
Abstract
Cryoconite is granular sediment found on glacier surfaces comprising both mineral and biological material. Despite long having been recognised as an important glaciological and biological phenomenon cryoconite remains relatively poorly understood. Here, we appraise the literature on cryoconite for the first time, with the aim of synthesising and evaluating current knowledge to direct future investigations. We review the properties of cryoconite, the environments in which it is found, the biology and biogeochemistry of cryoconite, and its interactions with climate and anthropogenic pollutants. We generally focus upon cryoconite in the Arctic in summer, with Antarctic and lower latitude settings examined individually. We then compare the current state-of-the-science with that at the turn of the twentieth century, and suggest directions for future research including specific recommendations for studies at a range of spatial scales and a framework for integrating these into a more holistic understanding of cryoconite and its role in the cryosphere.
I Introduction
Cryoconite is granular sediment found on glacier surfaces comprising both mineral and biological material. Due to its dark colour, cryoconite efficiently absorbs solar radiation and ‘drills’ quasi-circular holes up to tens of centimetres deep into glacier ice surface. These are known as cryoconite holes, and are very common features on ablating ice surfaces worldwide, although cryoconite can also be found in other environments such as stream deposits and scattered on ice surfaces (Hodson et al., 2008). Despite having been recognised as important glaciological and biological entities in the nineteenth century (e.g. Agassiz, 1846; Nansen, 1906; Nordenskiöld, 1875) cryoconite and cryoconite holes remain poorly understood. They are thought to be ‘ice cold hotspots’ of biodiversity and microbial activity on glacier surfaces (Edwards et al., 2013a) and are regularly examined; however, studies have been predominantly reductionist in approach and based upon assumptions of discrete habitat boundaries and simple thermodynamics established by early polar explorers over a century ago. This review begins by discussing the properties of cryoconite and the environments in which it is usually found, before summarising current knowledge of cryoconite biology and discussing its wider significance. We generally focus upon cryoconite in the Arctic in summer, with Antarctic and lower-latitude settings examined individually. We conclude by comparing the current state-of-the-science with that at the turn of the twentieth century, and suggest directions for future research.
II What is cryoconite?
The term ‘cryoconite’ (from the Greek ‘kryos’ and ‘konis’, meaning ‘cold-dust’) has been used interchangeably to describe the entire mass of insoluble impurities, including discrete bio-engineered granules (Edwards et al., 2014a; Langford et al., 2010) on glacier surfaces or the discrete granules only. In this paper, particles that do not contain both biotic and abiotic components are not considered to be cryoconite, being inorganic supraglacial debris or cryo-tolerant biota. We consider this distinction to be important since the presence and action of biota, along with geological and hydrological processes, influences the formation, morphology and biogeochemistry of cryoconite (Hodson et al., 2008; Langford et al., 2010; Takeuchi et al., 2001b). In this paper, cryoconite refers to
Discrete, aggregated granules of mineral and organic matter, either within cryoconite holes or elsewhere in the supraglacial zone.
1 Composition of granules
The materials comprising cryoconite can be divided into two main types: organic and inorganic. Organic matter (OM) includes living and dead microbes, their exudates, products of decomposition and allochthonous biotic and biogenic matter (e.g. Hodson et al., 2008; Langford et al., 2010; Takeuchi, 2002a, 2002b; Takeuchi et al., 2010; Wientjes et al. 2011). While there is clearly variability in OM abundance in cryoconite worldwide (Table 1) it invariably represents a key component of cryoconite granules. Spatial variability in OM has also been identified across individual glaciers (Langford et al., 2014; Stibal et al., 2010, 2012a). The quantity and quality of OM is known to influence aggregate formation in terrestrial soils, but this has rarely been examined in cryoconite (Langford et al. 2014).
Percentage fractions of organic matter (OM) in cryoconite samples (error is 1σ).

In addition to biotic and biogenic OM, persistent organic pollutants (POPs) with carcinogenic and mutagenic potential (Hodson, 2014) can be incorporated into cryoconite. These molecules resist environmental degradation and might bioaccumulate in cryoconite. Whilst they likely have little impact on-site, they may influence downstream ecosystems. These pollutants have been linked to industrial emissions and are often deposited along with inorganic matter such as black carbon (BC) (Aamaas et al., 2011). BC, a product of incomplete combustion of fossil and biofuels, has attracted research attention because it is an extremely effective absorber of heat, both in the atmosphere and after deposition on ice and snow, possibly accelerating melt (Clarke and Noone, 1985; Hansen et al., 2000; Jacobsen, 2004; Xu et al., 2009a). The longevity of BC on ice surfaces is probably enhanced by entrainment into cryoconite, likely reducing the aggregate albedo (Bøggild, 2011; Xu et al., 2009b), although it is also possible that cryoconite microbes metabolise BC and reduce its potency.
Inorganic matter in cryoconite is dominated by mineral fragments, often dominated by phyllosilicate, tectosilicate and quartz (e.g. Edwards et al., 2011; Hodson et al., 2010a; Langford et al. 2010; Stibal et al., 2008a, 2008b); however, differences in source geology likely cause geographic variation in cryoconite mineralogy. Bullard (2013) pointed out that supraglacial dusts (that presumably form cryoconite granules) generally include materials from various local and distal sources, citing studies undertaken in Arctic Canada (Lawrence and Neff, 2009; Zdanowicz et al., 2000), Antarctica (Bory et al., 2010), the central (Drab et al., 2002; Prospero et al., 2012; Tegen and Rind, 2000) and south-west marginal zones of the Greenland Ice Sheet (Bøggild et al., 2010; Wientjes et al., 2011). This was recently corroborated by Nagatsuka et al. (2014) who used Sr and Nd isotopic ratios to describe contrasting origins for silicate minerals in cryoconite on a selection of central Asian glaciers. Fine dusts deposited from high atmospheric suspension are particularly important in the interior zones of large glaciers and ice sheets, while on smaller glaciers and ice-sheet margins a greater proportion of minerals are likely derived from local sources (Stibal et al., 2012a). While material can be transported from local sources (including exposed valley sides and moraines) to the ice surface by gusts of wind (Bøggild et al., 2010; Oerlemans et al., 2009), Porter et al. (2010) found unconsolidated morainic material to be reworked onto the surface of a Svalbard glacier by debris flows. The dominant sources of cryoconite minerals can also vary across glacier surfaces (Langford et al. 2011). Other possible sources include release from englacial storage by ablation (Atkins and Dunbar, 2009; MacDonnell and Fitzsimons, 2008; Wientjes et al., 2011), outcropping of basal tills (Stibal et al., 2012a) and deposition of micrometeorites (particularly noted on the Antarctic and Greenland ice sheets). Differences in source geology and depositional regimes likely explain geographical variations in the size of mineral fragments in cryoconite. For example, Takeuchi et al. (2010) found mineral fragments between 1.3 and 98 μm diameter in China, while Zarsky et al. (2013) found between 0.02 and 2000 µm in Svalbard.
The mineralogy of ice surface debris (including cryoconite) might influence spatial patterns of melt. Casey et al. (2012) found the mineralogy of supraglacial debris on Himalayan glaciers to correlate with surface temperatures and spectral reflectance. Since cryoconite mineralogy has been shown to influence its colour and reflectivity (Takeuchi et al., 2014; Tedesco et al. 2013) it may similarly influence ice surface albedo (Sugiyama et al., 2014). Mineralogy might also influence cryoconite microbes (Carson et al., 2009). For example, since cryoconite microbial communities are often phosphorus (P) limited (Mindl et al., 2007; Säwstrom et al., 2002; Stibal and Tranter, 2007), phosphates from rock debris might provide crucial nutrients. Tazaki et al. (1994) found silicate clays are entrained on snow algal cell surfaces. This is likely analogous to processes occurring in cryoconite, although direct evidence of mineralogical controls on cryoconite microbial activity has not yet been presented. Analysis of heavy metals has indicated the occurrence of mineral–microbe interactions (Nagatsuka et al. 2010) and Hodson et al. (2010a) highlighted the importance of cryoconite biota for extending rock-water interactions and catalysing chemical weathering. Dittrich and Lüttge (2008) suggested that microbes actively control water–solid interactions. Nevertheless, deep understanding of mineral–biota interactions in cryoconite is still lacking. Further inorganic material in cryoconite includes heavy metals (Singh et al., 2013) and radionuclides (Tieber et al., 2009), the accumulation and release of which may impact supraglacial and proglacial ecosystems.
The literature therefore indicates geographic variations in OM (further details in Section IV) and inorganic matter arising from different depositional and post-depositional processes. Quantifying the relative contributions of OM and inorganic matter sources and the impacts upon melt and microbial processes remain important outstanding research questions.
2 Shape and size of cryoconite granules
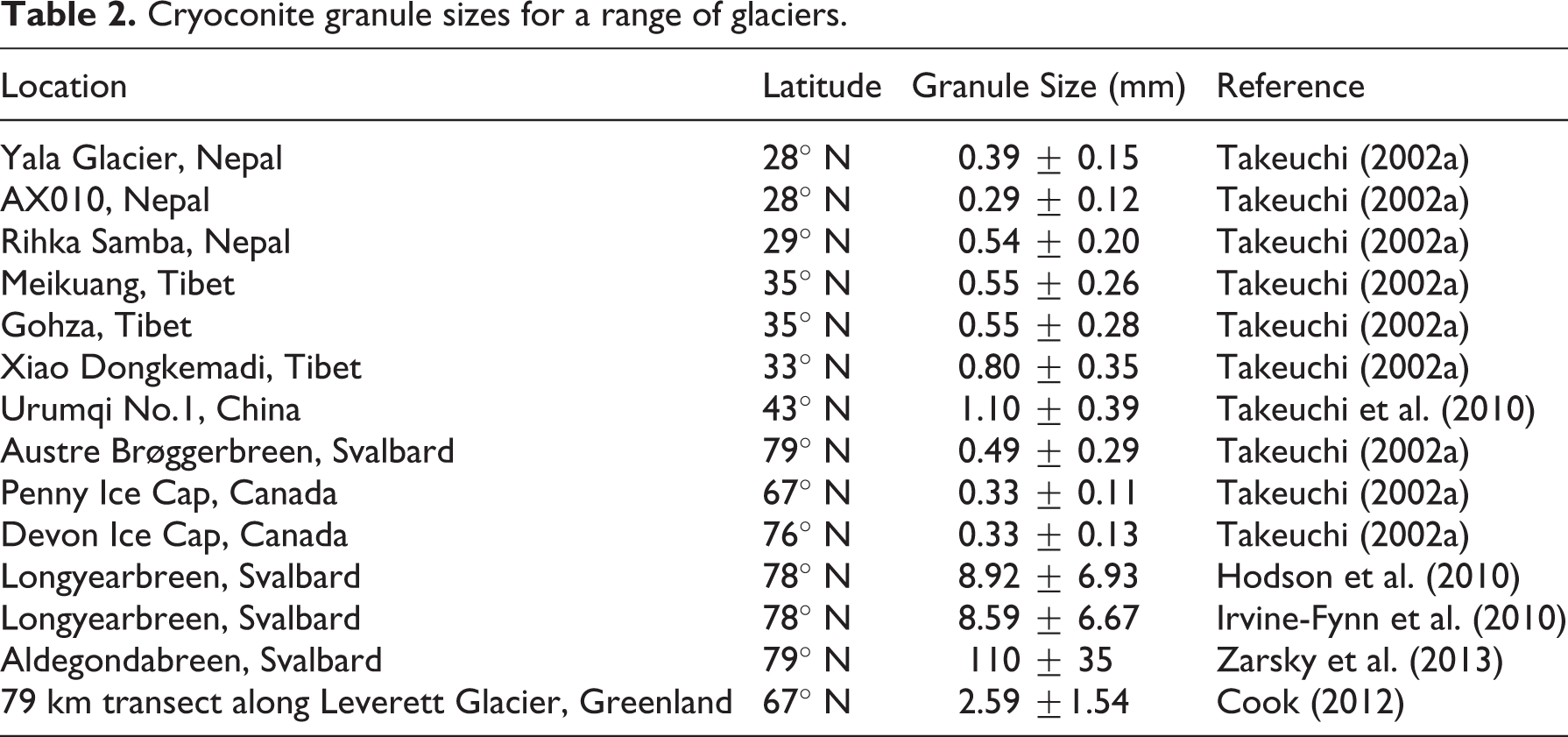
Cryoconite granules tend to be quasi-spherical, however their size and morphology is highly spatially variable (Figure 1, Table 2), likely reflecting the local balance between microbial growth, physical aggregation and erosion (Irvine-Fynn et al., 2010). Variations in granule size have been reported between glaciers; for example Zarsky et al. (2013) found larger granules to be more common on Aldegondabreen (Svalbard) than on other glaciers, possibly due to gentle slope gradients, fine material particles, nutrient input from debris avalanches and atmospheric deposition and high rates of microbial activity. Also on Aldegondabreen, Langford et al. (2014) identified an “edge effect” whereby granules were generally smaller towards the central areas of the glacier. This was attributed to biochemical and photophysical processes including TOC and carbohydrate concentration. The strong correlation between granule size and OM content (Langford et al., 2010, Stibal et al., 2010; Takeuchi et al., 2010) suggests that biota play a key role in determining the shape and size of cryoconite granules.

(a) Discrete quasi-spherical cryoconite granules of 1–2.5 mm diameter sampled ∼2km inland of the ice margin on the Greenland ice sheet near Kangerlussuaq; (b) a complex of cryoconite granules ca. 10 mm diameter, sampled ∼34km inland of the ice margin on the Greenland ice sheet near Kangerlussuaq. Scale bar applies to both images.
Cryoconite granule sizes for a range of glaciers.
Growth and proliferation of autotrophic cyanobacteria at granule peripheries has often been suggested to be a driver of granule growth because their filamentous morphology enables them to entangle debris (Langford et al., 2010). Heterotrophic microbes might limit this growth by decomposing OM (Hodson et al., 2010a). However, decomposition of OM produces ‘sticky’ humic substances that may enhance granule cohesion (Langford et al., 2011; Takeuchi, 2002a; Takeuchi et al., 2001b). Granule size is therefore probably co-limited by the binding ability of filamentous autotrophs, the adhesion potential of other OM and rates of decomposition by heterotrophs (Takeuchi et al., 2010). The potential for cryoconite microbes to control granule size and morphology was demonstrated by Takeuchi et al. (2001b) who treated half a batch of cryoconite with biocide (CuSO4) before leaving them on a glacier surface over an ablation season. Algal mats formed in untreated samples, whereas treated samples remained loose and disaggregated. The contrasting morphologies suggest that meltwater, wind or gravity driven movement of granules might influence their shapes. This is supported by the even distribution of cyanobacteria over granule surfaces (e.g. Hodson et al., 2010a; Takeuchi et al., 2001a, 2001b) despite only one side of settled grains being exposed to sunlight. Irvine-Fynn et al. (2011) showed continuous hydraulic redistribution (but very little net displacement) of cryoconite over a small plot on a Svalbard glacier that could be crucial for maintaining homogenous growth rates over their surfaces.
Cryoconite granules also exhibit distinctive internal microstructures (e.g. Langford et al., 2010; Takeuchi et al., 2001b, 2010) where granule interiors are dominated by mineral fragments and heterotrophic microbes, while granule surfaces are inhabited by photoautotrophs (e.g. Langford et al., 2010; Takeuchi, 2002a; Takeuchi et al., 2001b). This probably results from the extinction of light within the outer few mm of cryoconite granules. Indeed, Hodson et al. (2010b) showed both cyanobacterial photosynthesis and photosynthetically active radiation (PAR) receipt to be concentrated at granule surfaces. Thin section microscopy of Chinese cryoconite by Takeuchi et al. (2010) revealed more complex internal microstructures including concentric rings of OM that may have resulted from annual biomass accumulation or cycles of erosion and regrowth (Takeuchi et al., 2010). The number of rings per granule ranged from 2 to 7, with a mean of 3.5. Some granules contained sub-granules with their own concentric rings, suggesting amalgamation due to tight packing and rapid cyanobacterial growth. The amalgamation of individual granules might explain reports of large complex granules with diameters of up to 40 mm (Hodson et al., 2010a). Figure 2 shows four distinct classes of cryoconite microstructure proposed by Takeuchi et al. (2010).
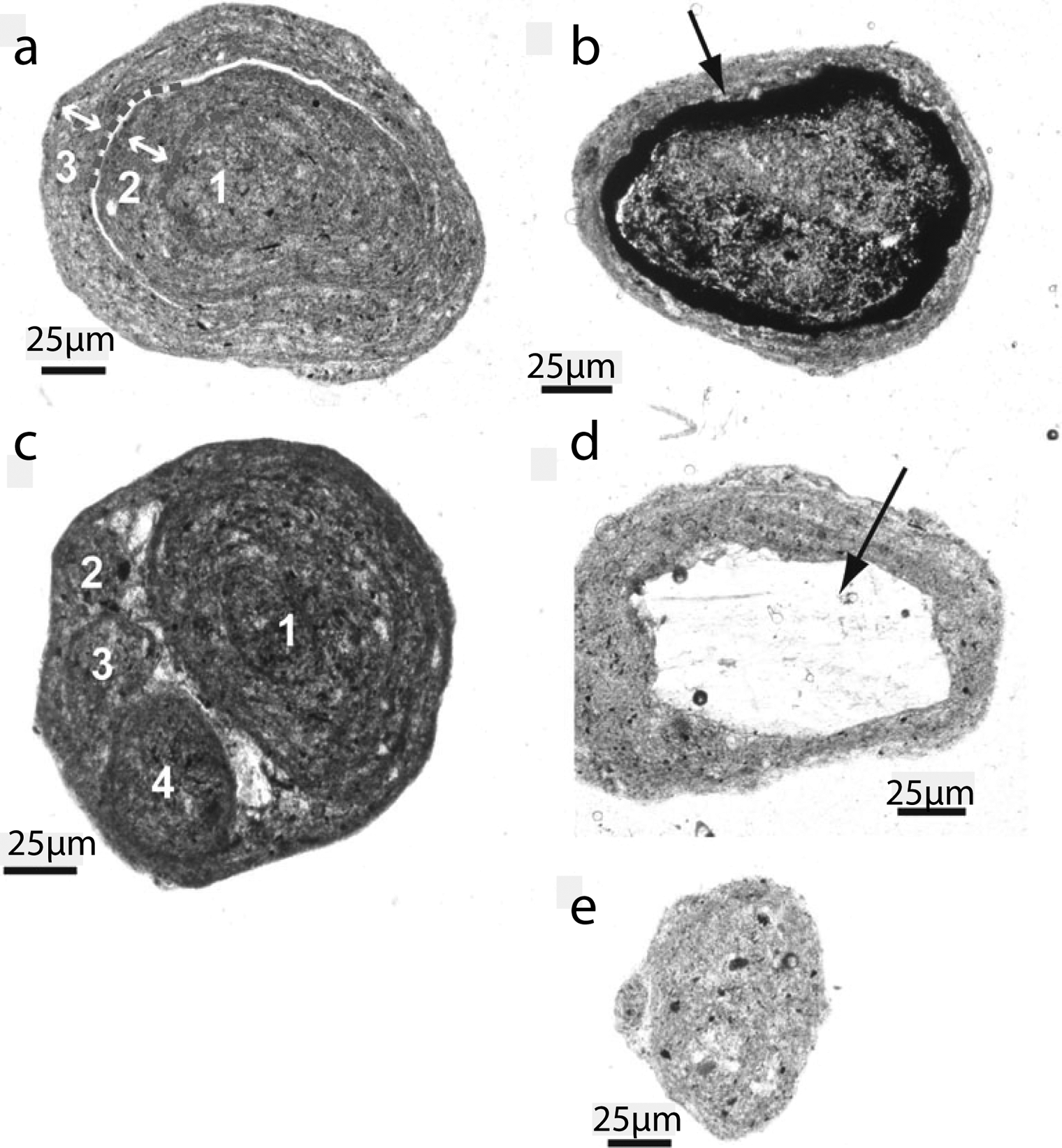
Cryoconite granule structures: (a) simple concentric ring structure; (b) granule containing a dark humic ring; (c) granule containing subgranules, each with individual concentric ring structures; (d) granule with a large central mineral grain; (e) an example of a granule with no specific internal structure. (Reprinted from Takeuchi et al. (2010), Annals of Glaciology with permission from the International Glaciology Society.)
In general, larger and more stable grains exist where cyanobacteria are more abundant (Takeuchi et al., 2010). This is because granule growth is driven by the growth and proliferation of microbes, entanglement of debris in cyanobacterial biomass and cementation by cohesive humic and extracellular polymeric substances (Hodson et al., 2010b; Langford et al., 2010, 2014; Takeuchi et al., 2001b, 2010). However, the precise biotic and abiotic processes controlling granule growth and erosion are still poorly understood, and we have little knowledge of their spatio-temporal variations. These are important knowledge gaps since larger granules probably have longer residence times, and therefore more prolonged albedo-reducing effect on ice surfaces (Bøggild et al., 2010; Hodson et al., 2010a; Irvine-Fynn et al., 2011) while also providing more microhabitats for varied microbial communities (Langford et al., 2010; Zarsky et al., 2013). This suggests granule size could have important glaciological and ecological implications, as well as potentially being an indicator of local biogeochemical processes (Langford et al., 2014).
III Cryoconite holes
1 Formation and evolution of cryoconite holes
Cryoconite exists in many several supraglacial environs (Figure 3) of which cryoconite holes are by far the most common and best understood. Cryoconite holes are quasi-cylindrical depressions on ice surfaces that commonly have depths and diameters ranging from millimetres to tens of centimetres. Cylindrical holes are common; however, complex shapes also form and likely reflect interactions between regional and local topography, aspect, ice type, hydrology and sediment dynamics (Cook et al., 2010; McIntyre, 1984).

A range of cryoconite habitats. (a) Wide shot of ice surface at ‘S6’, 38 km inland of the ice margin, Greenland ice sheet. Cryoconite holes are visible in the foreground, stream cryoconite and cryoconite mantle is visible in the midground. (b) Aerial view of cryoconite holes at the same location. (c) Loose cryoconite granules, fine dusts and algae dispersed upon the ice surface (T. Irvine-Fynn for scale).
Research to date has almost invariably assumed circular hole planforms, presumably to provide a simplified model system for studying the fundamental mechanisms of hole development. It is now well established that cryoconite holes form due to accelerated melting of ice beneath accumulations of cryoconite (Agassiz, 1846; Gribbon, 1979; McIntyre, 1984; Nordenskiöld, 1875; Phillip, 1912; Wharton et al., 1985). This occurs because cryoconite has low albedo relative to the surrounding ice surface, meaning it efficiently absorbs solar radiation. Gribbon (1979) proposed a conceptual model (see equation (1), Table 3) showing the solar radiation received by cryoconite sediment diminishing as it sinks in ice. There is, therefore, a critical depth where the melt beneath cryoconite is equal to that of the surrounding ice surface. This is referred to as the ‘equilibrium depth’. Equilibrium depths vary over space and time according to environmental conditions (particularly the balance between radiative and turbulent heat fluxes in the local ice surface energy budget), the albedo of cryoconite and the extinction coefficient of light (k) defined by Beer’s law. Beer’s law describes the exponential decay of solar radiation (I) as a function of distance (z) through ice (Oke, 1987)

Equations relating to the thermodynamics of cryoconite hole evolution.

Coupling between equilibrium depth and environmental conditions suggests cryoconite hole depth could be a crude indicator of synoptic energy balance (McIntyre, 1984). McIntyre (1984) used polythene screens to shade holes and observed dramatic changes to hole shapes, confirming the role of direct solar irradiance as a primary control on hole morphology. Microbial activity had previously been suggested to contribute significantly to the heat balance at the hole floor (Gerdel and Drouet, 1960) but this was discredited by the work of McIntyre (1984), Fogg (1988), Gribbon (1979) and Wharton et al. (1985). Recent work by Hollesen et al. (2015) has shown that bioheat in organic rich permafrost can accelerate ice-melt, but this is unlikely to occur in cryoconite unless it is extremely rich in organic matter or present in thick deposits. There is evidence that the melt water in cryoconite holes might influence hole evolution by acting as a radiation filter, sink of latent heat and medium for convection and advection of heat away from hole floors (McIntyre, 1984). Sinking dark cryoconite beneath a layer of reflective water may also raise the albedo of the ice surface, altering the local energy balance regime (McIntyre, 1984). Hydraulic processes might therefore influence hole morphology, but this has not yet been examined in detail.
Podgorny and Grenfell (1996) provided an analytical model of radiation absorption by cryoconite that demonstrated the importance of solar radiation and sediment albedo; however, it dealt specifically with cryoconite holes developing on the floors of melt pools on sea ice and is probably not directly applicable to holes on terrestrial ice. Similarly, a model for shortwave radiation-driven cryoconite hole formation was produced by Jepsen et al. (2010) and supported by field and laboratory experiments; however, this model considers closed holes in perennial lake ice and omitted atmospheric exchanges of sensible and latent heat, limiting its application to cryoconite holes on Antarctic lakes. No solvable model for cryoconite hole depth evolution on Arctic ice currently exists.
Most studies of cryoconite hole evolution have assumed that heat fluxes are exclusively vertical, although we now know that is not the case. Cook et al. (2010) proposed a process of horizontal evolution in response to sediment supply. Horizontal heat fluxes through thickened layers of cryoconite sediment promote melting of hole walls, causing the hole to expand at the base and the holes walls to overhang. Cryoconite can then fall into the newly created space at the hole periphery and the sediment layer thins. A new equilibrium state is attained when cryoconite is spread into a layer just one grain thick (single grain layer, ‘SGL’) and the widening at the hole floor has propagated upwards due to surface ablation, producing a hole with straight walls and a wide aperture. The precise mechanism of granule redistribution across the expanded hole floor remains uncertain, but recent observations (Cook, unpublished data) suggest the primary mechanism is probably gravity-driven sliding of cryoconite down sloped hole floors due to uneven solar irradiance, while redistribution driven by air bubbles escaping from the melting hole floor may contribute (Cook et al., 2010) in areas where cryoconite granules are small.
Cryoconite holes can therefore evolve in three dimensions in response to changes in the local ice surface energy budget and sediment supply. They tend towards equilibrium morphologies characterised by maximal areal coverage and exposure of granules to solar irradiance, which promotes photosynthesis (Cook et al., 2010). These equilibrium states are common in cryoconite holes in slow moving, low gradient ice that is free from topographic shading during periods of shortwave radiation dominated surface energy balance (Cook et al., 2010; 2012; Hodson et al., 2010a), although in steep, hummocky ice and in areas where rates of sediment delivery are particularly high or during periods of turbulent heat flux-dominated surface energy balance, thicker sediment layers exist. This implies that although the process of sediment layer equilibration might be ubiquitous in cryoconite holes, the attainment of SGL can sometimes be inhibited by synoptic conditions. The fundamental processes of vertical and lateral equilibration of cryoconite holes can be summarised in a conceptual model (Figure 4) wherein thermal energy absorbed by cryoconite debris is directed to the hole floor (QV) or transferred laterally for melting hole walls (QL) in varying proportions according to the thickness of the debris layer (equations (5) and (6), Table 3).

Flow diagram depicting the vertical and horizontal development of cryoconite holes. The term ‘I*’ refers to solar radiation and ‘SGL’ stands for single grain layer. Ice is depicted using light grey shading and water is depicted using diagonal hatching.
There are several specific challenges that must be overcome before a solvable model of cryoconite hole evolution can be developed. Firstly, the mechanism of sediment redistribution on hole floors must be better constrained. Second, a much deeper understanding of the thermodynamic processes operating in cryoconite holes, including in the water column, is required. Furthermore, equations (5) and (6) (Table 3) relate thermal fluxes to ice–debris contact area, but this assumes constant layer thickness and will therefore only be applicable to holes in morphological equilibrium, which are only common on stable, slow moving ice in the interior zones of large glacier and ice sheets. Elsewhere, shallow holes contain thick sediment layers and granules can be dispersed directly upon the ice surface (Hodson et al., 2007, 2008; Irvine-Fynn et al., 2011; Stibal et al., 2008) and understanding these may require a revised approach. Current models are limited to well-developed cryoconite holes and there is a lack of research into hole initiation which likely relies upon complex surface debris dynamics, microscale hydraulics and spatial heterogeneities in surface melt rates (Lancaster, 2002). Finally, these models assume an uninterrupted pathway towards morphological equilibrium, whereas periods of turbulent flux dominated ice surface energy balance can reduce the depths of cryoconite holes, sometimes causing them to melt out and redistribute sediment onto the ice surface (Hodson et al., 2007). Similarly, natural cryoconite systems can be disturbed by rainfall events. Only during extended periods of meteorological stability characterised by high incident radiation receipt can cryoconite holes be expected to evolve as described by the models in Table 3.
2 Cryoconite holes and synoptic energy balance
The complex interplay between synoptic energy balance conditions and cryoconite hole evolution was studied by Hodson et al. (2007) who showed that frequent disruption and emptying of Svalbard cryoconite holes occurred in areas of high melt rate. This was due to hydraulic mobilisation of sediment from shallower holes. Hodson et al. (2010b) and Irvine-Fynn et al. (2011) both used time lapse imagery to show that during sunny periods when radiative fluxes dominate ice surface energy balance, cryoconite holes deepen as predicted by Gribbon (1979). This causes the albedo of the ice surface to increase locally since dark sediment sinks further beneath melt water which, while still dark has greater reflectivity than cryoconite. In contrast, periods of turbulent flux dominated energy balance cause cryoconite holes to shallow, moving dark cryoconite closer to the ice surface and reducing the local ice surface albedo. Tracking of individual cryoconite granules suggested continuous redistribution of cryoconite sediment, usually in random directions and resulting in little net displacement, but occasionally melt water flow caused rapid movement of cryoconite between holes (Irvine-Fynn et al., 2010). Hodson et al. (2010a) further suggested frequent hydraulic redistribution of granules between cryoconite habitats and glacier zones. Patterns of melt-in and melt-out of cryoconite influence the microtopography and therefore roughness of the ice surface, with implications for surface albedo (Cutler and Munro, 1996; Warren et al., 1998) and turbulent heat fluxes (Munro and Davies, 1977). These observations imply that cryoconite hole morphology is not only an indicator but may also be a driver of energy balance across melting ice surfaces. In general, periods of prolonged radiative-flux dominated ice surface energy balance are associated with deeper, more stable cryoconite holes.
3 Feedbacks between cryoconite granules and cryoconite holes
The current model of cryoconite granule formation has been inferred from the relationships between OM and granule morphology, along with observations of granule microstructures and biogeochemistry. Langford et al. (2010) suggested the following mechanism:
Blooms of filamentous and unicellular phototrophs (particularly cyanobacteria) on ice, snow and slush surfaces act as nets for wind-blown debris (Figure 5(a)). During snow and slush melt, debris is strained through these nets, forming biofilms that are deposited on ice surfaces. Autotrophic activity adds sticky extracellular polymers to these nets, enhancing their ability to retain particulates and prevent them from being washed down-glacier in suspension in melt water (Figure 5(b)). As these aggregations grow they become increasingly able to resist entrainment into flowing meltwater and therefore settle in microtopographic lows on ice surfaces.

Scanning electron microscope images of cryoconite from 38 km inland on the Greenland Ice Sheet (near Kangerlussuaq) showing (a) a ‘net’ of cyanobacterial filaments binding mineral fragments; (b) EPS mineral interactions along the sheath of a cyanobacterial filament.
After the granules have settled, cryoconite holes can begin to form. Hole formation is primarily driven by abiotic processes of radiative and turbulent heat fluxes; however, microbial activity alters the albedo of cryoconite granules (Takeuchi, 2002a; Takeuchi et al., 2014; Tedesco et al., 2013). Since albedo controls energy absorption and therefore thermal fluxes through cryoconite granules, accelerated melt could be described as microbially mediated. Once granules reside on cryoconite hole floors, further redistribution is unlikely due to the low competence of hydrologic flow through the weathering crust, unless heterogeneous irradiance causes hole floors to slope (Cook et al., 2015). Hydrological monitoring by Cook et al. (2015) also suggests that even if granules are entrained into flowing meltwater, granules are unlikely to be evacuated onto the ice surface. Integrating this information with the well-known association between stability and extended periods of net carbon fixation on Arctic ice surfaces (Anesio et al., 2009; Cook et al., 2012; Hodson et al., 2007) and the tendency for cryoconite holes to widen in favour of autotrophy (Cook et al., 2010) suggests that photosynthesis-driven granule growth creates and maintains favourable conditions for further photosynthesis on Arctic ice surfaces. This in turn drives further granule growth and stabilisation (Langford et al., 2010, 2014). This feedback mechanism is not currently well-understood, but it does point towards the high ecological significance of cryoconite autotrophs, possibly representing keystone taxa (Paine, 1969) or autogenic ecosystem engineers (Jones et al., 1994).
In summary, cryoconite granule formation is ultimately driven by biological processes and results in the formation of cryoconite holes, which further promote biological activity. However, this explanation is a great oversimplification that cannot explain the complex associations between the changeable abiotic environment, biological processes and hole morphology. Complex coupling exists between cryoconite surface coverage, albedo, microtopography and melt processes. These processes may have implications for the process of glacier and ice sheet wastage and require further investigation. What is abundantly clear is that cryoconite holes should be viewed as diverse, dynamic, responsive entities on ice surfaces.
IV Cryoconite biology
1 Microbes in cryoconite
Cryoconite is an important microbial habitat and a major component of supraglacial ecosystems (Anesio and Laybourn-Parry, 2011). Although recognition of this is commonly credited to Hodson et al. (2008), Kohshima (1984a) identified a complex microbial community in ice and snow around cryoconite holes twenty-four years earlier. Kohshima’s subsequent work in the Himalaya, Tien Shan and Andes (Kohshima, 1984b, 1985, 1987a, 1987b, 1989) described abundant and diverse microbiota and meiofauna and linked them with accelerated ablation via ice surface albedo reduction. Takeuchi et al. (1998) later explicitly linked algal growth on a Himalayan glacier to summer mass balance. Kohshima and Takeuchi’s work during this period provided a crucial basis for developing our understanding of microbially-mediated glacier wastage and recognition of supraglacial microbial ecology; Kohshima et al. (1993: 1) stated: “Himalayan glaciers are never abiotic environments. They are simple and well closed ecosystems; housing various microbes, insects and copepods”, cementing a new paradigm of glacier ecology that recognised the diversity and significance of life on ice.
In the late twentieth century, Japanese scientists made advances in the microbiology of the supraglacial environment, while European and American scientists focussed specifically upon cryoconite. Wharton et al. (1981) broke a 20 year hiatus in cryoconite ecology research, identifying cryoconite holes as microbial niches. Wharton et al. (1985) further described them as discrete ecosystems with “distinct boundaries, energy flow and nutrient cycling”, establishing an ecological context with an inherent reductionism that persists to the present day. Vincent (1988) further suggested that cryoconite ecosystems were more complex than other glacial habitats. Despite this work, an assumption of lifelessness on ice surfaces permeated cryosphere literature until at least the late 1980s with writers such as Pyne (1986) stating that there was “no terrestrial ecosystem… in Antarctica”. It was a further twenty-five years before Anesio and Laybourn-Parry (2011) proposed the cryosphere to be one of the earth’s major biomes, with cryoconite holes representing important sites of concentrated microbial activity and biodiversity. Globally, between 1021 and 1026 cells have been estimated to be contained within the porous near surface ice that provides the substrate for cryoconite hole formation (Irvine-Fynn and Edwards, 2014).
Within cryoconite holes, biota is found both in cryoconite granules and the overlying meltwater (e.g. Säwstrom et al., 2002), with by far the greatest microbial abundance in the granules (Anesio et al., 2009; Mieczen et al., 2013). Anesio et al. (2009) found the concentration of bacteria in cryoconite meltwater to be ∼300× less than in debris. Nevertheless, microbes in melt water may represent important contributors to the supraglacial ecosystem. Stratification of ciliates in the water column in cryoconite holes was identified by Mieczen et al. (2013) in accordance with temperature and nutrient concentration gradients. Sixteen phyla of ciliate were identified in total with algiverous and mixotrophic species inhabiting the surface layers and bacterivores occupying the deeper water adjacent to bacteria-rich cryoconite sediment. Suspension of microbial cells in melt water flowing through porous near-surface ice called the ‘weathering crust’ (Muller and Keeler, 1969) may provide an important mechanism of mixing and redistributing microbes in the supraglacial zone. This porous ice layer likely permits admixtures of soluble resources to be homogenised and translocated through the surface ice (Irvine-Fynn and Edwards, 2014; Irvine-Fynn et al., 2012). Punctuating this porous ice layer are cryoconite holes, which likely act as longer-term storage units for biomass (Cook et al., 2015; Hodson et al., 2008) and sites of enhanced biogeochemical cycling, probably utilising resources delivered by hydrologic fluxes through the weathering crust.
Cryoconite granules represent by far the most biodiverse supraglacial microbial habitat, harbouring communities of several trophic levels. Food webs in cryoconite are underpinned by photoautotrophy: primary producers use mainly solar energy to fix atmospheric CO2 into OM, providing substrate for heterotrophs. Of the primary producers, cyanobacteria often dominate both the biomass and the C fixation in cryoconite holes (Christner et al., 2003; Hodson et al., 2010; Mueller et al., 2001; Porazinska et al., 2004; Stibal and Tranter, 2007; Stibal et al., 2006; Takeuchi et al., 2001b; Zarsky et al., 2013) while simultaneously promoting the growth of granules by entangling minerals and additional OM. Heterotrophs metabolise OM both fixed by primary producers in situ (autochthonous) and delivered from elsewhere (allochthonous). Heterotrophic communities in cryoconite are often diverse and usually dominated by a wide range of bacteria. This has been demonstrated using culture-dependent studies (e.g. Margesin et al. 2002) and clone libraries (e.g. Cameron et al., 2012; Edwards et al., 2011). In Svalbard, at least seven phyla of heterotrophic bacteria were identified in clone libraries (Edwards et al., 2011), with their abundance varying between different glaciers. Sampling of Arctic, Antarctic and Alpine cryoconite by Anesio et al. (2010) showed bacterial abundances ranging from 0.05 × 109 cells g−1 in Antarctica to 1.40 × 109 cells g−1 in Svalbard. Rates of bacterial carbon production were not correlated to abundance and varied greatly across the glaciers. In addition to bacteria, higher heterotrophs also inhabit cryoconite holes, surviving by grazing upon smaller organisms. These include tardigrades, rotifers, copepods, ice worms and midge larvae (e.g. DeSmet and Van Rompu, 1994; Zawierucha et al. 2013). Several taxa of tardigrada and rotifera in Spitsbergen cryoconite holes were characterised by DeSmet (1988; 1990; 1993), DeSmet et al. (1988) and DeSmet and Van Rompu (1994). Ciliates usually represent the most complex protozoa in cryoconite holes and are crucial for nutrient recycling through metabolism of primary producers (Mieczen et al., 2013). To date, only a few studies have isolated yeasts and fungi in cryoconite (Edwards et al., 2013b; Margesin and Fell, 2008; Singh and Singh, 2012) and only Edwards et al. (2013b) evaluated their spatial variability, finding distinct communities on adjacent glaciers and suggesting ice surfaces could represent reservoirs of fungal diversity.
Viruses are prominent in cryoconite holes (Anesio et al., 2007; Bellas and Anesio, 2013; Bellas et al., 2013; Hodson et al., 2008; Säwstrom et al., 2007) and could be considered ‘predators’ exerting top-down controls on bacterial populations since they infect and cause the death of bacteria through viral lysis. Viral lysis is crucial for bacterial mortality and thereby influences the recycling and export of carbon and nutrients in cryoconite holes. Viral abundances of about 0.6 × 106 ml−1 and 20 × 106 ml−1 in water and sediment have been identified (Anesio et al., 2007) in Svalbard. The frequency of virus-infected cells in cryoconite communities (13%) was shown by Säwstrom et al. (2007) to exceed temperate freshwater ecosystems (2%). Most recently, Bellas et al. (2015) analysed viral genomes from Arctic cryoconite metaviromes revealed genomic signatures of unusual life strategies which are thought necessary for longer-term interaction with their hosts. Viral shunts represent a crucial mechanism by which DOC is recycled or made labile in cryoconite holes, thus affecting the flow of resources to higher trophic levels and truncating the cryoconite food web (Laybourn-Parry et al. 2001).
Similarities in community structures in polar cryoconite have been illustrated using 16 S rRNA gene clone libraries (Cameron et al., 2012), implying a degree of cosmopolitanism in cryoconite microbial assemblages. However, the abundance of particular species has been shown to vary locally (Cameron et al., 2012), suggesting adaptation to environmental conditions at the community level. Stibal et al. (2008) used epifluorescence microscopy to determine the abundance of phototrophs (0.25 × 103 to 8.0 × 103 cells mg−1) and heterotrophs (10 × 103 to 50 × 103 cells mg−1) in cryoconite from Werenskioldbreen (Svalbard), finding much more biodiversity in heterotrophic communities than the cyanobacteria-dominated autotrophic communities (further described by Stibal et al. 2006; Stibal and Tranter, 2007). At lower latitudes and on some small polar glaciers, heterotrophs have been suggested to be more abundant than primary producers, and may also be supported by allochthonous OC. For example, Edwards et al. (2013c) found genes associated with proteobacteria and bacteriodetes to be more abundant than those associated with cyanobacteria in a metagenome of cryoconite taken from Rotmoosferner (Austrian Alps). Examining community structures is useful for understanding microbial dynamics on glaciers, but has also been linked to cryoconite granule morphology, perhaps indicative of biotic-abiotic feedbacks between microhabitat structure and microbial ecology. For example, Takeuchi et al. (2001c) described poorly aggregated, fine-grained cryoconite in Patagonia where both autochthonous and allochthonous OM were relatively scarce, limiting granule building efficacy. In contrast, cryoconite in the Tien Shan had more abundant biota and higher OM fractions and consequently formed larger, more stable granules.
While cryoconite hole biota can be categorised into distinct trophic levels, functionality is often shared by various organisms throughout the community. For example, granule growth results from the production of cyanobacterial biomass, but also through humification of OM and production of EPS by heterotrophic bacteria, fungi and yeasts. Biogeochemistry is mediated by several organisms including nitrogen fixing bacteria and those involved in ammonification and nitrogenation, including ammonia-oxidising Archaea (Zarsky et al., 2013). Biological darkening of cryoconite granules (and the wider ice surface) is carried out by organisms including algae and bacteria whose activity results in the accumulation of dark humic material and photo-protective pigments (Quesada et al., 1999; Takeuchi, 2002a; Takeuchi et al., 2010, 2014; Tedesco et al., 2013).
To date, microbiological and molecular studies on glacier surfaces have been limited to snapshots, and we await insight into temporal shifts and environmental responses in cryoconite at the community level. However, molecular analyses are becoming increasingly affordable and accessible and are consequently being employed more frequently. It is becoming increasingly clear that close examination of ecosystem structure and function will be crucial for understanding biotic–abiotic processes on glacier surfaces. For a more comprehensive review of the microbes inhabiting cryoconite holes, we direct the reader towards Kaczmarek et al. (2015).
2 The origins of cryoconite biota
The origins of cryoconite biota are currently unclear, with robust evidence in favour of any dominant source or mode of delivery currently lacking. However, several hypotheses have emerged from the literature, mostly linking cryoconite biota to the aeolian biome. Swan (1992) proposed the existence of an aeolian biome based upon observations of microalgae, bacteria and spiders on high altitude and polar ice. Active microbes have since been confirmed to survive in suspension in the atmosphere, some of which are eventually deposited on ice surfaces (Pearce et al., 2009; Sattler et al., 2001; Swan, 1992). There, cryo-tolerant species survive, especially under favourable conditions in cryoconite holes, whereas others perish due to environmental and competitive stresses (Anesio and Laybourn-Parry, 2011; Pearce et al., 2009). Cryoconite biota may therefore be delivered to glacier surfaces directly from the atmosphere via both wet and dry deposition. This may be from local or distant sources. Alternatively, biota may be deposited onto fresh snow packs or precipitated out of atmospheric suspension in snow fall (probably providing nuclei for ice crystallisation; Edwards et al., 2014a) and either redistributed hydrologically following snow melt or incorporated into glacier ice in the accumulation zone. Microbes interred englacially may be stored for 102–105 years, some being preferentially partitioned into water veins (Mader, 1992) before being melted out in the ablation zone. Biota in basal ice and sediment may be extruded onto the ice surface by thrust faulting or via pressure ridges and subsequently blown upglacier. There may also be biota blown or simply dropped onto glaciers from local valley sides.
Suspension of microbes in the high atmosphere may pre-select microbial communities for survival on glacier surfaces. Evidence suggests long term survival in the atmosphere requires similar adaptation to survival on ice surfaces (tolerance to UV, cold and desiccation; Pearce et al., 2009) and viable communities may therefore be shaped to some degree prior to deposition. However, there are commonalities in species abundance between cryoconite holes, soils, freshwater, marine, frozen lake and activated sludge habitats (Edwards et al., 2011) that indicate wide cosmopolitanism of many cryoconite microbes. Furthermore, post-depositional structuring of communities has been suggested to be dynamic over short time scales, responding rapidly to local biotic and abiotic stresses (Edwards et al., 2014b; Priscu and Christner, 2004). There is most likely a fine balance between cosmopolitanism and endemism that reflects complex associations of sources and modes of delivery of biota, post-depositional ecological dynamics and local environmental regimes that collectively shape cryoconite microbial communities. Amplicon pyrosequencing of cryoconite bacterial 16 S rRNA genes from six Arctic and Alpine glaciers and the southwestern margin of the Greenland Ice Sheet (Edwards et al., 2014b) reveals that cosmopolitan generalist taxa (cf. Barberan et al. 2012) are predominant in cryoconite bacterial communities (Figure 6). This is consistent with the notion that priority effects may be important in colonization of nascent cryoconite granules by bacteria, first indicated by Edwards et al. (2013c) as a result of phylotype abundance distribution models supporting deterministic community assembly as a result of several stages of succession. To an extent, environmental variability may buffer against priority effects (cf. Tucker & Fukami, 2014), promoting species richness and evenness as insurance against fluctuating environmental conditions. Indeed, varying levels of bacterial community evenness correlated with ecosystem function measurements in Svalbard cryoconite (Edwards et al., 2011). By these means, perturbations at the ice surface may trigger reconfiguration of cryoconite associated microbiota. Further support that cryoconite granules are formed as a result of microbial succession is provided by the striking negative correlation in the relative abundance of taxa assigned to r-selected, early-colonizing Betaproteobacteria versus typically K-selected, late-colonizing Alphaproteobacteria (Figure 6; Edwards et al., 2014b). It is clear that the processes of microbial succession in supraglacial habitats such as cryoconite not only require, but merit further study as currently, our understanding of these processes is basic yet the implications for the development of biodiverse communities as a result of glacier melting are considerable. In short, the question of whether microbial succession interacts with glacier atrophy prior to the development of the glacier forefield is raised.
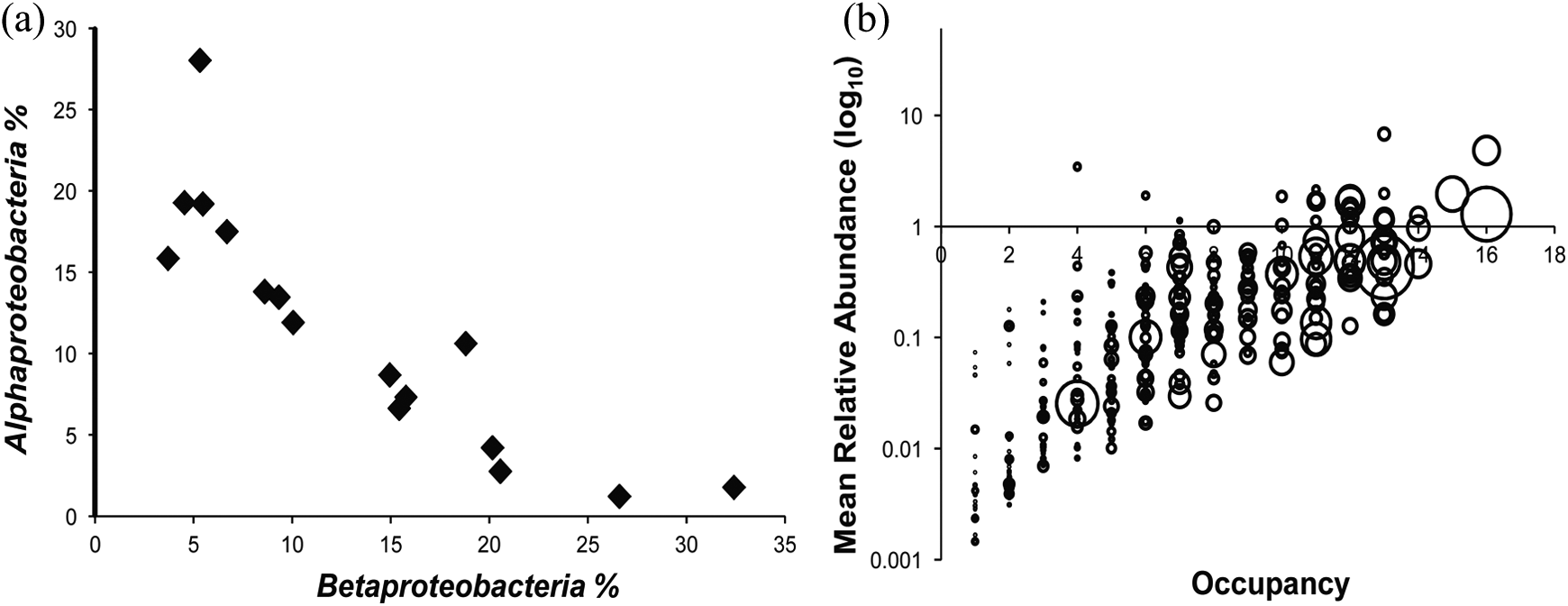
Population trends in Arctic and alpine cryoconite bacterial communities revealed by 16S rRNA gene amplicon pyrosequencing (Edwards et al., 2014b). (a) A negative correlation between the relative abundance of 97% id OTUs assigned by ribosomal database project taxonomy to Alphaproteobacteria and Betaproteobacteria classes is clearly apparent (Pearson’s r = −0.88, p ≤ 0.0001). (b) Dominance of ‘generalist’ OTUs sensu Barberan et al. (2012). Bubble size is proportional to cumulative relative abundance of each Operational Taxonomic Unit. Raw data are available at EBI-SRA (PRJEB5067-ERP004426).
3 Cryoconite biogeochemistry
Biogeochemical cycling in cryoconite holes has been intensely researched over the past decade due to the potential for storage, transformation and export of nutrients. Cryoconite ecosystems have been found to be co-limited by temperature and nutrient stress (Säwstrom et al., 2002), meaning knowledge of hole biogeochemistry is crucial for understanding cryoconite hole microbiology. Research has concentrated upon three main elements: carbon (C), nitrogen (N) and phosphorus (P), with a particular focus upon C. These nutrients will be studied individually here.
Carbon
Cryoconite holes represent particularly active sites of C fixation (autotrophy) and oxidation (heterotrophy) in the supraglacial zone (Hodson et al., 2010a, 2010b), with rates comparable to warm, nutrient rich environments being measured in cryoconite (Anesio et al., 2009, 2010; Hodson et al., 2007, 2008). C transformations may determine glacier albedo through aggregation and darkening of cryoconite (Bøggild et al., 2010; Hodson et al., 2010a, 2010b; Kohshima et al. 1993; Takeuchi, 2009; Takeuchi et al., 2001a, 2001b, 2001c) and influence the quality and quantity of C exported to extraglacial environments (e.g. Hood et al. 2009).
Net ecosystem production
Net ecosystem production (NEP) describes the balance between autotrophy (fixation of CO2 into organic molecules) and heterotrophy (metabolism of organic molecules back into atmospheric CO2):

This has been a particular focus for glacier microbiologists because NEP determines whether a community represents a C sink or a C source (e.g. Hodson et al., 2010b; Stibal et al., 2008a), and also whether there is an overall increase in dark OM on a glacier surface. In cryoconite holes autotrophy is opposed by heterotrophy, perhaps providing biotic control on granule morphology and glacier ablation (e.g. Hodson et al., 2008). NEP both drives and indicates community structure and function and hole biogeochemistry. For example, C fixation increases the pH of cryoconite hole melt water (Stibal and Tranter, 2007; Stibal et al. 2010), particularly important in Antarctic systems where cryoconite holes are decoupled from surface and atmospheric exchanges by thick ice lids. Several crucial abiotic controls upon NEP have been identified, including sediment arrangement (Cook et al., 2010; Telling et al., 2012a), PAR, N and P availability, solar angle and hydrologic regime (e.g. Hodson et al., 2007, 2010b; Irvine-Fynn et al., 2011; Mindl et al., 2007; Stibal et al., 2008b, 2012b; Telling et al., 2012a). Cook et al. (2010) and Telling et al. (2012a) suggested a threshold thickness of 3 mm above which systems are net heterotrophic, below which net autotrophic, and at which NEP is balanced. Telling et al. (2012a) found sediment thickness to explain more than half the variation in NEP in Arctic cryoconite due to the increased surface area of cryoconite exposed to PAR. Cook et al.’s (2010) mechanism of lateral equilibration suggests that given low gradient, slow moving ice and moderate sediment delivery, cryoconite holes evolve towards robust net autotrophy. Where ice surface are steeper or faster moving thick, net heterotrophic layers are more likely to form. Therefore, although rates of microbial activity in cryoconite holes are broadly similar to aquatic ecosystems (e.g. Anesio et al., 2009) NEP varies considerably over both space and time (Table 4). This has been further corroborated by various Greenland (Cook et al., 2012; Stibal et al., 2010, 2012b; Yallop et al., 2012) and Svalbard (Langford et al., 2014) transect studies which have found spatial patterns of NEP controlled primarily by granule size limits imposed by disaggregation and removal of biomass by melt water, while nutrient and PAR availability are also important. Under seasonal snow, photosynthesis probably ceases and systems become net heterotrophic; however, this remains untested. Outside of cryoconite holes, thick, net heterotrophic accumulations of cryoconite can develop under distinct redox and hydrological conditions (Hodson et al., 2008).
Previously reported rates of PP, R and NEP in debris in natural cryoconite holes. The methods used to derive these data are shown: DO2 refers to changes in dissolved oxygen concentration, 14C refers to incorporation of traceable radioisotopes of C and ΔTDIC refers to changes in dissolved inorganic C concentration. ‘n.d.’ refers to values that were not reported. A and H refer to net autotrophy and net heterotrophy respectively.
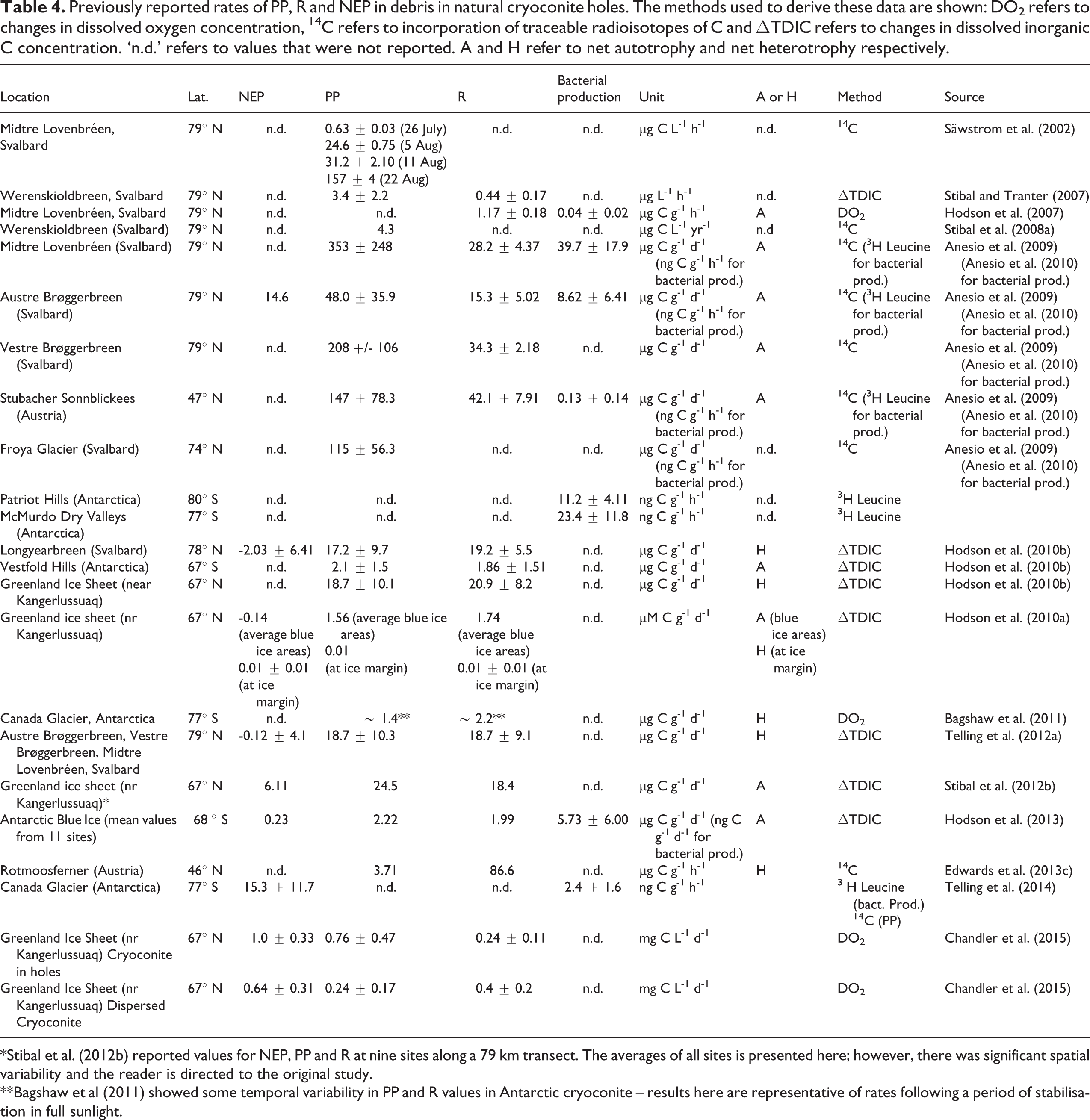
*Stibal et al. (2012b) reported values for NEP, PP and R at nine sites along a 79 km transect. The averages of all sites is presented here; however, there was significant spatial variability and the reader is directed to the original study.
**Bagshaw et al (2011) showed some temporal variability in PP and R values in Antarctic cryoconite – results here are representative of rates following a period of stabilisation in full sunlight.
NEP in cryoconite holes is impacted by bacterial activity (Foreman et al., 2007; Hodson et al., 2007; Mindl et al., 2007) predominantly via heterotrophic C oxidation, much of the energy from which drives bacterial growth and production. Anesio et al. (2010) examined bacterial production in Antarctic, Arctic and Alpine cryoconite, finding rates between 0.13 ng C g−1 h−1 (Stubacher Sonnblickees, Austria) and 39.7 ng C g−1 h−1 (Midtre Lovenbreen, Svalbard). The same study found bacterial doubling times to usually be <5 days (where >60 days was suggested to signify negligible contribution to cryoconite biogeochemistry) apart from in Antarctic melt water and radioactive cryoconite on Stubacher Sonnblickees (Austria). However, bacterial production only represents transformation of OC into biomass, omitting growth and respiration (Anesio et al., 2010; Hodson et al., 2007). To characterise bacterial C fluxes, bacterial growth efficiency (BGE) and respiration rates are also required (Hodson et al., 2007). Anesio et al. (2010) estimated bacteria in Arctic cryoconite to use only 0.4–2.4% of available OC (1.2–7% in Antarctica), suggesting autochthonous production alone was more than sufficient to sustain bacterial production. Low rates of production were therefore unlikely to be due to OC limitation. Temperature, P, viral lysis, and grazing of bacteria by ciliates were identified as much more likely limiting factors, although on Stubacher Sonnblickees, rates were probably also limited by radioactivity (∼140,000 Bq) persisting from the 1984 Chernobyl disaster and bomb tests in the 1950s and 1960s (Tieber et al., 2009).
Recognising the need for standardised reporting of NEP, PP and R, Telling et al. (2010) evaluated various measurement techniques. Measuring changes in dissolved inorganic C (ΔTDIC) consistently outperformed radiolabel incorporation and dissolved O2 methods. A standard procedure was proposed by Hodson et al. (2010b) and Telling et al. (2010) whereby ΔTDIC incubations last for whole days and results normalised for dry mass. Rates of PP, R and NEP previously reported in the literature are shown in Table 4.
Allochthonous and autochthonous carbon
Cryoconite microbes utilise OC synthesised in situ (autochthonous) and delivered from elsewhere (allochthonous). Autochthonous production involves fixation of atmospheric CO2 into organic molecules predominantly by photosynthesis. Allochthonous OC is primarily deposited by wind and comprises fragments of flora, mineral dusts, microbes, BC and anthropogenic pollutants from local and distant sources (e.g. Hodson, 2014; Stibal et al., 2012b; Swan, 1992). Englacial OC can be released by melt out and provides a significant source of ancient bioavailable OC for downstream ecosystems (Stubbins et al., 2012) and likely provides an energy source for cryoconite microbes. Subglacial OC might also be delivered to the supraglacial zone by thrust faulting (Stibal et al., 2012b), supplementing allochthonous OC in cryoconite holes in some locations.
Autochthonous and allochthonous OC together (TOC) provide crucial substrates for heterotrophy. It is uncertain whether OC demands in cryoconite holes are met by autochthonous production, or whether heterotrophy is sustained primarily by OC from allochthonous sources (Edwards et al., 2013b; Stibal et al. 2008a, 2010; Telling et al., 2012a). It has emerged that in general, the interiors of large ice sheets and glaciers are characterised by autochthonous OC accumulation and allochthonous OC from distant sources whereas ice sheet margins and small glaciers are dominated by local allochthonous OC (Stibal et al., 2012a; Telling et al., 2012a). However, variations in topographic shading, incident radiation, sediment layer thicknesses, allochthonous OC and nutrient supply and ice type likely establish local heterogeneity and dynamism in cryoconite carbon budgets. For further information on allochthonous and autochthonous OC dynamics we suggest Stibal et al.’s (2012a) review.
Carbon export
OC produced or transformed in cryoconite holes can be redistributed by meltwater and ultimately provide a source of labile C for nearby terrestrial, marine or lacustrine ecosystems (Hood et al., 2009; Lawson et al., 2014; Swan, 1992; Wharton et al., 1985). This is usually by entrainment into supraglacial streams or possibly transport in solution through the low density ice comprising the weathering crust (top ∼2 m of ice surface; Irvine-Fynn and Edwards, 2014). Whole cells and organic molecules dissolved in water (dissolved organic carbon: DOC) are probably transferred between habitats within the supraglacial zone and other glacial and extra-glacial environments, although no measurements of either dissolved or particulate OC exported from cryoconite holes have yet been made. This pathway might be important for seeding proglacial streams, glacier fed lakes and subglacial sediments, as well as providing nutrients and energy sources for pre-existing downstream ecosystems, and requires research, although the stark contrast in microbial abundance between cryoconite sediment and overlying meltwater suggests robust binding of OM to cryoconite debris and limited transfer into suspension or solution. This indicates that cryoconite currently provides POC storage in the supraglacial environment. Nevertheless, cryoconite and other supraglacial habitats have recently been suggested to be important metacommunities influencing downstream biodiversity and community structure (Wilhelm et al., 2013). For further information regarding DOC and POC export to rivers and oceans, we suggest consulting Stibal et al. 2012’s review.
Carbon cycle models
In situ measurements of NEP, PP and R in cryoconite holes have informed models of regional and global C cycling. Anesio et al. (2009) upscaled local measurements of NEP to all glaciated areas outside of Antarctica, suggesting net C fixation of ∼64 Gg a−1. Their analysis assumed rates of NEP in cryoconite obtained from a small number of sites in Svalbard, Greenland and European Alps were globally representative and temporally constant. Further, the extent of the summer ablation zone was assumed constant, compensated for using a conservative melt duration. This approach overlooks seasonal shifts in irradiance, temperature, ice and sediment dynamics that may significantly impact rates of C cycling. Furthermore, Anesio et al.’s (2009) spot measurements of NEP were 10–100 times greater than subsequent measurements made on some of the same glaciers, possibly due to methodological issues (Anesio et al. 2010). Hodson et al. (2010a) estimated C fluxes on the Greenland ice sheet based upon cryoconite alone. Spatio-temporal evolution of the ablation zone was incorporated, where PP was limited to the bare ice area while R occurred in both bare ice and slush. Mass per unit area of cryoconite and rates of PP and R (derived from short transects from the ice margin near Kangerlussuaq, Greenland) were assumed spatially uniform. Later, Cook et al. (2012) showed the assumption of spatial uniformity was not justified, and in fact the greatest mass loading, spatial coverage and rates of biological activity for cryoconite occurred further inland than the end of Hodson et al.’s (2010a) transects. Furthermore, Hodson et al. (2010a) measured late-season communities approaching acquiescence. Cook et al. (2012) obtained measurements from a 79 km transect (the entire ablation zone) to estimate C fluxes from a 1600 km2 area of the Greenland ice sheet, suggesting additional transects would be required to upscale further. Their model incorporated spatially variable coverage, mass loading and rates of PP and R, as well as including surface algal blooms. Spatial variability in cryoconite distribution and rates of activity had a strong influence on C-flux estimates, and the inclusion of surface-dwelling algae potentially greatly increased C fixation along the transect. However, that model is fundamentally limited in a number of ways: First, there remains an assumption that there are only two discrete ecosystems that contribute to supraglacial C fluxes; Second, it assumes microbial activity, spatial coverage and mass loading are constant throughout a melt season and that there is no redistribution of debris or biomass. Third, it assumes no changes in albedo, hydrology or other abiotic factors occur during the melt season and makes no attempt to model any associated feedbacks. Fourth, rates of algal productivity were based upon 14C incubations and loosely constrained estimates of algal respiration. Finally, deriving cryoconite distribution in photographs by calibrating against measurements made in small quadrats is unlikely to be accurate (recently corroborated by Chandler et al., 2015, who found high spatial and temporal variability in cryoconite coverage and productivity on the Greenland Ice Sheet). Nevertheless, these papers encouraged appreciation of the supraglacial biome and its significance for global atmospheric exchanges.
Nitrogen
Despite Bayley (1891) identifying nitrogenous material on ice surfaces, and abundant studies of N cycling in polar oceans (Dittmar, 2004), ice cores (Olivier et al., 2006), soils (Nordin et al., 2004), rivers (Tockner et al. 2002), subglacial sediments (Wynn et al., 2007) and snowpacks (Hodson, 2006; Hodson et al., 2005; Jones, 1999; Williams et al., 1996), supraglacial N cycling was only recently considered in detail. Tranter et al. (2004) found high inorganic: organic N ratios in Antarctic cryoconite holes, implying recycling rather than fixation of nitrogenous compounds, likely due to isolation by thick ice lids. In contrast, holes in Svalbard showed net NH3 production, indicating fixation of N2 by cyanobacteria (Hodson et al. 2008). Telling et al. (2012b) showed N fixation to be energy demanding and only undertaken by N fixers when allochthonous bio-available N is scarce. Therefore, active fixation indicates insufficient allochthonous bio-available N and may limit growth. Säwstrom et al. (2002) showed Svalbard cryoconite communities were not N limited, highlighting the influence of N-fixing bacteria on ecosystem function. Telling et al. (2011, 2012b) investigated N cycling across the ablation zones of Svalbard glaciers and the Greenland ice sheet. N2 fixation was active in cryoconite holes in both regions, implying insufficient allochthonous bio-available N to sustain microbial production. Evidence of denitrification and ammonification was also identified in Greenland (Telling et al., 2011) and NifhH genes have been detected across the south-west Greenland Ice Sheet (Telling et al., 2012b). Allochthonous sources included anthropogenic N compounds and guano from bird colonies (guano was corroborated as a source of bioavailable N by Hodson et al., 2006, 2008; Zarsky et al., 2013). Other stages of N cycling including microbial ammonium assimilation (Zarsky et al., 2013) and nitrification (Ansari et al., 2013; Hodson et al., 2010c; Wynn et al., 2007) have been observed on Svalbard glaciers. Amplification of nifH genes indicated genetic potential for nitrogen fixation by bacteria (Cameron et al., 2012) and Archaea (Zarsky et al., 2013) in Svalbard cryoconite. Catalysis of nitrification (Hodson et al., 2005; Wynn et al., 2007) and N2 fixation (Telling et al., 2011) in cryoconite communities might contribute organic N substrates to subglacial and proglacial ecosystems. The nutrient demands of cryoconite holes and wet snowpacks account for most of the annual NH4 + deposition on Midtre Lovenbréen (Svalbard) (Anesio et al., 2009; Hodson et al., 2005, 2010c; Säwstrom et al., 2002). N-addition experiments have shown N availability not to be a limiting factor in Svalbard cryoconite, suggesting that N-cycling microbes contribute significantly to cryoconite communities (Hodson et al., 2008). Recently, Edwards et al.’s (2013b) cryoconite metagenome showed an abundance of functional genes related to N, Fe, S and P cycling that suggested efficient nitrogen recycling on Rotmoosferner (Austria). Rates of N fixation have been shown to increase in late summer once bioavailable N stores have depleted (Anesio and Laybourn-Parry, 2011). Supraglacial N cycling remains poorly understood, although recent work by Segawa et al. (2014) used isotopic and RNA evidence to directly trace N cycling on a glacier surface in central Asia, showing that intense N cycling supports abundant microbial life on glaciers.
Anthropogenic nitrogen
Allochthonous N deposition down-regulates N2 fixation, increasing available energy for microbial growth and proliferation, although Telling et al. (2011) showed N limitation is unlikely to primarily control biomass production in cryoconite holes. Anthropogenic pollutants have increased the concentration of bioavailable N in polar snowpacks such that cryoconite microbes no longer need to fix atmospheric N2, since activity can be sustained by allochthonous inputs (Telling et al., 2011). Hodson et al. (2010c) monitored nutrient budgets in Svalbard after a period of pollutant deposition and observed shifts in nutrient cycling that impacted the entire glacier catchment. Anthropogenic industrial activity produces nitrogenous compounds that can be suspended in the atmosphere and deposited on glacier surfaces (Kozak et al., 2013), possibly influencing nutrient cycling, community dynamics and NEP in these areas. Heavy metals have also been found to accumulate in cryoconite in the Arctic (Singh et al., 2013). It is well known that in extraglacial areas nitrogenous inputs from industrial emissions, road vehicles and agricultural fertilisers have a strong influence upon ecosystem dynamics.
Phosphorus
Little is known about P cycling in the supraglacial zone, although cryoconite communities are likely P-limited (Hodson et al., 2008; Mindl et al., 2007; Säwstrom et al., 2002; Stibal and Tranter, 2007; Stibal et al., 2008b, 2009). Typical P concentrations on Arctic ice surfaces have been estimated at < 0.1 µM (Stibal et al., 2009). Säwstrom et al. (2002) showed bacterial production in cryoconite to increase following P addition, but only above 12°C, suggesting that cryoconite communities may be co-limited by P availability and temperature. In Svalbard, Stibal et al. (2008b) reported P deficiency in cryoconite melt water relative to cryoconite sediment, indicating that the majority of P in the supraglacial zone was bound to cryoconite debris. Mueller et al. (2001) suggested that P demand likely exceeds P supply in the supraglacial zone because atmospheric deposition is very low and there are no mechanisms of biotic P generation that can compensate. Stibal et al. (2009) agreed that inorganic P is probably scarce and rapidly depleted in cryoconite, but found that microbes respond by recycling P from OM using phosphatase (an enzyme used to scavenge P from organic complexes). The efficiency of this process is uncertain. Adding bioavailable P to cryoconite incubations resulted in the inhibition of phosphatase activity, indicating insufficient allochthonous P supply in natural cryoconite systems. Importantly, Stibal et al. (2009) identified several difficulties with measuring P concentrations in cryoconite, in particular only a fraction of the total bioavailable P present seems to be accessible to microbes, meaning basic chemical extraction techniques are not suitable for assessing the amount of P available for cryoconite microbes.
V Antarctic and lower-latitude cryoconite
Much of the current knowledge about cryoconite has come from work in the Arctic; however, there is a significant body of literature concerning cryoconite in Antarctica and on low-latitude glaciers. Antarctica is generally much colder and drier than the Arctic, meaning cryoconite phenomena contrast greatly with those described in Section II. Antarctic holes form by upwards melting from entombed sediment in summer (Fountain et al., 2008), retaining a thick ice lid that decouples debris and melt-water from atmospheric and ice surface exchanges. This reduces flushing of cryoconite holes and facilitates the accumulation of solutes leached from surrounding ice and produced by microbial activity. Concentrations of Cl- ions in Antarctic cryoconite holes have suggested continuous periods of isolation from atmospheric exchanged of up to 11 years (Bagshaw et al., 2007; Fountain et al., 2004, Tranter et al., 2004). This can result in extreme hydrochemical conditions within cryoconite holes that are unique to Antarctica (Hodson et al., 2008; Tranter et al., 2004). This probably explains the contrasts in biota between Antarctic and Arctic cryoconite. Antarctic cryoconite generally contains little OM (Table 1) and bacterial abundance, productivity and bacterial doubling times have been suggested to be much lower in Antarctic cryoconite than in the Arctic (Anesio et al., 2010). Nutrient limitation arises due to the isolation of Antarctic cryoconite holes, necessitating recycling (Tranter et al., 2004). This increases the concentrations of ammonia, nitrate and dissolved P concentrations and lowers the pH of cryoconite melt water (Mueller et al., 2001) and causes modes of biogeochemical cycling in Antarctic cryoconite holes to contrast with that in the Arctic. In particular, contrasts in nitrogen cycling have been identified between Arctic and Antarctic cryoconite due to inhibited transfer of atmospheric N2 to cyanobacteria in Antarctic holes (Tranter et al., 2004). The OM in Antarctic cryoconite was found to be less heterogeneous than in Arctic cryoconite (Pautler et al., 2013) probably due ice lidding and remoteness limiting allochthonous inputs from plants, mosses and sphagnum from deglaciated land. Similarly, Bagshaw et al. (2013) described clear disparities between cryoconite and valley-floor OC in the McMurdo Dry Valleys, showing cryoconite to support net autochthonous OC production. Antarctic cryoconite is therefore probably more reliant upon autochthonous production of OC to support microbial activity. The different conditions in Arctic and Arctic cryoconite holes has been shown to generate contrasts in microbial community structures. Mueller and Pollard (2004) showed that inter-hole mixing in open Arctic cryoconite holes resets community structures and homogenises ecosystems, whereas hydrologic and atmospheric isolation in Antarctica establishes more optimised communities in areas of different environmental pressures. Microbial communities are less cosmopolitan in the Antarctic because the more extreme environmental stresses promote endemism. An additional cryoconite habitat has been identified in Antarctic blue ice areas (Hodson et al., 2013). These regions of smooth, wind-polished ice contain entombed cryoconite that absorbs solar radiation and melts a subsurface layer of liquid water, supporting microbial activity. The biogeochemistry of these habitats is characterised by low rates of PP, R and melt water replenishment due to low temperature and solar irradiance. These habitats require very low temperatures and persistent strong winds to form and are therefore unique to Antarctica.
Despite being isolated from surface hydrologic and atmospheric exchanges, cryoconite holes are important components of Antarctic glacier hydrology. About a third of all supraglacial melt on Wright Lower glacier was attributed to the formation of cryoconite holes which then stored up to 47,000 m3 of water (MacDonnell and Fitzsimons, 2008). Furthermore, Antarctic cryoconite holes can interconnect and link with supraglacial streams and lakes via discrete conduits (Bagshaw et al., 2007; Fountain et al., 2004, 2008; MacDonnell and Fitzsimons, 2008; Tranter et al., 2004). Up to 56% of cryoconite holes on Canada Glacier were estimated to be hydrologically connected (Fountain et al., 2004), providing a subsurface drainage mechanism that is likely unique to cold Antarctic glaciers. Periodic ‘stripping events’ have also been reported in Antarctica, where the upper layer of glacier surface is suddenly removed by rapid melt (Fountain et al., 2004; MacDonell and Fitzsimons, 2008). This probably provides the primary mechanism of cryoconite redistribution on Antarctic glaciers (Fountain et al., 2004). These events occur roughly decadally on Antarctic glaciers (Bagshaw et al., 2013), whereas they may be annual in the Arctic. Seasonal evolution from hydrologic isolation to interconnectedness was proposed by MacDonnell and Fitzsimons (2012), who also suggested that all cryoconite holes are ultimately erased by ablation, dependent upon water supply, ablation rate and local topography. Telling et al (2014) also showed that periodic melt events provide nutrient fluxes into cryoconite holes that stimulate biological activity including nitrification, phosphorus uptake and bacterial production. Antarctic cryoconite holes have additional importance as analogues for microbial habitats on other icy planets including Mars (e.g. MacClune et al., 2003; McKay and Stoker, 1989; Paerl and Priscu, 1998; Priscu et al., 1998, 1999a, 1999b; Schulze-Makuch and Grinspoon, 2005; Tranter et al., 2010; Wentworth et al., 2005). Understanding microbial processes in Antarctic cryoconite holes may therefore have wider reaching implications than our own planet and contemporary timescale.
Cryoconite on low-latitude mountain glaciers is distinct from cryoconite on polar glaciers and ice sheets. This is primarily due to local glaciological and meteorological conditions. Low-latitude glaciers are generally small, fast flowing and subject to large diurnal fluctuations in energy balance. Solar radiation is more intense during the day at lower latitudes; however dark nights and variable cloud cover can produce complex melt dynamics. Due to the abundance of deglaciated land in close proximity to most low-latitude glaciers there is greater deposition of aeolian dusts and soluble ions from terrestrial and anthropogenic sources. Takeuchi and Li (2008) showed that this causes rapid microbial production and high cryoconite coverage on Asian glaciers. High melt rates on low-latitude glaciers tend to produce shallower cryoconite holes with shorter life spans than those on polar ice (Takeuchi et al., 2000), and more frequent redistribution across ice surfaces might therefore be expected. High microbial production and abundant inorganic impurities encourage diverse microfauna and meiofauna to inhabit the glacier surface. These include glacial midges, copepods, collembolan, tardigrada and rotifers. Contrasting organisational taxonomic units and metabolite profiles has revealed different microbial community structures and functions in cryoconite from Arctic and Alpine glaciers (Edwards et al., 2014b). Cryoconite biogeochemistry on low-latitude glaciers is probably similar to that of ice sheet margins, where fast flowing and rapidly melting ice favours net heterotrophy (Stibal et al., 2012a). Edwards et al. (2014b) suggested a latitudinal control upon NEP whereby lower latitudes are associated with net heterotrophy and high northern latitudes associated with net autotrophy. There is a geographical bias in the literature that favours polar over mountain glaciers, despite mountain glaciers being very sensitive to albedo shifts (Ming et al., 2012; Oerlemans et al., 2009; ), important for water security (Edwards et al., 2014a) and particularly poorly understood in terms of their response to climate change (IPCC, 2013).
There are therefore stark geographical contrasts in cryoconite morphodynamics, biogeochemistry and microbial ecology. However, these seem to arise from variations in the physical environment in which the cryoconite exists rather than from different processes. This suggests commonalities in the fundamental mechanisms governing cryoconite and cryoconite habitat properties that require elucidation, and indicates the potential for general, widely applicable cryoconite models to be developed.
VI Cryoconite, climate change and anthropogenic pollution
The response of Earth’s ice to climate warming can be broadly characterised by increasingly negative mass balance, retreat or acceleration towards lower elevations. Since ice masses store ∼70% of Earth’s freshwater, this threatens to destabilise a crucial domestic, agricultural and industrial resource relied upon by over a billion people (Hodson, 2014) and a crucial biome harbouring unique microbial assemblages (Anesio and Laybourn-Parry, 2011).
Glacier and ice sheet wastage is strongly influenced by ice surface albedo, which may in turn be influenced by supraglacial ecology. Ice albedo can be lowered by algal blooms on ice surfaces (Yallop et al., 2012), dust deposition (Bøggild et al., 2010), melt water production (Gruell, 2000) and also the aggregation of cryoconite granules. Once aggregated the growth and proliferation of microbes, entanglement of allochthonous debris, production of humic substances and photo-protective pigmentation further decrease granule albedo (Hodson et al., 2008, 2010a, 2010b; Irvine-Fynn et al., 2010; Takeuchi, 2002a; Takeuchi et al., 1998, 2001b, 2010; Yallop et al., 2012). Aggregation of cryoconite also enhances the longevity of minerals and OM on ice surfaces, therefore prolonging their albedo lowering effect. For example Hodson et al. (2010b) found cryoconite distribution to explain 53% of the albedo variance on Longyearbreen (Svalbard), although complex interactions between cryoconite coverage, hole evolution, microtopography, surface roughness and energy balance (e.g. Cook et al., 2010; Hodson et al., 2007, 2010b; Irvine-Fynn et al., 2010, 2011) imply complex relationships exist between cryoconite morphodynamics and ice surface melt rate. Increased temperature might cause more frequent melt-out and enhanced hydraulic redistribution of cryoconite over ice surfaces (Hodson et al., 2008) and therefore alter supraglacial melt patterns. As identified by Hodson et al. (2007, 2010a) and Cook et al. (2012), longer ablation seasons due to climate warming will likely extend annual periods of photoautotrophy, possibly promoting growth of cryoconite granules and proliferation of surface dwelling algae, further reducing ice surface albedo. On the Greenland ice sheet, warming was estimated to strengthen net autotrophy and increase atmospheric C fixation (Cook et al., 2012); however, albedo and melt feedbacks were not considered, and increased snowfall in a warmer climate could promote heterotrophy (Jones, 1999). Given recent suggestions of melt-water disruption mobilising cells and partially disaggregating cryoconite granules (Langford et al., 2014), climate-forced changes to glacier hydrologic regimes will likely impact cryoconite morphology and alter environmental stresses on incumbent organisms, with unknown impacts upon ice surface albedo and melt rates. It is currently unclear whether additional melt will enhance nutrient delivery and therefore cohesive OM production promoting cryoconite growth, or enhance disturbance and disaggregation of granules promoting redistribution. In a palaeoglaciological context, Abbot and Pierrehumbert (2010) suggested cryoconite might contribute to albedo-driven deglaciation from ‘Snowball Earth’ events, implying its significance as a melt catalyst over geologic timescales.
As well as being a potential melt catalyst, cryoconite is a store of nutrients and biodiversity in otherwise depauperate or ultra-oligotrophic environments. For example, nutrient concentrations in Antarctic cryoconite holes have been found to be much higher than in surrounding ice and sediments (Bagshaw et al., 2013; Tranter et al., 2004). Since cryoconite holes contribute to supraglacial hydraulic regimes (Fountain et al., 2004; Hodson et al., 2008) the transformation and accumulation of nutrients within them influences the geochemistry of melt waters exported to nearby valley ecosystems via ephemeral streams. Export of nutrients and biota from ice surfaces to deglaciated areas will be sensitive to climatic changes because warmer temperatures ultimately transform glacial zones into proglacial zones (Prowse et al., 2006), accompanied by release of glacier-stored nutrients and changes to mechanisms of nutrient cycling (Stibal et al., 2012b). Glacier hydrologic regimes are vulnerable to dramatic changes due to increased temperature, with mass loss leading to greater isolation of the supraglacial zone due to basal refreezing or, where refreezing does not occur, increased melt could tightly couple the supraglacial, subglacial and extraglacial zones (Hodson et al., 2008), impacting biogeochemical transformations and export to forefields. In the Arctic, Wilhelm et al. (2013) showed that cryoconite contributes to ecological succession in glacier fed streams, particularly following glacier retreat. Cryoconite has also been suggested to be an important source of microbes that recolonize freshly exposed land following glacier retreat (Kaštovská et al., 2005). Increased summer snowmelt might increase the delivery of nutrients and biota to glacier surface habitats and also the extraglacial zones (Hodson et al., 2008). Glaciers have been shown to provide sources of ancient, bioavailable OC to marine, riverine and lacustrine ecosystems and cryoconite may therefore play an important role in defining the carbon cycling regime in diverse environments (e.g. Hood et al., 2009; Mindl et al., 2007; Singer et al., 2012; Stubbins et al., 2012). The role of cryoconite in this process is not currently well understood.
The role of cryoconite granules for contaminant dynamics on glacier surfaces is also uncertain. Hodson et al. (2010b) suggested cryoconite to catalyse supraglacial chemical weathering by extending rock-water interactions, with implications for both glacial and extraglacial geochemistry. Other impurities that influence glacier albedo (e.g. BC, mineral dust) or are ecologically damaging (e.g. POPs, heavy metals, pesticides) may have their residence on glacier surfaces prolonged, and therefore their impact exacerbated, by incorporation into cryoconite. Several types of POP were identified in cryoconite in the European Alps by Lee et al. (2011), particularly on glaciers used for recreation. Genes for antibiotic resistance have been identified in cryoconite in a range of non-Antarctic glaciers, indicating widespread contamination by anthropogenic antibiotics (Segawa et al., 2013). Incorporation into cryoconite might cause bioaccumulation of contaminants. Alternatively, burial of dark particulates under brighter biomass at granule surfaces and metabolism of pollutants might decrease their potency (Hodson, 2014). Potentially harmful radionuclide concentrations have been identified in Alpine cryoconite (Tieber et al., 2009) originating from the 1984 Chernobyl disaster and the bomb tests of the 1950s and 1960s, illustrating the potential for cryoconite contaminant storage. The potential for increased delivery and export of supraglacial contaminants in a warmer climate has not yet been studied in detail. However, the role of supraglacial BC as an amplifier of climatic change has recently been recognised (Casey et al., 2012; Hodson, 2014). There remains great uncertainty about the role of cryoconite in remediating the climate impacts of BC on ice surfaces. Incorporation of BC into cryoconite may prolong its residence, but may reduce its impact by covering it in lighter-coloured biomass. BC may also provide an allochthonous source of bioavailable OC to be metabolised by cryoconite heterotrophs, diminishing its potency (Hodson, 2014). Further studies are required to uncover the role of cryoconite in BC dynamics in the supraglacial zone.
Cryoconite therefore plays a complex and as yet uncertain role in supraglacial climate processes. The properties of cryoconite and cryoconite habitats will likely respond to shifts in climate, while potentially amplifying climate changes through albedo feedbacks. The role of cryoconite in bioremediating or exacerbating the impacts of contaminants and anthropogenic pollutants is still unknown. Furthermore, biogeochemical exports from ice surfaces will likely respond to climate changes and have far-reaching impacts for polar ecosystems. However, these processes are poorly understood and require urgent attention.
VII Is cryoconite science back to where it started?
The history of glacial microbiology could arguably start with the work of Kohshima (1984) and Wharton et al. (1985); certainly appreciation of supraglacial ecosystems has only become widespread since. However, microbial life on ice was hypothesised by ancient Greeks (Sattler et al. 2002) several thousand years previously. In 1818 James Ross described ‘crimson cliffs’ of biologically inoculated ice in Greenland, and soon after cryoconite holes were documented by Agassiz (1846) who suggested that their formation was related to energy balance. The microbiological significance was recognised by Nordenskiöld (1875), and the association between cryoconite and albedo-driven melt dynamics was established by Nordenskiöld (1875) and Nansen (1906). In fact, many of the fundamental principles underpinning modern supraglacial ecology had already been established by the early 1900s, and the major questions facing cryoconite scientists at the end of the nineteenth century were the same as those we face in the second decade of the twenty-first century. It is therefore crucial to look back to the “prehistory” of cryoconite research, both to direct future research and to appreciate the remarkable insights of the early polar scientists.
A.E. Nordenskiöld coined the term ‘cryoconite’ to describe cylindrical ‘ice pipes’ containing ‘remarkable powder’ that hampered his crossing of Greenland in 1870 (Leslie, 1879). The term originates from the Greek ‘kryos’ (cold) and ‘konis’ (dust). The holes he documented were “a foot or two deep, from a couple of lines to a foot in diameter, and so close together that it was impossible to find between them room for the foot, much less a sleeping sack” (Leslie, 1879). Nordenskiöld postulated that these holes formed by solar heating of dark cryoconite debris and this was corroborated by several of his contemporaries, notably Von Drygalski (1897), who further suggested that stable, low gradient ice and multiple melt seasons as prerequisites for hole formation, and subsequently Hobbs (1910), Phillip (1912), Kayser (1928), Wegener (1930) and Poser (1934). Some debate surrounded the relative importance of diffuse and direct radiation for hole development, with Phillip (1912) suggesting direct transmission through hole walls to be the dominant control, whereas Brandt (1931) and Wagner (1938) thought diffuse radiation was more important. Sharp (1947) made early suggestions of lateral expansion of cryoconite holes, presaging Cook et al. (2010) by 63 years, proposing convection currents in cryoconite hole water as a primary driver. Gajda (1958) later summarised cryoconite hole formation that was heavily reliant upon the work of early polar scientists and, although efforts have been made to describe the system numerically and some further insights made since (e.g. Cook et al., 2010; Gribbon, 1979; McIntyre, 1984) remains relevant today.
Observations of cryoconite hole dimensions and frequency distribution have been made regularly between Nordenskiöld (1875) and the present day, and in fact earlier reports were occasionally provided by scientists such as Agassiz (1846). Early data was predominantly gathered in Greenland (e.g. Hobbs, 1910; Nansen, 1906; Nordenskiöld, 1875; Steinbock, 1936; Von Drygalski, 1897). Extensive empirical observations in the ‘cryoconite zone’ (between the ice margin and 700 m elevation) in Thule (Greenland) were provided by Gajda (1958), who found spatial variations in cryoconite hole sizes between the ice margin and snow line. Similar measurements have been made recently, with Stibal et al. (2012b), Yallop et al. (2012) and Cook et al. (2012) examining elevational gradients of cryoconite dimensions in South-West Greenland. The suggestion that cryoconite hole morphology could provide an indicator of synoptic meteorology was proposed by Steinbock (1936) and reiterated by Gajda (1958), Gerdel and Drouet (1960), Brochu (1975) and Gribbon (1979). Recently, cryoconite holes have begun to be recognised as components of the hydrologically active porous 1–2 m of surface ice on glaciers known as the ‘weathering crust’ (Irvine-Fynn and Edwards, 2014; Irvine-Fynn et al., 2012; Muller and Keeler, 1969); however, Gribbon (1979) referenced unpublished work by Thurmann that showed the frequency distribution of cryoconite holes to be dependent primarily upon supply of sediment transported through the weathering crust and temporarily stored in interstitial pores. Gerdel and Drouet (1960) also highlighted weathering crust decay as a key control on hole development, reinforcing work by Von Drygalski (1897).
Upon examining individual granules of cryoconite, Nordenskiöld and other nineteenth century scientists quickly realised they comprised both mineral and biological components. Nordenskiöld showed cryoconite to form “small, round balls of loose consistency” composed mainly of quartz, feldspar, augite and OM as well as mysterious magnetic particles. Nordenskiöld’s botanist revealed the OM to be largely composed of brown-coloured polycellular algae. Around the same time, Swedish explorer N.O. Holst made similar observations, describing cryoconite as “little balls as big as beans” (Lindahl, 1888). On the origin of cryoconite, however, these scientists disagreed. To Nordenskiöld, cryoconite mineralogy indicated wind deposited terrestrial dust from distant sources, supplemented by a cosmic component. Holst, amongst others (including von Lasaule, Zirkel and Swedmark) argued that cryoconite contained “nothing but the ordinary components of primitive rock” (Lindahl, 1888). Petrographic analyses by Bayley (1891) identified feldspar, quartz, mica and hornblende as dominant constituents, with minor contributions from garnet, zircon, magnetite, augite and sillimanite, all thought to have been deposited by wind from a region of crystalline schist’s, although Bayley (1891) also suggested commonalities with deep-sea sediments supporting cosmic contributions. Later analyses by Von Drygalski (1897) and observations of non-uniform cryoconite distribution (Nansen, 1906) largely discredited cosmic origins (Garret and Verzella, 2008). It is possible that some of the particles that Nordenskiöld (1875) identified as cosmic were derived from anthropogenic activity. The industrial revolution was underway but he likely considered Greenland too remote to receive contaminants. It is now known that these pollutants provide bio-available C and N to polar glaciers and darken ice (e.g. Hodson, 2014; Kozak et al., 2013). Later, researchers such as Kornerup and de Quervain suggested cryoconite to be formed from clays or even decomposition products of ptarmigan guano. Guano has been recognised recently as a potentially important N source on some mountain glaciers (Zarsky et al., 2013) and near penguin colonies in Antarctica (Hodson, 2006). As research continued, a consensus that cryoconite comprises terrestrial materials emerged, although sources vary between glaciers (Kayser, 1928). Gajda (1958) and Gerdel and Drouet (1960) highlighted wind deposition from local deglaciated land as a primary source of cryoconite minerals, corroborated by a negative correlation between granule size and distance from the ice margin, supporting Nansen (1906). Although a contemporary debate continues regarding the details of sediment delivery to glaciers (e.g. Bullard et al., 2013; Langford et al., 2014; Tedesco et al., 2013; Zarsky et al., 2013), the early twentieth century consensus that minerals are deposited by wind from local and distant sources, supplemented by inputs from adjacent exposed rock faces and subglacial pressure faulting, persists.
OM in cryoconite holes has been studied intensely during the nineteenth, twentieth and twenty-first centuries. Nordenskiöld and Berggreen had already identified cyanobacteria as the dominant species inhabiting Greenland cryoconite when Whittrock (1885) observed algae and, later, Von Drygalski (1897) reported cyanobacteria, chlorophytes and desmids in nearby samples. Furthermore, Nordenskiöld (1875) identified a near-ubiquitous dust covering the ice between cryoconite holes and identified it as brown coloured polycellular alga that was also present in cryoconite sediment. The same algae interested Nansen (1906), who described ice surfaces as “dirty, grayish, or even brownish” due to “dust” containing 16 diatomaceous taxa of algae, thought to be deposited from the atmosphere (Nansen, 1906; Nordenskiöld, 1875). Nansen (1906) identified the same algae on sea ice and in cryoconite holes (Garrett and Verzella, 2008). Nansen (1906) reported “big basilums of a simple rod-like appearance, rapidly oscillating and often forming long chains” on ice surfaces. Such filamentous bacteria were later recognised as crucial for cryoconite aggregation (Langford et al., 2014) and providing particulate OC for cryoconite ecosystems (Hodson et al., 2008).
Steinbock (1936) provided the only major study of supraglacial biota undertaken during the first half of the twentieth century, reporting cyanobacteria, various algae, protozoa, ciliates and other microorganisms in cryoconite on Disko Island (Greenland). He expressed the opinion that cryoconite holes represent well-established and well-organised habitats for diverse organisms, and possibly facilitated the in situ survival of microbes throughout past ice ages, presaging Wharton et al. (1985) by almost 50 years. The longevity of cryoconite holes was questioned again much later by Hodson et al. (2007) and Anesio et al. (2009) amongst others who confirmed multi-year and possibly multi-decadal residence on glaciers. Apart from Odell (1949), who reported ice worms in Alaskan cryoconite, it was over two decades before further investigations into cryoconite biota were published, when Charlesworth (1957) observed algae, rotifers, pollen and even insects and small dead birds. Gerdel and Drouet (1960) found 13.2–20.1% of Greenland cryoconite to be composed of OM including cyanobacteria, unicellular desmids, rotifers, fungi and diatoms. Gerdel and Drouet (1960) noted that organisms in cryoconite holes primarily utilise radiation in the same waveband that is most effectively transmitted through ice and referred to measurements by Thacker and Babcock (1957) showing ∼10% of full sunlight is optimal for algal photosynthesis, implying that cryoconite holes might provide optimal light conditions for autochthonous C fixation. This concept was revisited by Cook et al. (2010) half a century later. Therefore, despite only recently becoming widely recognised, the concept of biotic-abiotic equilibration in cryoconite hole systems has been present in the literature for over half a century.
The early polar explorers did not overlook the impact of microbes, including those in cryoconite, as glacier melt catalysts. Nordenskiöld described them as ice’s “greatest enemy” and implicated cryoconite in the waxing and waning of earth’s ice over geologic time. He explained that dark algae on ice both “accelerates melting” and “drills holes all over the ice” (Leslie, 1879). This was frequently corroborated in the twentieth century, notably by Gajda (1958) who examined cryoconite scattered on ice surfaces in ubiquitous layers of varying thickness and concentrated ‘stripes’ ∼10 cm in width. In both, granules varied from fine to coarse and were reddish-brown in colour (probably due to the presence of filamentous algae and photo-protective pigmentation, although this was not explicitly identified). An important observation was the influence of sediment layer thickness on ablation rate, where uniformly distributed, thin layers of cryoconite had greatest melt impact. This was later revisited by Cook et al. (2010) and Telling et al. (2012a) who recognised the significance of sediment arrangement for NEP. Cryoconite, according to Gajda (1958), “undoubtedly has an effect on the melting of glaciers, ice and snow surfaces of land, rivers, lakes and seas”. Current efforts to understand the regional and global impacts of cryoconite phenomena are therefore grounded in over a century of scientific thought.
Therefore, by the end of the nineteenth century, ice surface microbiota was already known to be widespread, diverse, concentrated into cryoconite holes and responsible for accelerating glacier wastage. The arrangement of sediment within cryoconite holes was known to be a crucial determinant of albedo and radiation absorption was identified as a control upon biotic activity. Cryoconite holes were known to contain complex ecosystems to which minerals and inoculants were delivered largely by wind from various sources, supplemented by debris falls from valley sides and extrusion of material from the basal bedrock. There has long been an appreciation that cryoconite mass loading varies geographically and over time, yet we still lack understanding of spatiotemporal variations in spatial distribution of cryoconite. Crude linear upscaling of mass loading from the plot scale to the regional, ice sheet or even global scale by Anesio et al. (2009), Hodson et al. (2010a) and Cook et al. (2012) were foreshadowed by similar work in the late 1800s (Bayley, 1891), implying that our understanding of cryoconite as a global phenomenon has advanced little over the past century. Similarly, although a numerical basis for cryoconite hole formation has had attention our understanding of hole morphology has not progressed far beyond that of the early twentieth century. Cryoconite holes had been established as energy balance phenomena by the turn of the twentieth century, with researchers such as Nansen (1906) even recognising biological darkening of minerals contributing to melt differentials. What were major uncertainties a century ago remain so today, even at the broadest scale. This includes regional and global scale impacts of cryoconite phenomena, supraglacial biotic-abiotic feedbacks, relationship with climate change and the role of microbial processes in glacier scale melt dynamics. Linking microbial and glacier scale processes remains a major challenge for twenty-first century scientists. Crucially though, the twenty-first century has seen growing appreciation of cryoconite as a component of a wide and complex supraglacial ecosystem.
VIII Where should cryoconite science go from here?
1 Directing future research
Glaciology has progressed from an abiotic to a biotic paradigm; however much of the modern literature assumes the supraglacial biome to be simple, quasi-static and exclusively comprising organisms contained in discrete cryoconite holes and algal blooms. Regarding cryoconite, there remains a tendency to ignore the diversity of hole shapes and sizes present on natural ice surfaces and instead discuss (and sample) symmetrical, cylindrical holes with flat floors and even sediment arrangements. Since hole morphology impacts the photic conditions at the hole floor (Cook et al., 2010) and cryoconite microbes are sensitive to changes in their environments (Edwards et al., 2014b) it follows that hole morphology probably influences cryoconite microbial ecology and therefore deserves research attention. Furthermore, if cryoconite holes undergo morphological evolution and migration then their contribution to supraglacial melt might be continuous. It has also recently been recognised that cryoconite holes are not spatially isolated entities occasionally interlinked by ephemeral, discrete hydrological flowpaths, but rather represent dynamic sites of organic and inorganic matter storage and high microbial activity punctuating a spatially expansive aquatic ecosystem within the porous near-surface ice layer known as the weathering crust (Irvine-Fynn et al., 2012; Muller and Keeler, 1969). This weathering crust ecosystem is then widely connected to other glacial and extraglacial environments (Figure 7). Since the top of the weathering crust represents the interface between ice and atmosphere and is exposed to incoming solar radiation, it is the primary site of radiative and sensible heat exchange and its properties therefore strongly influence supraglacial melt rates. Key variables in the surface energy balance include surface roughness and albedo, both of which are influenced by microbial activity, yet biology has not been incorporated into any predictive models of glacier melt. To do so will require better knowledge of the interactions between microbes and the ice matrix they inhabit. To improve our understanding of ice-microbe interactions, microbial ecology and biogeochemistry in the supraglacial zone, cryoconite and other microbial habitats must be studied in concert with the evolution of ice surface roughness, topography and hydrology. We propose a framework for integrating these biological and glaciological processes and introduce the term ‘biocryomorphology’ in reference to ice-microbe interactions.
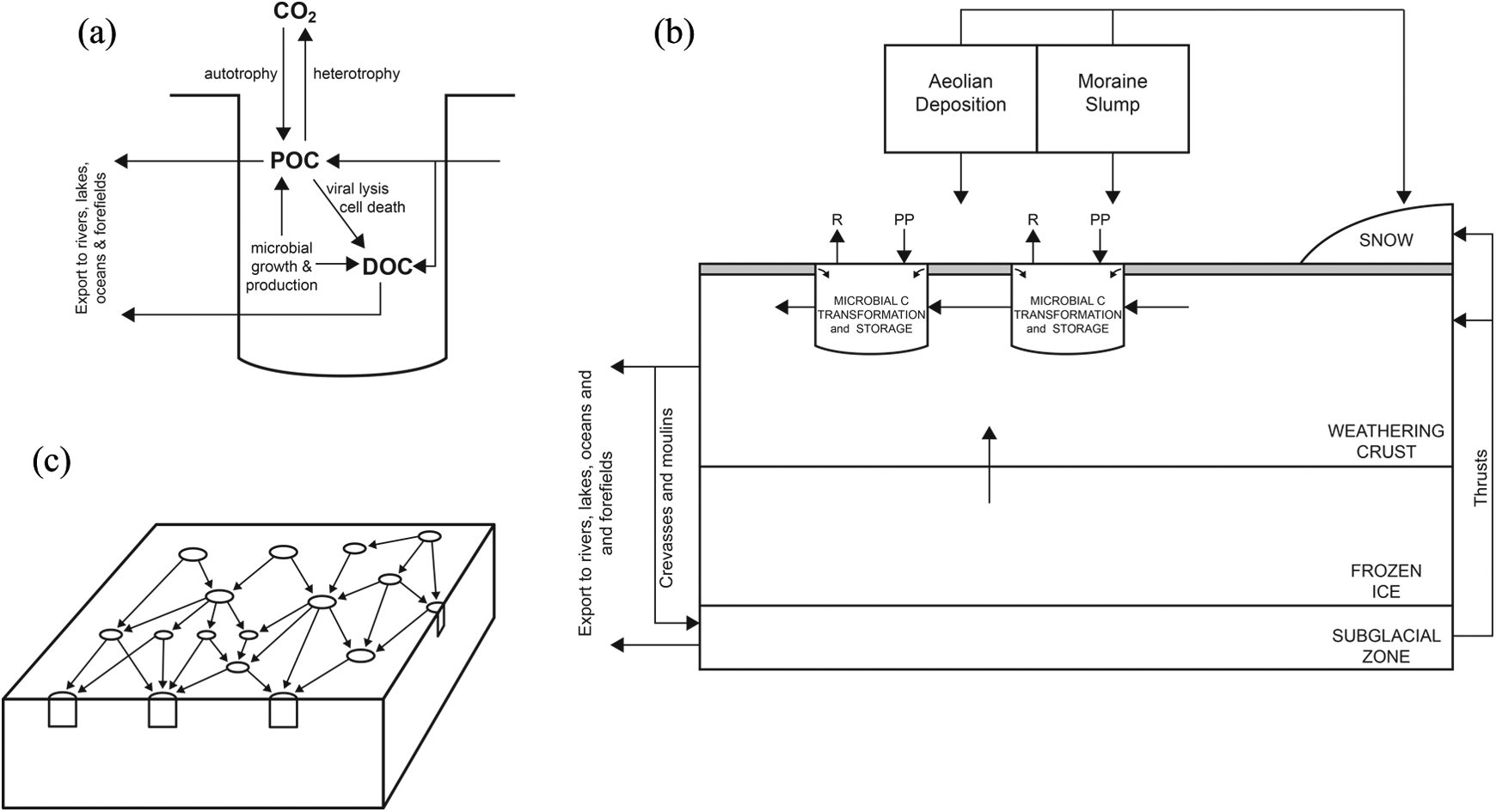
Three diagrams that show carbon fluxes through supraglacial systems. Black arrows denote flux of C. (a) Carbon cycling within an individual cryoconite hole; (b) Connectivity between cryoconite holes and other glacial and extraglacial environments; (c) Interconnectivity between cryoconite holes via hydrologic fluxes through the weathering crust. We gratefully acknowledge Antony Smith (DGES, Aberystwyth University) for his assistance with this figure.
1.1 Biocryomorphology
A systems-based strategy for studying cryoconite holes has been suggested before (MacDonnell and Fitzsimons, 2008; Stibal et al., 2012); however a robust framework for implementing such a strategy is lacking. To address this issue, we suggest integrating biological and glaciological processes, in particular ice surface roughness, near surface hydrology and microbial ecology under the term ‘biocryomorphology’. The overall aim is to develop a holistic, process-based understanding of ice-microbe interactions for integrating microbial processes into glaciological, biogeochemical and climate models. Cryoconite provides a crucial starting point for biocryomorphic investigations, since there are reciprocal relationships between microbial activity, hole morphology and surface energy balance that are yet to be constrained. Gribbon (1979) and Cook et al. (2010) imply that stable hole morphology is only likely in conditions of constant, even irradiance where there is no addition or growth of cryoconite granules and granules are evenly distributed over hole floors. This combination of conditions is unlikely, suggesting that holes often exist out of equilibrium and adapt their shapes and sizes in response to changing environmental conditions. The well-studied cylindrical holes probably represent a point along a morphological continuum that includes dispersed cryoconite, shallow and irregular holes, cryoconite alluvium, colluvium and stream-bound debris. This growth and movement of cryoconite holes indicates potential significance as catalysts of supraglacial melt and highlights their role in shaping ice surfaces. Furthermore, since microbial community structure-function relationships are known to be sensitive to environmental conditions, there may be concomitant changes in cryoconite microbial ecology. Irvine-Fynn et al. (2012) demonstrated transport of microbes through the top 1–2 m of porous ice on Arctic glaciers, indicating that models of cryoconite hole biogeochemistry should incorporate delivery and efflux of cells and nutrients via hydrologic flow through the weathering crust. Some measurements of nutrient delivery and export have been made in Antarctic cryoconite systems, notably by Tranter et al. (2004), Fountain et al. (2008), Bagshaw et al. (2013) and Telling et al. (2014). Interactions with other glacial (including seasonal and perennial snow, impermeable ice beneath the weathering crust and subglacial ice) and extraglacial (including atmospheric exchanges, moraines, rivers, lakes, oceans and glacier forefields) environments are certain to occur but are currently weakly addressed (Figure 7). Biocryomorphic investigations should take these connections into account. A holistic understanding of ice surfaces must incorporate cross-scale linkages from molecular-level processes to the dynamics of entire glaciers and ice sheets. The subsections below introduce specific research questions at a range of scales that provide crucial starting points for developing an integrated, biocryomorphic understanding of ice surfaces.
1.2 Molecular scale
Concomitant with the recognition of glacial surfaces as loci of biological activity, the molecular microbial ecologist’s toolbox has expanded rapidly. High throughput DNA sequencing of marker gene amplicons (for example 16S/18S ribosomal RNA genes), metagenomes, metatranscriptomes and microbial genomes promises a sequence-centred, systems orientated understanding of cryoconite ecosystems. However, the application of high throughput sequencing to cryoconite ecosystems is in its infancy. At the time of writing, only nine glaciers and the margin of the Greenland Ice Sheet (Edwards, 2014b; Hamilton et al., 2013; Segawa et al., 2014; Stibal et al., 2014; Zarsky et al., 2013;) have any peer-reviewed published cryoconite 16 S bacterial amplicon sequencing datasets and only the margin of the Greenland Ice Sheet has been studied over a melt season (Stibal et al., 2014); one metagenome is reported (Edwards et al., 2013b). To our knowledge, only one glacier has published eukaryotic marker gene amplicon data (Hamilton et al., 2013) and there are no published metatranscriptomes. Evidently sequence space in the cryoconite ecosystem has been poorly explored, but it is also clear that sequencing for its own sake could not be justified. Therefore, we propose that molecular tools be applied within a systems-orientated framework, which integrates across both the suite of ‘omics approaches (i.e. meta- genomics, transcriptomics, proteomics, metabolomics) and tools and approaches from biogeochemistry and glaciology. Such a framework could encompass mapping cryoconite ecosystems at spatial and temporal scales ranging from individual metabolic reactions to cell-mineral interactions to microbial dynamics within single granules, and reach beyond the scale of the parent ice mass. An integrated framework such as this would firstly serve to map cryoconite ecosystems across scales of space and time spanning several orders of magnitude, secondly understand the mechanisms of cryoconite ecosystem structure, stability and function, and thirdly, when integrated with climate and ice mass models, underpin a predictive understanding of cryoconite ecosystem dynamics in the context of contemporary climate change. At a time when projects with the objective of mapping Earth’s microbiome and its metabolic potential (Gilbert et al., 2014) are being realised, it is vital that ecosystems with the potential to sense and amplify the profound changes occurring in the cryosphere are not left out in the cold
1.3 Microhabitat scale
Cryoconite granules provide microhabitats for autotrophic and heterotrophic microbes. Having relatively few trophic interactions and abiotic influences, cryoconite granules might provide excellent model systems for microbial ecology in other environments. Their high density allows them to settle on ice surfaces their albedo is lowered by biological activity, making them the primary agents of biocryomorphic evolution. However, our understanding of granule formation and stabilisation remains basic and there are currently only a few studies (e.g. Langford et al., 2010; Takeuchi et al., 2010) explicitly examining granule initiation, growth and erosion. Key gaps in our understanding of cryoconite microhabitats include the origin of cryoconite materials, the mechanisms by which those materials are assimilated into granules, their longevity and stability, granule morphodynamics and granule-scale microbial ecology. Links between atmospheric processes, including wet and dry deposition of minerals, biota and anthropogenic compounds onto ice and snow, post-depositional transformation, redistribution and interaction with cryoconite granules are unknown. Biogeochemical transformation and storage within cryoconite granules, which may govern the export of organic and inorganic compounds to other environments is poorly understood. These processes determine the properties of granules which collectively determine hole morphology and therefore influence patterns of albedo and melt rate that shape the wider ice surface, thereby connecting molecular and cellular level processes with those operating at the habitat scale and beyond.
1.4 Habitat scale
Diverse hole shapes and sizes on ice surfaces imply evolution and even migration of cryoconite holes in response to changing surface energy balance and sediment dynamics. The mechanisms by which this occurs are currently unknown but likely depend upon microbially-mediated growth and stabilisation of cryoconite granules and heterogeneous melt rates across hole floors, which are in turn driven by granule distribution and patterns of light and shade. Ice surface topography may therefore control hole morphology by determining patterns of hole-floor shading, and is also reciprocally influenced by cryoconite-induced changes in surface energy balance. Lateral equilibration (Cook et al., 2010) should also be integrated into a biocryomorphic understanding of cryoconite evolution since variable granule distributions may drive asymmetric hole expansion, providing a mechanism of migration.
The storage of cryoconite, the photic conditions on hole floors and the continuous flow of meltwater through the weathering crust make cryoconite holes analogous to continuous flow immobilised biomass bioreactors, which are commonly utilised in industry to optimise synthesis of various biomolecules. Based upon cyanobacteria research in other environments, potentially valuable biomolecules synthesised in cryoconite holes likely include UV-screening pigments such as scytonemin and mycosporine-like amino acids (Cockell and Knowland, 1999; Garcia-Pichel and Castenholz, 1991; Quesada and Vincent, 1993), photosynthetic pigments such as phycoerythrin or phycocyanin (Hodgson et al., 2004; Squier et al., 2002) and complex carbohydrates that can be used as industrial thickeners, gellifiers and stabilisers in the paint, cosmetic, pharmaceutical and food industries as well as having potentially useful antiviral properties (Colica and Philippis, 2014). These same molecules might also represent useful biomarkers in the supraglacial environment (Langford et al., 2014). There is also great potential for microbial communities in cryoconite holes to provide novel antibiotic compounds, antifreeze proteins and cold-active enzymes (Singh et al., 2014a, 2014b). Future research could therefore fruitfully examine the potential of cryoconite as natural biofactories on ice surfaces, simultaneously improving our understanding of ice-microbe interactions and offering opportunities for novel research in biotechnology and bioprospecting. A holistic understanding of ice-microbe interactions must also take into account other microbial habitats, including surface-dwelling algae and cryoconite outside of cryoconite holes. Rates of C cycling in cryoconite dispersed on ice surfaces have been estimated several times (e.g. Chandler et al., 2015; Hodson et al., 2007) and the importance of algal blooms for supraglacial albedo and biogeochemistry have been investigated by Yallop et al. (2012) and Cook et al. (2012); however, spatiotemporal changes and interactions between these environments remain unknown. Analogising cryoconite holes as bioreactors, examining their morphological evolution and incorporating additional microbial habitats will yield important insights into habitat-scale biogeochemistry and microbiology and facilitate the integration of molecular, microhabitat, habitat and glacier scale processes.
1.5 Glacier scale
Recent work by Irvine-Fynn et al. (2012) suggests the entire weathering crust (porous ice in the top 1–2 m of the supraglacial zone) represents a dynamic ecosystem through which cells are continuously transferred by meltwater. Due to both light intensity and ice permeability attenuating with depth in the weathering crust, there may be a depth dependent down-glacier transfer of cells (Irvine-Fynn and Edwards, 2014). As well as a medium for hydrologic and biologic transport, the weathering crust provides the substrate for cryoconite hole formation, and the role of cryoconite as a component of the weathering crust is uncertain. Cook et al (2015) recently demonstrated the utility of cryoconite holes as natural piezometers penetrating the weathering crust. Entrainment of microbes by cryoconite might punctuate the down-glacier transfer of cells and prolong their exposure to favourable light and nutrient conditions. Transformation of OC and modification of community structures in cryoconite holes might define to some extent the composition of OM exported to other environments and also influence glacier ablation. Cell fluxes, storage and growth within supraglacial ice were shown to be potentially important for glacier albedo feedbacks and seeding of downstream ecosystems by Irvine-Fynn et al. (2012), while weathering crust evolution might release and activate dormant microbes interred in near-surface ice (Dancer et al. 1997). Further research is required to advance understanding of microbial processes throughout the weathering crust, and this may help us to relate processes at the microbial scale to those operating within cryoconite holes, and then at the regional scale and beyond. This will be essential for developing predictive models microbially-mediated glacier melt.
1.6 Landscape scale
Figure 7 illustrates connections between the supraglacial zone and other glacial and extraglacial environments. This includes inputs from moraine slumps, snow melt and atmospheric deposition, transport of cells and nutrients through the weathering crust, storage and biogeochemical transformations in cryoconite holes and ultimately export of nutrients, biomolecules and biomass to rivers, lakes, oceans and forefields. These wide interconnections urgently require research attention because landscape-scale processes respond to, and establish the initial conditions for smaller scale biocryomorphology. An integrated biocryomorphic understanding of supraglacial ice-microbe interactions will therefore necessarily include landscape-scale processes.
IX Conclusions
This paper reviewed cryoconite literature from the earliest reports to the present day. Early polar explorers were shown to be remarkably prescient in establishing fundamental knowledge that continues to underpin cryoconite science today. We now have ample evidence to support early claims that cryoconite holes are biologically distinct from other habitats and we have begun to recognise specific community structures and functions defined by specific environmental niches. Evidence is also accumulating to support nineteenth century suggestions that cryoconite and other supraglacial microbes darken ice surfaces and contribute to glacier wastage. However, we also recognise that several assumptions that originated with the nineteenth century polar scientists are now outdated. This includes the traditional view of cryoconite holes as discrete, biologically isolated entities. Instead, they should be considered hotspots of biodiversity and biogeochemistry within a dynamic and continuous glacier-wide ecosystem with connections to extraglacial environments. The analogy of cryoconite holes as natural continuous flow immobilised biomass bioreactors may prove useful in understanding the productivity of these ecosystems despite multi-dimensional stresses. The community should also trend away from the assumption that cryoconite holes have simple and stable morphologies, since cryoconite can clearly be identified in a wide range of environments on glacier surfaces, and there are likely feedbacks between hole shape and microbial ecology. Adopting a ‘systems’ approach that focusses upon relationships and feedbacks in and between the various glacial and extraglacial zones will allow us to better understand cryoconite as a bioglaciological entity. Attention should be placed upon linking processes at the scale of individual microbes, cryoconite granules, cryoconite holes, plots, catchments, glaciers and landscapes. This should ultimately integrate bio-glaciological, hydrological, meteorological, climatological and geological processes. To this end, we identified several specific areas of interest for future studies, particularly building upon recent work framing cryoconite holes as components of a dynamic weathering crust ecosystem and employing molecular analyses, and suggested a ‘biocryomorphic’ framework for achieving an integrated understanding of the role of cryoconite in the cryosphere.
Footnotes
Acknowledgements
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Royal Society (grant number RG2013) and NERC (grant number NE/K000942/1). Nozomu Takeuchi acknowledges Grant-in-Aid No. 262470.