologically produced ordering of the objects of interest’ (p. 389).
In the case study I present here, the objects of interest are historical, but they have a contemporary reference that is also political. These objects of interest are climates of the past, which are used in comparison in order to qualify statements about contemporary climate change. Such political connotations of establishing comparability are, however, distant and not immediately apparent in the many practices that ensure comparability. In what I propose to call modular comparison, this distancing occurs by establishing and finding acceptance for a particular relation between a proxy – a type of algae – and CO2 levels. That relation then becomes a mobile module that can be shifted along time scales so as to secure climate comparability.
Studying comparison in paleoclimatology
As one textbook states, paleoclimatology uses ‘a wide range of techniques to extract, reconstruct, and interpret the history of Earth’s climate. Much of this history is recorded in four archives: sediments, ice, corals, and trees’ (Ruddiman, 2008: 17; see also Bradley, 1999: 6–7; Cronin, 2010: 28). Especially in the analysis of sediments, corals, and trees, paleoclimatology works with fossils. And fossils, when considered in their practical use by paleo-scientists, are comparative devices par excellence. Michel Foucault (1970) signals as much when he says, ‘The fossil is what permits resemblances to subsist throughout all the deviations traversed by nature’ (p. 156). Speaking of the classical age, in which natural history could be established largely on the basis of a treatment of fossils as containing comparative records, he highlights the basic elements of a practical, comparative use of fossils: similarity coinciding with variation on a temporal scale. This, I seek to show, is the key feature of the fossils used by the team of paleoclimatologists working where I conducted my fieldwork.
Chronology itself specifies a scale along which comparison becomes possible as differentiation within the same-scaled space. Chronology, however, is not a given feature of the world (Halbwachs, 1997; Kwinter, 2001; Schivelbusch, 1986). As Judith Treas (2009) has illustrated, chronological age in humans has become a ‘biographical necessity’ only in the 20th century and was not a common practice prior to it. In science as well, chronology, like any scale (Marston, 2000), is a product of standardization. The International Geologic Time Scale (subdivided in eon, era, period, epoch, and age), standardized by the International Commission on Stratigraphy (ICS), is the most prominent example here, and it is not a stable scale but instead is subject to deliberation and change. Such standardizations therefore also underpin comparability, but for now, I shall accept them as given and focus on the more fine-grained everyday production of comparability.
Crucial in scrutinizing comparison is the question of ‘where’ it is done (Mol, 2002: 218). This article is based on ethnographic fieldwork I carried out through weekly visits, for over a year in 2013 and 2014, to a Dutch paleoclimate research lab, which included a visit to a German lab with which the team I researched collaborates in the calibration of a particular ‘paleoproxy’. The lab is highly successful, measured by the dissemination of its published research, with regular contributions to journals such as Nature, Science, Nature Geoscience, Geology, and Paleoceanography. It specializes in the analysis of fossil algae sampled from sediment cores drilled as part of the Ocean Drilling Program (ODP, 1985–2003) and the Integrated Ocean Drilling Program (IODP, 2003–). The fieldwork involved observations of work practices by and interactions between paleoclimatologists. I also engaged in conversations with researchers while they were working. I jotted these down and recorded them in my field notes as brief interviews, although they were not formal interviews detached from work contexts. In addition, some conversations were extended by email conversations. During fieldwork observations, scientists would often explain what they were doing to me or would include me in their conversations. As fieldwork progressed, there was less need for them to do so, although sometimes I would ask questions, which gave observations the character of brief interviews – something described in other ethnographies of science (Garforth, 2012). Field notes of observations and conversations were made on site and were later extended in reports.
Calibrating a proxy, crafting comparability
To explain how comparability was achieved in the proxy calibration project I studied, it is necessary first to explain the project in some detail. I will focus on the general project aims and then zoom in on the experimental setup, in which stable isotopes will appear as comparability devices. Then, under the heading of ‘comparability techniques’, I identify various steps in the construction of comparability.
The aims of comparison: General research outline of a proxy calibration study
Carbon cycle reconstruction occurs by means of proxy analysis. A proxy, in the words of one of my respondents, is ‘a way to measure something via something else’. Or, as another respondent in the same conversation specified,
It rather concerns an estimation. A way to estimate something you cannot observe directly. It comes from the English word ‘approximation’. You make an assumption and another assumption and another assumption, and then you use a certain value to estimate the real value you wish to know.
Interestingly, the word ‘proxy’ does not come from ‘approximation’, but from procurare, which carries connotations of ‘caring’ and ‘managing’. While the above quote makes clear that, for this researcher, a proxy can be regarded as a form of translation of (a) value, it also involves a particular kind of care for, and management of, matter, which indicates the comparity work this researcher takes for granted. In this case, the proxy concerned is the ‘cyst’ of a particular type of algae, called dinoflagellates. Dinoflagellates are single-cell algae that, after reproducing, form a kind of organic plastic called a cyst, in which they reside during the dormant or ‘hypnozygotic’ phase in their lifecycle (Evitt, 1985; Pfiester, 1984) (see Figure 1).
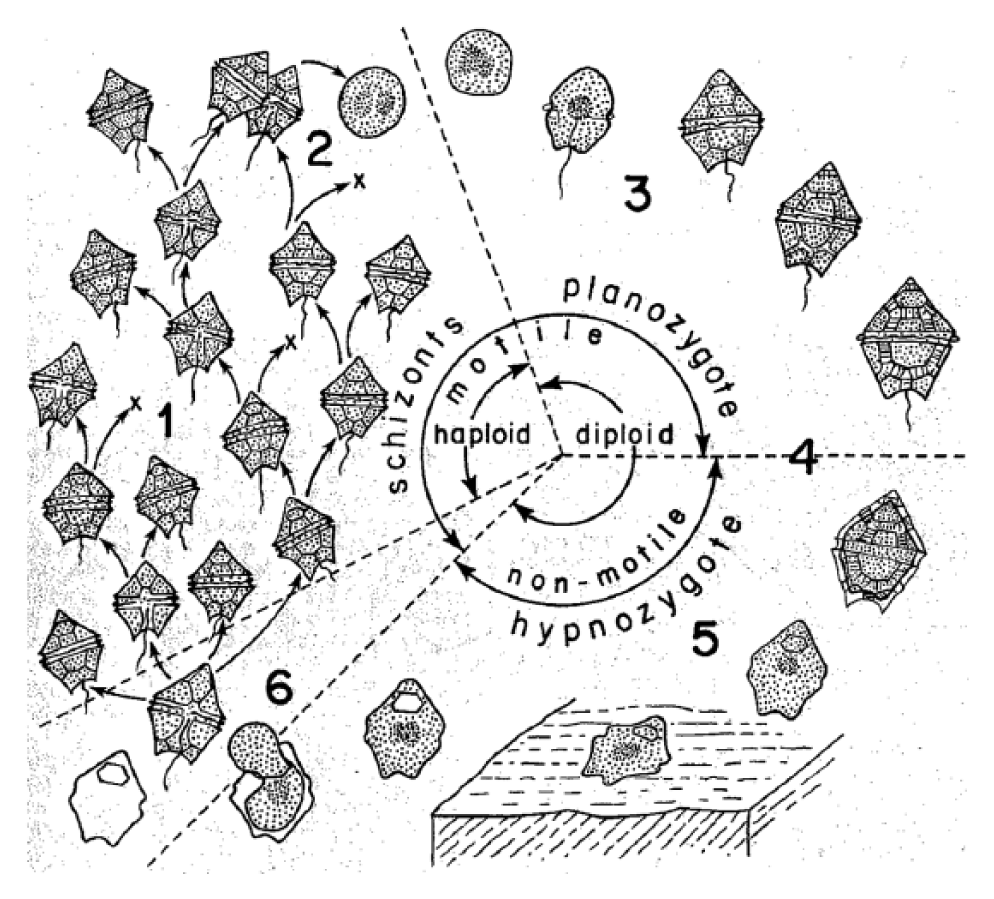
Life cycle of cyst-producing dinoflagellates.
Organic-walled ‘dinocysts’ are made of dinosporin, a type of biopolymer. After a variable amount of time, the algae break through the dinocyst wall and start a new reproduction cycle. Although the algae themselves are not preserved as fossils in ocean sediments, their cysts are, so the paleoclimate researchers look at the carbon composition of these hulls. Since organic-walled dinocysts are produced in part by taking in CO2 from the ocean, which they use for photosynthesis, they are assumed to preserve a record of the prevailing type of CO2 of the seawater in which they form. That means the isotopic composition of CO2 found in dinocysts might be taken as a way of indirectly measuring the isotopic composition of sea surface CO2, which bears a specific relation to sea surface temperature and is considered a crucial climate variable. Thus, the research project I studied was concerned with turning dinocysts into proxies for the carbon cycle during past eras and, via that route, eventually of past climates. As one of the researchers said, ‘Ultimately, the really holy grail of everyone working here, is CO2’. Proxy construction, then, has to do with finding the most reliable ways of reconstructing past oceanic and atmospheric carbon levels for the longest possible time. One needs to know the relationship between a dinocyst and sea surface carbon in order to draw conclusions about past sea surface carbon rates based on carbon found in dinocysts. But because of the small size of the research team (consisting of four researchers, with a dozen collaborators), calibration issues with consequences for carbon cycle reconstruction could be responded to immediately.
A first part of the ‘calibration’ of this particular type of proxy for CO2 reconstruction consists of a set of ‘biological culture experiments’ on actual, currently living dinoflagellates. This project, coordinated in the Netherlands but carried out in the German lab I visited, is aimed at assessing how dinoflagellates – and, therefore, the cysts they produce – acquire CO2.
2
It is therefore most fundamentally concerned with establishing a relation between dinocysts and sea surface CO2 levels that can be the basis for comparisons. The proposal used to acquire research funding describes the aim of the biological experiments as follows:
[T]hese analyses will provide the boundary conditions to constrain the generation of empirical models that describe the influence of the environmental variables on dinoflagellate and dinocyst chemistry, specifically δ13C and composition. (Project proposal, p. 11)
This description illustrates the need to standardize the relation between dinoflagellate cysts and CO2 in that it seeks ‘boundary conditions’ that will lead to ‘constraints’ on empirical models and hence on outcomes. It is here that the researchers seek a quantifiable relation between CO2 and dinoflagellate cysts that can subsequently, as a fixed ‘module’, be shifted across time scales in a form of modular comparison. This description is a good way of summarizing the emic take on their work, yet it is too general for an understanding of the ways in which comparability is achieved practically. That is what my fieldwork was aimed at understanding, so I next provide my own rendition of the experiments, using excerpts from my fieldwork notes, in order to more closely scrutinize the practice of comparison.
Experimental setup: Stable isotopes as comparability devices
Although the proxy to be eventually used for comparative paleoclimate research is the fossilized cyst of dinoflagellate algae, the biological culturing experiments were undertaken using living dinoflagellates in their ‘active’, non-cyst forming stage. As will become clear below, this then entails ensuring that the carbon composition of the cysts is analogous to the carbon composition of the active algae. The experiments were designed to grow algae in water of different CO2 concentrations, in order to assess how they take in CO2. Once that would be known, their carbon composition – or more specifically, the carbon composition of the cysts they produce – could be considered a ‘proxy’ for ocean and atmosphere carbon concentrations as the algae would ‘reflect’ these in the organic carbon they contain. Crucially, the experiments were not done on the dinocysts, although the carbon composition of the cysts is the focus of research, since dinocysts are the only fossil remains in sediments and nothing of the dinoflagellates remains.
The researchers wished to be able to determine the carbon isotopic composition
3
of fossilized cysts in order to then, based on knowledge of dinoflagellate carbon acquisition gained in culturing experiments, draw conclusions concerning past carbon cycles and climates. Stable isotopes were thus the key to comparability, but they needed to be measured and ‘stabilized’ in their relation to dinoflagellate acquisition. In the case I studied, the experiments were a way to determine the relation between the types of carbon isotopes in dinocysts and the types of carbon isotopes in the surrounding water. Efforts were aimed at distinguishing 13C from 12C. Such differences are measured by mass spectrometry and expressed in terms of a δ-value. δ13C, for instance, is in itself what one could call a comparability device. It expresses the relative presence of 13C and 12C in a sample in relation to a standard (usually the Pee Dee Belemnite). In the experiments I observed, the difference between the δ13C of the water and the δ13C of the dinoflagellates was of great interest. Mediating between the two was the ‘carbon fractionation factor’ for dinoflagellate 13C, which (in this case) reflects the ‘preference for’ or ‘discrimination against’ a particular type of carbon isotope in the transformation of inorganic to organic carbon. Fractionation is usually expressed in terms of differences in isotopic ratios of different compounds. In the case I studied, it was crucial that the dinoflagellates use both CO2 and bicarbonate ( They like to take up lighter CO2, which is the 12C isotope. If there is more [CO2] they can choose. If there is less they take what they can get.
The experiments were designed to fix the relation between ‘Particulate Organic Carbon’ (POC) and ‘Dissolved Inorganic Carbon’ (DIC). As the lab researcher said, POC refers to ‘what has been taken up in the cell’. DIC, on the other hand, refers to the carbon dissolved in the ocean. The core of the experiments was explained by the lab researcher as follows:
We study their biology [i.e. the dinocyst biology], how they react to different CO2 concentrations for example, and then we apply our knowledge to cysts we find from sediments.
Overall, more 12C in the dinocysts was taken to be indicative of a relative abundance of CO2 in the ocean, because the algae have to resort less to the heavier isotope (13C). The dinocyst is thus treated as a ‘mirror’ (a term used by respondents) of CO2 levels, and proxy calibration – by this analogy – could be seen as learning the angle of reflection and the possible refractions of the mirror. For the experiments, dinoflagellates were kept in sterile water bottles that were aerated with air of a known CO2 composition (Figure 2). The bottles were kept in darkness when no researcher was present, ‘just in case’, the researcher told me, ‘there is anything in there that may grow’.

Dinoflagellates growing in bottles aerated with controlled CO2 levels.
In order to provide optimum conditions for the cells, sea conditions were mimicked by placing them in round bottles on spinning bands, providing them with regular movement (Figure 3).

Dinoflagellates on spinning bands to simulate sea conditions.
The fractionation factor for 13C can be obtained using one CO2 level. However, researchers were interested in how fractionation changed with changing CO2 levels. More CO2, for instance, might mean that more 12C is available for the cells. How the cells’ preference changes with changing CO2 levels required comparison:
Since we are interested in how 13C fractionation is changing with CO2, we need at least 2 concentrations (more are always better, of course). (Email correspondence)
Four CO2 ‘treatments’ were used to establish how 13C fractionation changes with CO2. They had the following compositions: 180 ppm (parts per million), corresponding to the value of the Last Glacial Maximum; 380 ppm, corresponding to contemporary levels; 800 ppm, corresponding to IPCC projections for 2100; and 1200 ppm, corresponding to IPCC projections for 2200. The latter two values also correspond to distant past values, specifically to cases where a rapid carbon injection had taken place, such as during the Paleocene–Eocene Thermal Maximum (PETM; around 55.8 Ma). There was a constraint on the number of comparisons used that had to do with common use and equipment. As the researcher indicated in an email exchange,
Eventually, these four CO2 concentrations are also the ones commonly used here …, and since we have only four gas lines … ;-) but it makes sense.
Such constraints defined the parameters of comparability, but there were many other constraints, as well as several comparability controls, as I propose to call these comparability techniques. The next section is devoted to their discussion, which brings into view the complexity of and attention to making things comparable.
Comparability techniques
The establishment of the relation between ocean CO2 and organic carbon inside the dinoflagellates can be seen as a form of standardization because it then becomes the fixed input for comparisons of carbon cycles and climates across geological time scales. The aim was to ‘fix’ a set of relations between cyst δ13C and ocean water δ13C and then to be able to use that standardized set of relations for comparative purposes across time scales. Quantitatively, measurements of δ13C were performed by mass spectrometry. For the purposes of this article, these measurements are of less concern than the comparity work that precedes them. In order to standardize the relations between cyst and ocean carbon composition, a number of what may be called comparability techniques were used that are crucial in understanding this standardization as part of comparity work. Although the distinction is for analytical purposes, I shall distinguish comparability constraints from comparability controls. While both require practical, experimental work, constraints set certain parameters for the experiments, whereas controls are devised ‘in’ the experiments.
Comparability constraints
Comparability constraints resemble ceteris paribus assumptions in that they cordon off a comparity space to allow for a limited number of variables. Only the changes that were deemed relevant and measurable were considered ‘variables’, and therefore comparability constraints could in this case be considered as ways to accomplish commensurability. Researchers often reflected on this and came up with provisos, qualifications, and reflections on uncertainties regarding their work. Nonetheless, the standardized quantification in the form of a fractionation factor (or the difference between POC δ13C and DIC δ13C) involved a singling out of relevant quantifiable factors. That in fact means that these factors were not only measurable but also manipulable beyond the manipulability required for any measurement. In that sense, a comparability constraint is not a mere cognitive parameter but requires an active effort to exclude and select.
Two comparability constraints in particular were important in the work I observed. The first concerns the assumption that the dinoflagellate (cysts) researched had not undergone significant evolutionary changes for many millions of years. One could call this the comparability constraint of evolutionary constancy. The species Gonyaulax spinifera, for instance, is considered to have existed relatively unchanged for over 120 Myr. ‘Relatively unchanged’ can here be taken to mean that the dinoflagellates are stable for purposes of comparative standardization. Without evolutionary constancy, contemporary experiments would not yield values usable for comparison of climates in deep geological time.
This poses a dilemma to which the establishment of comparability must answer: one can establish a relationship in a laboratory in the present, but how is one to know that the relationship holds up for fossils of the ‘same’ algae millions of years ago? If the algae are not evolutionary similar, they may respond differently to environmental influences and may have different intakes of carbon isotopes. In that case, the relation established cannot be treated as a mobile measure of events, but has limited temporal use. The point of working with the algae, however, is that they promise to be one of very few fossils that allow for comparisons into deep geological time. It is imperative, in other words, to establish that the algae experimented upon can be treated as the ‘same’ species in order to calibrate a proxy that can operate as a mobile module across time scales.
The comparability constraint that settles the issue of evolutionary constancy is achieved by means of other kinds of comparison. Establishing that fossil algae along a continuous time scale have not significantly evolved is, in the end, done by a comparison of the structure and composition of algae from various periods, scrutinizing exemplary pictures of types (genera and species) and samples under a microscope. When they do not use materials defined by other sources, researchers do this by hand, looking through a microscope at samples of fossil algae cysts and comparing them with pictures and evolutionary descriptions of exemplary types (genera and species) from books or from online databases. Figure 4 is an example of a ‘model’ dinocyst that serves to identify genera and species, taken from an online database.
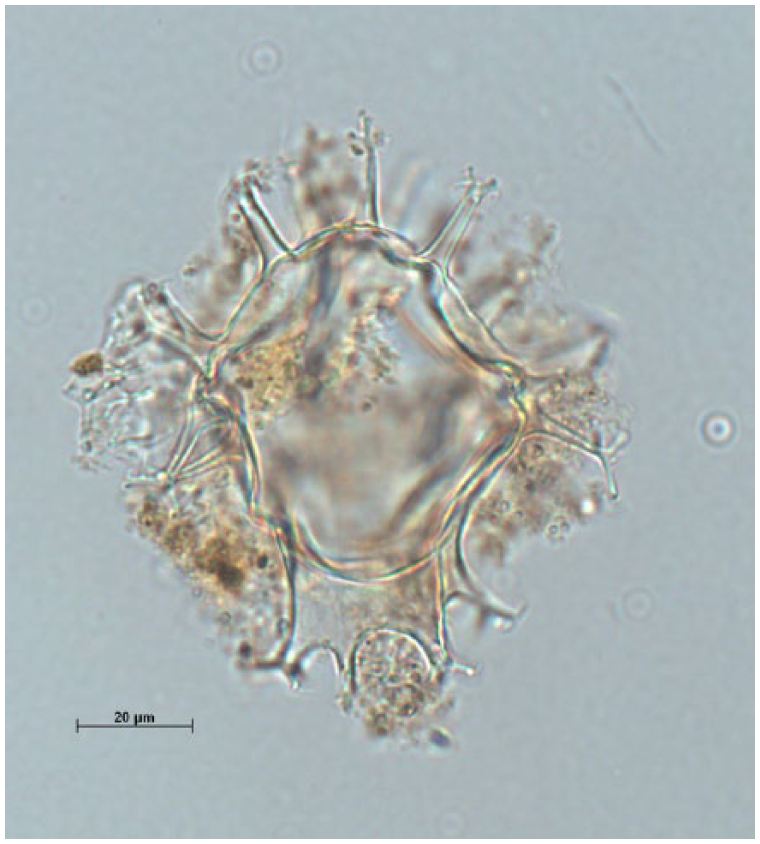
Spiniferites mirabilis dinocyst.
Specialized researchers have the experience to determine species on this purely visual basis. In case they need to learn more about the species, their morphology, their classification and names in existing literature, and the periods in which they occurred, atlases of dinoflagellates exist, although even those atlases may recognize the ‘special problems’ with and hence the ‘subjective’ character of some species assignments (e.g. Stover and Evitt, 1978: 5). The comparability constraint of evolutionary constancy is only possible on the basis of canonical and disseminated work that entails contingencies and uncertainties that are in the end and in liminal cases only resolvable through ‘subjective’ judgments.
A second comparability constraint concerned the similarity in terms of carbon composition between dinoflagellate cells and the cysts they form during the hypnozygotic stage in their lifecycle. One could call this the comparability constraint of cell-cyst carbon continuity. As noted above, the algae produce a kind of organic plastic hull in which they reside in their dormant phase, and these cysts are the only thing that remains of the algae as fossils. The researchers I observed were interested in using dinocysts for comparative purposes (for comparing climates along the geological time scale), but they used dinoflagellates during their ‘motile’ stage (i.e. active cells) for the experiments aimed at forging a proxy out of the dinocysts. In order for that proxy to be plausible, no new carbon should enter the cyst while it is being formed. Like the comparability constraint of evolutionary constancy, the cell-cyst carbon continuity constraint rests on another form of comparison, namely, the comparison between dinoflagellate and dinocyst. This comparison is in a way ‘worked around’ by considering the rate at which cyst formation takes place. While visiting the culturing experiments lab, the researcher told me that since cyst formation often occurs in a small amount of time (sometimes only ten minutes), ‘we assume that all the organic carbon that exists in the cyst is already present in the dinoflagellate’. To corroborate this, she showed me a time-lapse video of cyst formation. The rate of cyst formation thus becomes a proxy for a direct comparison between dinoflagellate cell and dinocyst. This is all the more important because, to my initial surprise, it turned out that the dinoflagellates used in the experiments do not produce dinocysts at all. They are even incapable of doing so. In part, this has to do with the difficulty of breeding cyst-producing dinoflagellates from culture banks. To her frustration, she had tried to do so over the course of several months but it hadn’t worked:
I just went to Canada for three months to induce cyst formation and to look at the difference between the cells and the cysts, and it didn’t work out. … Sometimes they don’t like each other, and they don’t hatch.
While talking with this researcher, I asked whether taking dinoflagellates out of the ocean would not be an option. She said it would not because of difficulties in comparing them, since the circumstances in which they had grown might differ. Also, she would have to delve into the atlases discussed above, but in her case – being an experimental researcher and not an expert in species determination – that would be difficult: ‘Then I need to identify the species and I’m not trained to do that. It’s not so easy, because the cysts look really different but the cells look very similar’. Using clones thus saves the trouble of ensuring evolutionary constancy by manually identifying species. Perhaps most importantly, though, it is actually common to work with non-cyst-producing algae because cyst formation would potentially ‘pollute’ the experiments: ‘I need to make a statement about what happened and if there’s cyst formation it’s not clear anymore’. In order to nonetheless plausibly assume cell-cyst carbon continuity, the speed of cyst formation provides the comparability constraint. A formation this fast has to make do with the organic carbon already inside the dinoflagellate.
Comparability controls
The use of dinoflagellates that produce no cysts can be considered a comparability control. It entailed the selection of clones, called ‘strains’, for the experiments, for what we might call a difference control. The following conversation, excerpted from my field notes, clarifies the use of strains:
They don’t form cysts, because I work with strains, clones.
What are strains?
They are like clones. One single cell that has been isolated by someone some time and it has divided further and further and it’s just a clone of itself.
Why do you work with such strains?
Because they [non-strains] may react slightly different. Also because I want a clean sample. I’d rather want a constantly growing sample. In the end also for publication it’s good to work with strains and then say we have been working with this strain and it’s all reproducible.
(R = researcher (respondent); A = author)
Strains come from culture banks, usually held in private companies, and they provide a difference control in that researchers are more likely, given their standardization, to get similar results when experimenting on them. In another scientific context, Karen Rader (2004) has illustrated how mice were standardized to serve as model organisms for research (see also Ankeny and Leonelli, 2011; Nelson, 2013; Sismondo, 1997). That standardization has everything to do with crafting a comparity space, in part by privileging certain planes of comparison. By reducing difference in the experiments, one controls which difference becomes the locus of interested variation. Since response to CO2 levels was the focus, difference in other variables was minimized. But this meant that the variable on which variation occurs is dependent, in the meaningful interpretation of its values, on various selections and suppressions in the larger ecology of the experiment. On one hand, having ‘a clean sample’ enabled the interpretation of outcomes. On the other hand, it introduced risks in the form of uncertainties. As the researcher said,
I think it’s actually better not to do that but it’s just so common. … There is also the question whether they react differently between strains and I think there are differences but the question is how big the differences are.
Using standardized strains presented the researcher with limitations that had much to do with comparability, since, in her opinion, more difference would be better: ‘the culture collections we’re using have one or two strains we can order, but you need more’. In part, however, she made clear that scientific conventions favored using strains. That which is, in her words, ‘common’ is also a scientific control: reproducibility and, as she said, the possibility of ‘making a statement’ entail this form of difference control. Although it presented its own problems, without it, outcomes would be ‘harder to interpret’, and hence comparability would be jeopardized.
A second comparability construct in the experiments concerned the limitation of the number of cells in the bottles. The researcher worked with ‘low cell densities’, in order to avoid what she called ‘a brown soup’. That is, with many cells in the bottle, various values, such as pH and alkalinity, become more difficult to measure:
With too many it’s difficult to measure the isotopic composition of the CO2 in the bottle compared with the cells, because it’s changing rapidly. Usually I measure every week. I relate the isotopic composition of the dinoflagellates to the isotopic composition of the inorganic carbon in the water and in the air in the bottle. If it changes all the time [which would happen with many cells in the bottle] I cannot do that. I would have to measure every day or more often, and I would not get a stable result.
In order to ensure a stable comparison between water and cells – a comparison crucial in establishing a proxy for comparative purposes in paleoclimate research – controlling density was thus crucial.
Finally, there was a work of comparability control concerning the ‘leakage’ of CO2 out of the dinoflagellate cell. Some amount of CO2 leaks from the cell after being taken in. In order to accurately calibrate the relation between carbon composition and seawater CO2, it is important to determine the loss of CO2 through leakage: ‘The relationship between carbon fractionation and CO2 is what interests us. We want to know the leakage and the carbon source to understand what we see’. Leakage was measured by means of membrane inlet mass spectrometry, which entailed measuring fluxes of CO2 in and out of the cell during light and dark cycles (one researcher described this as ‘live photosynthesis’). Leakage was species-dependent, so it had to be measured for each type of strain. Without determining leakage, the precise relation between the carbon composition of fossil dinocysts and the marine circumstances in which they grew would not be possible, eliminating comparability.
Proxy calibration: Comparing comparison
In order to test the relations established in the culturing experiments, the proxy (dinoflagellate fossil cysts) would be used to reconstruct sea surface temperatures and, through that, atmospheric temperatures, over a relatively recent time period, for which reliable reconstructions exist by other means (for instance, by ice sheet measurement). In other words, the comparative relation was to be calibrated further by means of a comparison with a validated method of comparison. This is one reason why ‘multi-proxy research’, which my respondents considered ‘advanced’, was favored: each proxy could function as a control on the others. The subproject for which ‘calibration’ was used as an emic term focused on the past 200 years, for which atmospheric CO2 values are considered known, but the wider time frame of 800,000 years for which ice core reconstruction is considered valid was mentioned as feasible for calibration as well. The aim was to focus mostly on a particular species of dinoflagellate (Spiniferites), which has the property of being ‘autotrophic’. This means it directly absorbs CO2 dissolved in the ocean water. If a relation between organic carbon in Spiniferites and atmospheric CO2 could be established, the project could be considered to have successfully calibrated this particular type of algae as a proxy, which from then on contains a relationship that can be established for millions of years ago on the basis of dinoflagellate fossil cysts.
In this calibration, the algae fossils need to perform, to show organic carbon ratios that, when calculated ‘back’ to established, validated ocean carbon ratios, yield values very closely resembling those calculated on the basis of existing ice core research. As one researcher wrote in an email, she aimed
at applying the species-specific 13C fractionation models to reconstruct CO2. Since there are reliable CO2 reconstructions based on ice-core data for the past 800kyrs, this will certainly help to validate the proxy.
If validated, the proxy will provide the basis for what I have here called modular comparison. That is, it may be blackboxed as a fixed or even standardized ‘module’ that can be used to compare different past climates along the International Geologic Time Scale. What the black box contains is a calibrated relationship, an ontological claim concerning the being of dinoflagellate cysts and their marine environment, specifically concerning their respective carbon compositions and the transfer of marine carbon to algae (cyst) carbon composition.
Conclusion: Making things comparable
Making things comparable, as we have seen, involves a variety of practices that are themselves comparative. In conclusion, I will draw out some of the forms of comparison involved in the practices of comparability I studied. They can be seen as a typology of comparison and they may be of use in analyzing other cases of comparability and comparison. The kind of comparison that was aimed for in the comparability practices I studied is what I propose to call modular comparison. This is a form of comparison on the basis of a standardized relation that is considered generalizable across a certain comparity space (in my case study a space of climate phenomena in chronological time). Such a standardized relation (in this case a relation between algae and ocean CO2) can be considered as a module that can be shifted across scales. Modular comparability requires the calibration and standardization of a proxy as a mobile module that relationally connects two or more events and that, being rendered mobile, can be shifted along a temporal or spatial axis to be applied elsewhere in the same object space. The relation thus specified in a modular type of comparability allows for the qualification and/or quantification of its terms at each point of the comparative scale.
The relation that is established as a mobile module in comparability practices resembles what Stengers has called a ‘rapport’. By this she means not just a relation between objects or entities but more fundamentally a logos of their connection that specifies how the objects exist and can be observed and quantified. She discusses the importance of experimental setups which ‘authorize [scientists] to claim that the subject matters that they address lend themselves to quantitative comparison’ (Stengers, 2011: 49). The creation of ‘rapport’, in other words, involves rendering a particular relation useful for comparability purposes. As an example, she mentions Galileo’s inclined plane, which enabled him to compare degrees of velocity. The entities whose relationship is established themselves participate in the process of comparison, and the creation of a rapport enables scientists ‘to claim that they benefit from states of affairs that they did not impose, that pertained to the phenomena studied, and that therefore could be turned successfully into tools for making comparisons’ (Stengers, 2011: 51). The experiments paleoclimatologists devised in order to establish the relation between algae and CO2 did not follow a designated path but were constantly adjusted to the recalcitrance of matter, for instance, when isotopic ratio measurements involved laser-burning samples that kept shooting away instead of burning. Describing Stengers’ account of comparison, Helen Verran (2011) speaks of ‘the painstaking labor of solidifying comparison as a thoroughly reliable participant in the here and now’ (p. 69). Such work may be understood as a work of rendering things comparable by establishing a comparity space.
My analysis here is intended to highlight this ‘painstaking work’ to further qualify what Stengers describes in a very general way as ‘rapport’. It involves the establishment of the way two phenomena respond and correspond to each other, which preferably constitutes a causal relation or ‘mechanism’, or simply a (statistical) correlation, and it then involves the supplanting of this relation, as it were fixed in a mobile module, to different times. Comparison then becomes possible by moving this relation up and down a temporal scale and by, for each selected sample, empirically qualifying and/or quantifying the given variable in the relation – the ‘proxy’ – and, thence, by way of deduction, qualifying and/or quantifying the other, which is generally the variable of interest. Values can be plotted to show, for instance, a temporal sequence in which sea surface temperature increases, sea ice volume decreases, or atmospheric CO2 increases. Figure 5 provides an example from a published research paper, in which drill core depth is correlated with geological time. The figure is part of what can be called an information infrastructure for comparison, 5 but it visually represents what is, in a more abstract sense, a comparity space, since various carbon proxy values appear relative to each other in the same space (here, the first column of the graphic visualization).

Data plot showing (selected sections) drill core depth and carbon proxy (δ13C), including indications for ‘paleo-events’ such as ‘PETM’, which refers to the Paleocene–Eocene Thermal Maximum, which occurred between 56 and 55 Ma years ago.
Figure 5 at the same time illustrates how chronological comparison is enabled through an object space, which the figure visually expresses. So, once the relation between algae and CO2-acquisition has been established in quantified terms, the proxy – dinoflagellate cyst carbon composition – can be considered a ‘paleothermometer’, and the comparison of climates becomes possible.
Establishing modular comparability in the proxy calibration I studied involved other forms of comparison. In order to calibrate a proxy, for instance, microscopic visualization needs to take place to sort out different types of algae, sometimes using reference books. This involved a form of what could be called figural comparison. In figural comparison, comparing two or more objects or events is to consider how well each can function as a picture of the other. A special type of figural comparison is what can be called typological comparison, when phenomena are compared against some standard, for instance, in the form of an archetype. In the context of paleoscience, one might think here of Hegel’s anti-evolutionary interpretation of fossils: as early, primitive renderings of forms that were later to fully crystallize. The fossil record, for Hegel, was an account of the less-developed embodiments of Idea and he did not consider fossils to be remains of living creatures, but merely primitive ‘anticipations’ of life forms by inorganic nature (Magee, 2010: 85–86). Contemporary paleoscience clearly does not hold this view of fossils, but it deploys forms of typological comparison, for instance, in the selection of strains of algae and in the use of the ‘dino atlas’, which contains standardized images of types of algae.
Finally, in professional comparison, comparison is in the hands of gifted and trusted subjects. The scientists calibrating the paleoproxy discussed in this article obviously count as such, as they are able to make recognized claims to ‘professional vision’ (Goodwin, 1994). Professional comparison thus enters the other forms of comparison deployed while establishing modular comparability. In the end, it is a crucial part of determining who gets to compare climates. There are other forms of comparison, such as organic comparison (a comparison of parts to a whole), but here I mention only the ones encountered in the analysis of proxy calibration.
This study has shown how comparison is done by means of a comparity work that involves the use of various comparability techniques. I have discussed comparability constraints and comparability controls. Constraints concerned the relative evolutionary constancy of algae and the continuity between cells and algae cysts in terms of their carbon composition. The comparability controls of this study were specific to the experiments in question, dealing with factors such as difference, density, and leakage. Obviously, these constraints and controls do not translate literally to other cases of establishing comparability. But comparability constraints and controls are at work across the sciences. Cordoning off a comparity space for the making of comparative evaluations by introducing ceteris paribus clauses is widespread in science and elsewhere.
Comparability hinges on stable reference points and on limits to the number of relevant actors in the comparison. I hope that the analytical vocabulary developed in the course of this study will prove helpful in the analysis of other kinds of comparability practices, including a variety of practices of monitoring, qualification, quantification, and evaluation. I believe that any approach foregrounding relations or the relational is aided by recognition of the ways in which relations are spun out across comparative practices. To relate, after all, is always also to bring under comparative scrutiny within some shared object space. Relations are thus to a significant degree conditioned by comparability. In a less general sense, scrutinizing how comparability constraints and controls are deployed and the variety of types of comparison involved therein may help the analysis of various kinds of classification, standardization, and translation.
Let me end with the recognition that there is, of course, a politics of comparison. Comparability is a complex achievement that, although constituted by an often invisible comparity work, tells a great deal about the ways the world – in this case, climate change – is rendered observable. To observe climate change is first to control and constrain a variety of objects – among others, in this case, algae and algae fossils – so that they may become useful in comparing. These are uncertain, contingent practices that likely never will enter global warming debates, but such practices are nonetheless vital to defining the stakes in these debates. And at times, the kind of technically detailed, intricate work I have discussed here does become the center of attention in these debates. Perhaps the most (in)famous example of this, the so-called hockey stick controversy, actually occurred in relation paleoclimatology and the type of proxy records used for temperature reconstruction (cf. Frank et al., 2010). Because ‘climate change’ is a comparative phenomenon, the kind of proxy calibration discussed here is crucial to the constitution of that phenomenon. At times, it becomes visible that the politics of climate change is a politics of comparison, and then the invisible work of establishing comparability may be critically scrutinized. Such scrutiny has not been my goal here. Rather, I have sought to uncover some of the intricacies in the work of establishing comparability, and I would argue that such intricacies are also important in getting a better grasp of the politics of climate change. Climate change becomes political when, beyond being a merely comparative concept, it constitutes a particular pattern of change. One can, for instance, claim that ‘global warming’ is a better concept than ‘climate change’ because the latter is a more depoliticized concept (Morton, 2013). Such a pattern can then be visualized, most famously, for instance, by means of a ‘hockey stick’ graph or by means of color-coded risk graphics (Mahony and Hulme, 2012). But such visualizations, and the politics in which they are enmeshed, rest on an intricate and often invisible work of ensuring comparability that occurs in myriad places in the monitoring of climate change and of which this article has provided one detailed example. Even disagreement about climate change plays out with reference to a comparity space usually agreed upon. Comparison and the work of making things comparable are at the root of ways of rendering global warming or ‘the Anthropocene’ visible and of politicizing them. Ensuring comparability, sorting out difference and similarity, is never innocent.
Footnotes
Acknowledgements
This article benefited greatly by comments from the following participants in the Monitoring Seminar at Erasmus University Rotterdam: Jess Bier, Sanne Boersma, Maja Hertoghs, Anne Slootweg, Friso van Houdt, Irene van Oorschot, and Rogier van Reekum. Likewise, comments by three anonymous reviewers, as well as by the Editor of Social Studies of Science, Sergio Sismondo, have been of great help. Last but not least, I thank my respondents for their generosity in sharing their everyday work practices with me. It has been great to learn from them.
Funding
This research is part of the European Research Council (ERC) funded project ‘Monitoring Modernity: A comparative analysis of practices of social imagination in the monitoring of global flows of goods, capital and persons’ (ERC Starting Researcher Grant, project number 283679).
Notes
Author biography
![]() ). Forthcoming from Cambridge University Press is Imagining Societies: A Critique of Immigrant Integration in Western Europe.
). Forthcoming from Cambridge University Press is Imagining Societies: A Critique of Immigrant Integration in Western Europe.