
Abstract
Almost a century ago, observed geographic patterns of plant phenology (such as leaf-out and flowering) were summarized in Hopkins’ Bioclimatic Law. This law describes phenology as varying along climatic gradients by latitude, longitude, and altitude. Yet phenological patterns are not only affected by contemporary climatic differences across space, but also by underlying geographic variations in plant genetics that arise from long-term climatic adaptation. The latter influence on geographic patterns in phenology has been undervalued to this day, mainly due to the difficulty of quantifying it. This study outlines a methodology for bridging this knowledge gap through delineating geographic adaption patterns using common garden and cloned plant phenology. Through synthesizing existing literature, typical geographic adaptation patterns in both spring and autumn phenology of many temperate tree species are identified. Under uniform environment, spring leaf-out of colder climate-adapted populations of a certain species is either earlier than warmer climate-adapted ones due to lower thermal requirements, or later because of higher chilling (for dormancy release) demands. The former leads to a countergradient pattern as it is opposite to an in situ observation, while the latter leads to a cogradient pattern. Autumn leaf senescence, on the other hand, expresses a consistent cogradient pattern that is related to latitude and constrained by the populations’ varied photoperiod requirements. These geographic adaptation patterns allow a clearer understanding of geographical variations in phenological responses to climate change, and provide a theoretical basis for spatially explicit phenological models. In addition, given that these adaptive patterns reveal genotype-based variabilities, they are potentially useful for more accurately tracking phenology-dependent ecosystem processes (e.g. species distribution) and non-weather-related vegetation changes. As a unique subfield of physical geography with broad environmental implications, this line of research needs to be further developed by furnishing a stronger and more explicit spatial structure into current phenological studies.
Keywords
I Introduction
Phenology investigates one of the most intriguing aspects of the environment, viz., seasonal rhythms of growth and reproduction cycles of living things on Earth (Lieth, 1974; Schwartz, 2003). Rooted in horticulture and agricultural meteorology, the scope of phenological research overlaps with climatology, biogeography, and ecology; and its significance is rejuvenated in connection to contemporary climate change (Morisette et al., 2009; Root et al., 2003). As a core of bioclimatology, which is “a science of relations between life, climate, seasons, and geographic distribution” (Hopkins, 1938), phenology features a focus on the timing of biological events that are driven by seasonal changes in meteorology. But the spatial dimension remains central in phenological inquiries, especially for sessile plants, as noted by Schnelle (1955) distinctively as phanogeographie (in German). A representative generalization of geographic patterns in plant phenology is embodied in the Bioclimatic Law (Hopkins, 1918, 1938), which explicitly links observed spatial patterns in phenological timing to primary temperature gradients by latitude, longitude (distance-to-ocean), and altitude. However, over the past century, the role of environmental/climatic drivers in shaping geographic patterns in plant phenology has lacked explicit consideration of the underlying genetic influence on the geography of plant phenology.
In particular, climatic controls of plant phenology operate at two drastically different temporal scales. One is instantaneous and accommodated by the phenotypic plasticity of plants, such as interannual weather effect on budburst timing. The other is long term, shaping plants’ genotypes through adaptation. The observed geography of plant phenology is hereby the result of both short-term and long-term climatic effects taking place over space at disparate temporal scales. Perhaps due to a limited integration of climatology and ecology (Pau et al., 2011), the two aspects of climatic influence have rarely been considered in tandem in identifying the causes of spatial variations in phenology. In fact, due to a ubiquitous emphasis on recent climate change, much attention has been given to short-term climate effects, while the genetic aspect of phenological variation is often neglected. As opposed to the directly measurable short-term climatic influences, long-term climate-shaped and genotype-based geographic variations are more difficult to quantify and separate from the observed patterns. Yet, for better understanding and modeling how the phenology of different species and populations of a species varies across space, it is critical to consider separately the phenotype-based geographic response and genotype-based geographic adaptation (Table 1). Here the geographic adaptation refers specifically to long-term climate-shaped genotypic differences over space, as being different from the geographic response that varies with year-to-year weather fluctuations.
Comparison of geographic response and geographic adaptation that control observed geographic patterns in plant phenology.
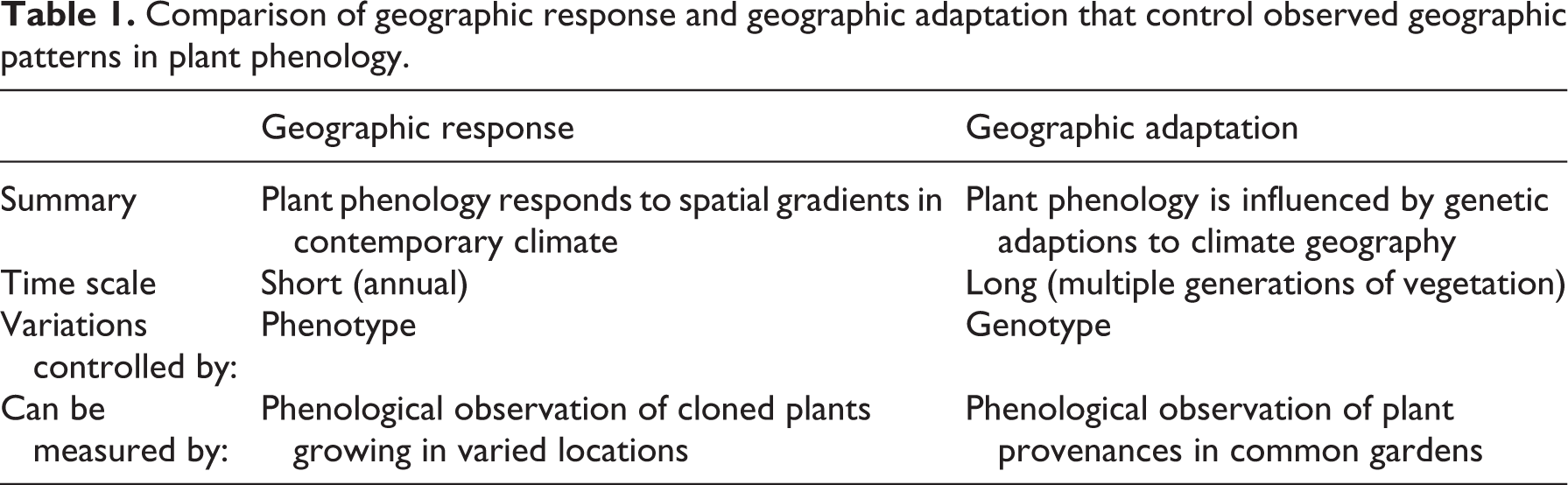
Clearly, genotype-based phenological variations are not completely determined by long-term environmental conditions via adaptation, but are also constrained by phylogeny (Davies et al., 2013; Davis et al., 2010; Pau et al., 2011; Willis et al., 2008). While more closely related species tend to leaf-out and flower at similar times, as phylogenetic conservatism indicates, they may do so because they are adapted to similar environments, implied by the notions of geographical conservatism and environmental filtering (Davies et al., 2013; Webb et al., 2002). Therefore, geographic adaptation may play a fundamental role in shaping genotype-based phenological variations over space, not only at the population and species levels, but also (at least in part) at the community and ecosystem levels.
Whereas the geographic response of plant phenology to climate gradients is relatively well-understood, the patterns of geographic adaptation remain unclear. Methodologically, geographic adaptation in plant phenology may be quantified using findings and principles of genecology, which studies adaptive differentiation of plant functional traits, often employing common gardens (Campbell and Sugano, 1979; Mátyás, 1996). But this body of research with particular respect to phenology is scattered, and has not been carefully examined in a way that would result in widely applicable geographic principles. On the other hand, the Bioclimatic Law offers foundational insight into the geography of plant phenology. But it relies on observed patterns only and falls short of accounting for the genetic diversity of plants, as noted previously. To move beyond the Bioclimatic Law, therefore, patterns controlled by geographic adaptation that are typical to many species need to be explicitly identified and clearly defined.
In this study, I synthesize literature from the rather loosely connected fields of phenology and genecology using common gardens, and incorporating findings from more recent cloned plant phenology research. The specific goal is to identify and define common geographic adaptation patterns and principles of plant phenology. This synthesis is focused on population level variations within respective species, but implications of the delineated patterns at the community and ecosystem levels are also discussed. Further, as the related knowledge base is mostly developed for temperature-driven plant phenology, especially for woody plants, I have focused the study on temperate trees. For plants growing in other environments, such as in drought-constrained climates where precipitation plays a more important role in triggering plant growth, additional research is required. Given the large amount of information on temperate trees and the typical seasonality they represent in the humid mid-latitude regions, the common patterns derived therein may serve as an initial yet fundamental step towards developing similar geographic principles in additional climates and biomes.
II Methods
Long-term climate-shaped genotypes interact with contemporary climate in complex ways to determine the timing of plant phenology. For many temperate plants, spring temperature drives phenology via two counter-balancing pathways (Chuine, 2000; Hunter and Lechowicz, 1992). First, a warming temperature is a prerequisite to resumed plant growth in spring, referred to as forcing or flushing temperatures. This is often measured with accumulated growing degree days or heat sum. Second, many temperate perennial plants only effectively respond to warming temperatures once winter dormancy (endodormancy) is released, which requires a substantial period of cold temperature accumulation (Coville, 1920; Rohde and Bhalerao, 2007; Schwartz and Hanes, 2010). This process is referred to as chilling or vernalization, through which plants are insured that the danger of frost has passed. The quantities of chilling and forcing, as well as the coordination mechanisms between the two, vary by species, and may be simulated in various ways (Chuine, 2000; Hanninen and Kramer, 2007). Further, photoperiod may interact with chilling and forcing to trigger spring budburst phenology. But its effect is limited to a few species and its role remains supplemental to the predominant effect of temperature (Basler and Körner, 2012; Caffarra et al., 2011b; Chuine et al., 2010). Autumn leaf phenology (growth secession and leaf senescence), on the other hand, though less studied than spring phenology, appears to be more routinely affected by shortening day length and cooling fall temperatures, with additional influences from environmental stresses (Fracheboud et al., 2009; Gallinat et al., 2015; Jeong and Medvigy, 2014; Xie et al., 2015). Therefore, on the one hand, plant phenology responds to year-to-year changes of these environmental factors (except for photoperiod, which is mostly static across years at a specific location). On the other hand, the geographic gradients of these climatic drivers entail a strong underlying geographic structure of plant phenology–climate (phenoclimatic) relationships that are embodied in different genotypes through long-term climatic adaptation.
Separating genotype-based geographic adaptation from geographic response with respect to their influence on the geographic patterns of phenology is possible through two methodological approaches (Figure 1). Both are based on the rationale that when one of the two variables is held as a constant, the observed variations are solely reflective of the other. First, by planting geographically different populations of the same species in a uniform environment, expressed phenological differences will be a function of genotypic variability only. This leads to an approach using common gardens, which permits the direct identification of adaptive variations within species in relation to different geographic origins. Field methodologies for common gardens (aka provenance tests), including the procedures for range-wide seed collection, the number of plantations, and the number of trees and replicates needed at each plantation, have been developed in forest genetics research (Wright, 1976). Many common gardens of tree species were established decades ago for silviculture and reforestation purposes (Morgenstern, 1996), and were later recognized to be valuable for examining intraspecific variations of plant growth in response to climate change (Mátyás, 1994, 1996). But most common garden studies were focused on juvenile plant survival and height growth, while phenology, as a supplemental trait related to growth, was observed only irregularly. Therefore, the phenological observation protocols used were often simple and lacking in detail. In spite of this limitation, essential adaptive variations of plant phenology are identifiable from common garden studies of many species.
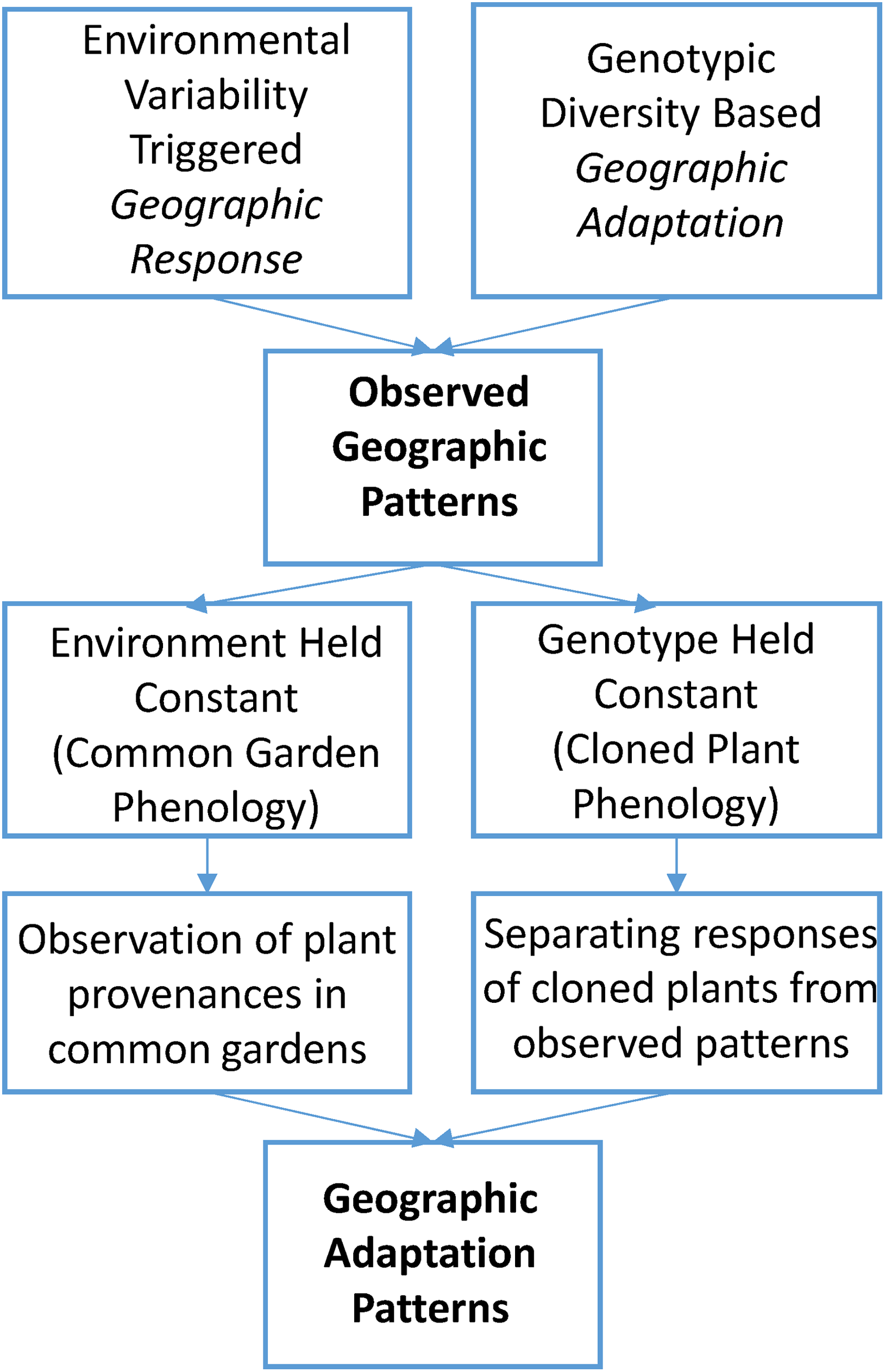
The methodological approaches to unveiling the geographic adaptation patterns in plant phenology.
An alternative approach holds the genetics constant while allowing the environment to vary, mainly through planting cloned plants (therefore, genetically identical) at different locations with diverse climatic conditions. The phenological response of cloned plants is uniformly a function of environmental differences. When compared to the phenology of common or naturally adapted plants, cloned plant phenology serves as a standard reference with respect to geographic response (cf. Table 1). Hence, to implement this approach two groups of individuals, viz., common plants and cloned plants, must be observed concurrently across a range of locations. The phenological difference between common plant and cloned plants reveals the degree of adaptation of the former. When these differences at various locations are plotted over an area of interest, the unveiled patterns would reflect variations contributed by geographic adaptation. The cloned plant approach is not as straightforward as common garden analysis and suffers a general lack of data. Although it complements common garden phenology by offering additional insights, cloned plant phenology research is still at its early stage of development (Liang and Schwartz, 2014). This synthesis is mostly based on ample literature on phenological observations in common gardens; limited findings from recent cloned plant phenology studies are discussed to provide a more complete view of the available methodological approaches.
III Adaptive patterns revealed in common gardens
The geographic adaptation variations observed in common gardens fall into three categories: (i) provenances from colder climates (e.g. related to higher latitudes or higher elevations) show earlier spring phenology than provenances from warmer climates (e.g. related to lower latitudes or lower elevations); (ii) conversely, provenances from colder climates have later spring phenology than those from warmer climates; and (iii) provenances from higher latitudes show earlier fall phenology than those from lower latitudes. The first case is referred to as being countergradient because the geographic gradient revealed in common gardens is opposite to that of populations growing in their source locations. And the latter two cases are cogradient given the same directions of the adaptation variations and the in situ observed geographic gradients. This terminology, adopted from evolutionary biology (Conover et al., 2009), helps illustrate and characterize the geographic adaptation patterns in phenology relative to the apparent geographic patterns (Figure 2). Below I provide examples of these patterns. While some representative cases are discussed in more detail, additional evidences are provided as citations.
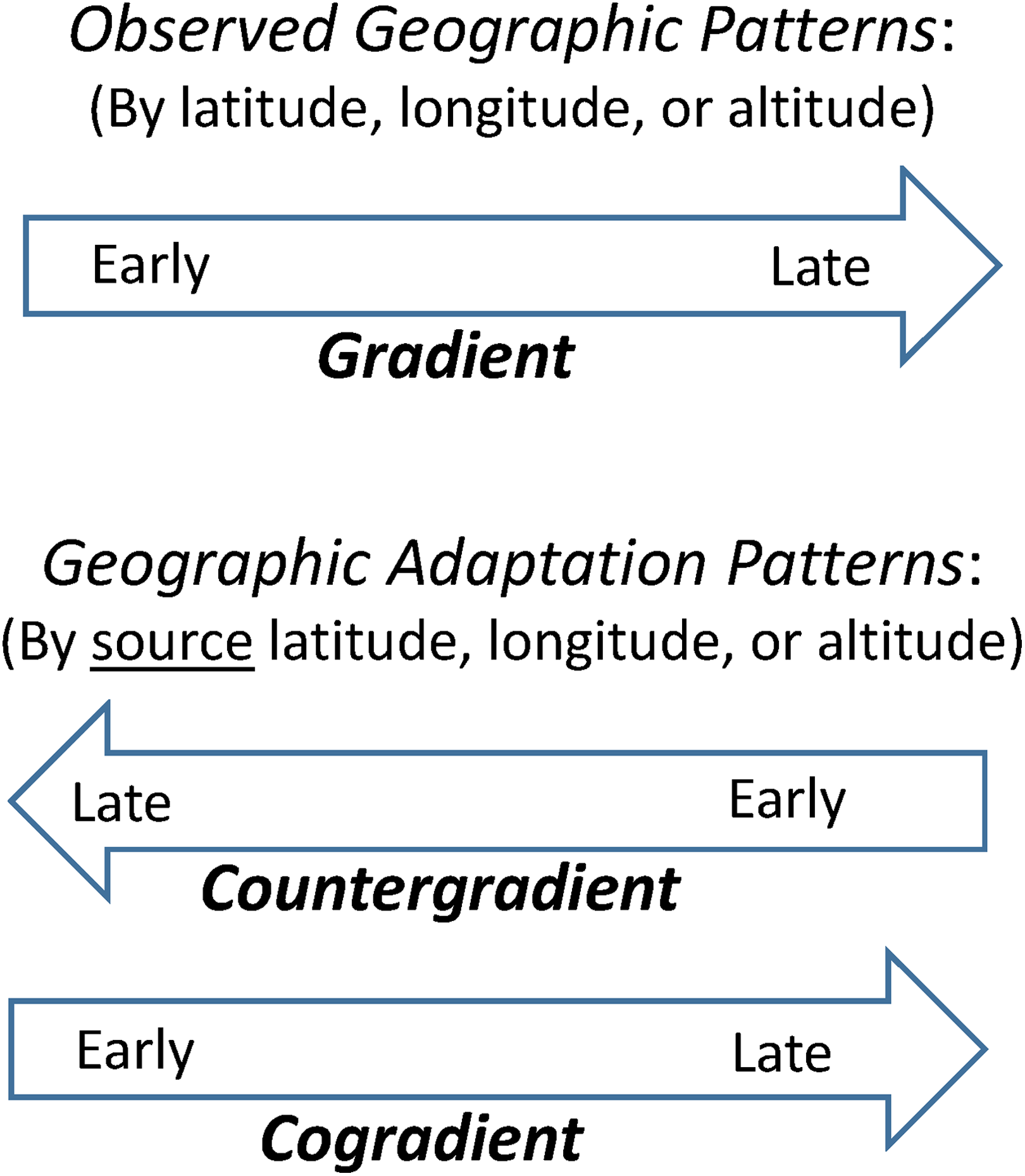
Concepts of countergradient and cogradient geographic adaptation patterns relative to gradient of observed patterns in plant phenology.
1 Countergradient pattern in spring phenology
A countergradient pattern in spring phenology involves bud break beginning with cold climate-adapted provenances and ending with warm climate-adapted provenances in a common garden. In a sugar maple (Acer saccharum Marsh.) common garden at Wooster, Ohio, northern provenances leafed-out sequentially earlier than southern provenances (Northern Hemisphere), showing a distinctive countergradient pattern (Kriebel, 1957). Further study at the same common garden using controlled chilling experiments showed that northern provenances required higher chilling than southern provenances for dormancy break, while under natural conditions at this common garden location in northern Ohio, trees from all sources obtained required chilling well before the end of the winter (Kriebel and Wang, 1962). Given that the chilling was not a constraint at this common garden, lower forcing requirements of the populations from colder climates led to earlier spring phenology, in turn manifesting a countergradient pattern.
This spatial trend is reversed (i.e. cogradient), however, for the same sugar maple progenies growing in a common garden in Gainesville, Florida, implying a delayed chilling fulfillment of the northern provenances at the warmer location (Kriebel and Wang, 1962). A similar experiment conducted at Gainesville, Florida, for red maple (Acer rubrum) also indicated that northern provenances of the species required higher chilling, while a provenance from the Everglades region of Florida showed no chilling requirement (Perry and Wu, 1960). Therefore, the expression of a countergradient pattern in spring phenology is related to a dominant influence of differential forcing requirements that are contingent upon an early fulfillment of chilling requirements for most provenances.
Additional common garden studies reporting a countergradient pattern similarly attributed this adaptation variation to differential forcing requirements of provenances. For instance, the spring phenology of balsam poplar (Populus balsamifera L.) was studied using data from two common gardens located, respectively, near the southern and northern edges of the species’ range in Canada and Alaska (Olson et al., 2013). The countergradient adaptive variation along latitude was more pronounced in the northern common garden than in the southern one, assumedly due to the fact that the chilling requirements of all provenances were fulfilled even earlier at the northern location, allowing for the effect of differential forcing to be manifested more thoroughly there.
Furthermore, the same countergradient pattern in Europe is sometimes more related to maritime-continental and topographic effects, leading to an adaptive geography predicted primarily by longitude and altitude. For example, leaf-out of European beech (Fagus sylvatica L.) occurred earlier in common gardens for provenances from inland Eastern Europe and higher elevations (Gömöry and Paule, 2011; Robson et al., 2013). Provenances adapted to warmer lowland and Atlantic drift-warmed coastal Western Europe appeared to require more forcing to trigger budburst, and thereby had delayed phenology in common gardens. In addition, the spring phenology of European beech is known to be sensitive to photoperiod (Heide, 1993; Vitasse and Basler, 2013), yet a lack of latitudinal gradient suggests that the influence of varied heat sum requirements is predominant in shaping its geographic adaptation pattern (Gömöry and Paule, 2011; Von Wuehlisch et al., 1995).
Examples of additional species that manifested a countergradient pattern in common gardens include silver birch (Betula pendula Roth) and downy birch (B. pubescens Ehrh.) (Myking and Heide, 1995); European ash (Fraxinus excelsior L.) (Pliura and Baliuckas, 2007); Norway spruce (Picea abies) (Eriksson et al., 1978); black spruce (P. mariana) (Johnsen et al., 1996); white spruce (P. glauca) and red spruce (P. rubens) (Blum, 1988); and Douglas fir (Pseudotsuga menziesii) (Campbell and Sugano, 1979).
2 Cogradient pattern in spring phenology
A cogradient pattern is observed as frequently as a countergradient pattern in the common garden studies of many tree species, which may have higher chilling requirements in general for dormancy release. In such cases, provenances adapted to colder climates leafed-out later due to delayed chilling fulfillment than those from warmer climates; and the effect of differential forcing requirements was apparently overridden. This is because if dormancy release is delayed from winter into spring, the start dates of forcing accumulation likely diverge for different provenances (with the cold climate-adapted ones being delayed further). In addition, the heat sum buildup occurs at a faster rate at a later date in spring due to higher effective temperatures, henceforth reducing the temporal differences of bud break caused by differential forcing requirements. These, in turn, lead to manifestation of the differences of dormancy break timing across provenances. In a study of red oak (Quercus rubra) in the southern Appalachians, provenances from different elevations were grown in common gardens located across an altitudinal range similar to that of the acorn sources (McGee, 1974). In all common gardens, bud break occurred sequentially from low-altitude provenances to high-altitude provenances, displaying a consistent cogradient trend. This phenomenon suggests a strong chilling-controlled spring phenology of the species.
In another case, leaf unfolding and seed germination of sessile oak (Quercus petraea) were also found to be earlier for southern provenances than northern provenances in common gardens (Alberto et al., 2011; Deans and Harvey, 1995; Ducousso et al., 1996). These common gardens of sessile oak were located in either low-lying French cities near the Atlantic coast (Alberto et al., 2011; Ducousso et al., 1996) or at Edinburgh, Scotland, which has a similarly temperate climate (Deans and Harvey, 1995). The locations of these common gardens optimized the chance of hindered chilling fulfillment for cold climate-adapted provenances. Although the role of chilling is not explicitly mentioned in the above studies, the adaption of sessile oak to higher risks of frost damage through delayed spring phenology in colder climates is acknowledged. A lack of understanding of chilling as a determinant of budburst timing variation, however, might have led to unjustified assumptions that low-latitude provenances require less heat sum for sessile oak (Deans and Harvey, 1995; Liepe, 1993) and other species such as white ash (Fraxinus americana L.) (Marchin, 2010).
Additional studies that documented a cogradient pattern of spring phenology include black cherry (Prunus serotina Ehrh.) (Barnett and Farmer, 1980; Cech and Carter, 1979); black walnut (Juglans nigra L.) (Bey et al., 1971); silver maple (Acer saccharinum L.) (Ashby et al., 1992); field elm (Ulmus minor) (Ghelardini et al., 2006); paper birch (Betula papyrifera Marsh.) (Hawkins and Dhar, 2012); and maritime pine (Pinus pinaster) (Desprezloustau and Dupuis, 1994). This partial list may imply generally higher chilling requirements of the species. But caution needs to be exercised to avoid overgeneralization, given additional influences of the climates and interannual weather fluctuations at respective common garden locations. Given the importance of understanding this context, below I elaborate in more detail the factors that control the expression of alternative geographic adaptation patterns in spring phenology.
3 Factors controlling the expression of adaptation patterns in spring phenology
The interactions of different environmental factors that trigger spring phenology are complex and difficult to entangle. The usually very large interannual variations of budburst phenology suggest that temperature’s role is predominant over photoperiod, given the stationarity of the latter. However, the temperature effect alone, comprising two counteracting mechanisms (i.e. forcing and chilling), is far from being straightforward, but may lead to varied and sometimes ambiguous expressions of geographic adaptation patterns. Detailed phenological observations with concurrent meteorological measurements in a white ash common garden (located in the species’ central range) suggested that while a strong chilling-controlled cogradient pattern may be shown in a year with a mild to normal prior winter, an unusually cold winter can lead to manifestation of a countergradient pattern instead in the same common garden (Liang, 2015).
Therefore, the expression of a countergradient or a cogradient pattern in spring phenology in a specific common garden likely depends on: (i) the respective levels of chilling and forcing that are required by a given species in general; (ii) the direction and magnitude of temperature change simulated by seed transfer from populations’ source locations to the location of the common garden; and (iii) the winter and spring temperature regimes during the years of observation at the common garden location. The first two factors have been discussed sufficiently. To further expand on the role of interannual weather change, the use of alternative scenarios can provide some insights. Specifically, in a cold winter-cool spring scenario, when chilling demand is unanimously met for all provenances in winter and forcing is built up gradually in spring, a marked, differential forcing-induced countergradient pattern is likely expressed. On the other hand, in a mild winter-warm spring scenario, when chilling fulfillment is significantly delayed and dormancy release sufficiently restrained for all provenances, and with fast subsequent thermal accumulation, a distinct differential chilling-controlled cogradient pattern would be manifested. Furthermore, in a mild winter-cold spring scenario, budburst of cold climate-adapted provenances is likely delayed due to insufficient winter chilling while the provenances from warm climate are similarly delayed hereafter by a lack of spring forcing, possibly leading to indefinite patterns. Finally, in a cold winter-warm spring scenario, earliest budburst is expected for all provenances because both chilling and forcing requirements are optimally satisfied. This scenario may lead to a weak countergradient pattern, given an early fulfillment of chilling requirements and a subsequent faster (compared to the cold winter-cool spring scenario) accumulation of forcing temperatures.
4 Cogradient pattern in autumn phenology
In comparison to spring phenology, autumn phenology of temperate plants is predicted by a less complex set of routine environmental cues, primarily declining day length and cooling temperature (Jeong and Medvigy, 2014; Yu et al., 2016). Perhaps contrary to the understanding that fall phenology is more complex than spring phenology, the environmental drivers of fall phenology are simpler in the sense that a counteracting temperature effect (i.e. chilling vs. forcing) is absent and the less variable day length plays a more significant role. Photoperiod is known to govern growth secession and dormancy induction of many trees (Rohde and Bhalerao, 2007). Although additional environmental factors such as heat, frost, wind, and especially drought may also trigger early leaf senescence in temperate forests (Leuzinger et al., 2005; Munn-Bosch and Alegre, 2004; Panchen et al., 2015; Xie et al., 2015), they operate more as stress factors rather than routine drivers under normal weather conditions. It is difficult, therefore, to tease out the impact of these factors on the geographical adaptation patterns of autumn phenology in temperate regions.
Notably, a nearly unanimous cogradient pattern of autumn phenology is found in common gardens in spite of disparate expressions of either a cogradient or a countergradient pattern in spring phenology of the same species (examples follow). As in situ fall leaf coloration or leaf fall occurs earlier in the north and later in the south (Northern Hemisphere), a cogradient pattern in fall phenology timing indicates an adaptation gradient that matches this observed geographic transition, which is the opposite to that of spring phenology. As evidenced by limited common garden studies using detailed phenological observation protocols, the cogradient pattern in fall phenology appears to be highly consistent across years and features smoother spatial transitions than that of spring phenology (Liang, 2015). Such a cogradient pattern can only be attributed to a latitudinal gradient of photoperiod rather than temperature. Otherwise, assuming that northern provenances are more tolerant to cooling fall temperatures, they would retain leaves longer than southern provenances. This would lead to a hypothesized countergradient pattern, which is contrary to the observation. Indeed, the high-latitude provenances are likely adapted to higher critical photoperiod (longer day length) thresholds (Rohde and Bhalerao, 2007) and, therefore, have earlier leaf senescence than the low-latitude provenances in common gardens. This is supported by the fact that photoperiod change plays a leading role in regulating the timing of fall phenology (Allona et al., 2008; Way, 2011). Such a mechanism favors northern populations by more effectively minimzing the risks of frost damage at higher latitudes. However, this does not rule out the role of temperature in the context of interannual variability. For plants growing at their original latitudes, while photoperiod sets up the fundamental limits to the timing of leaf senescence, temperature fluctuations (along with environmental stresses) do further influence the process of triggering plant growth secession, thereby causing interannual variations.
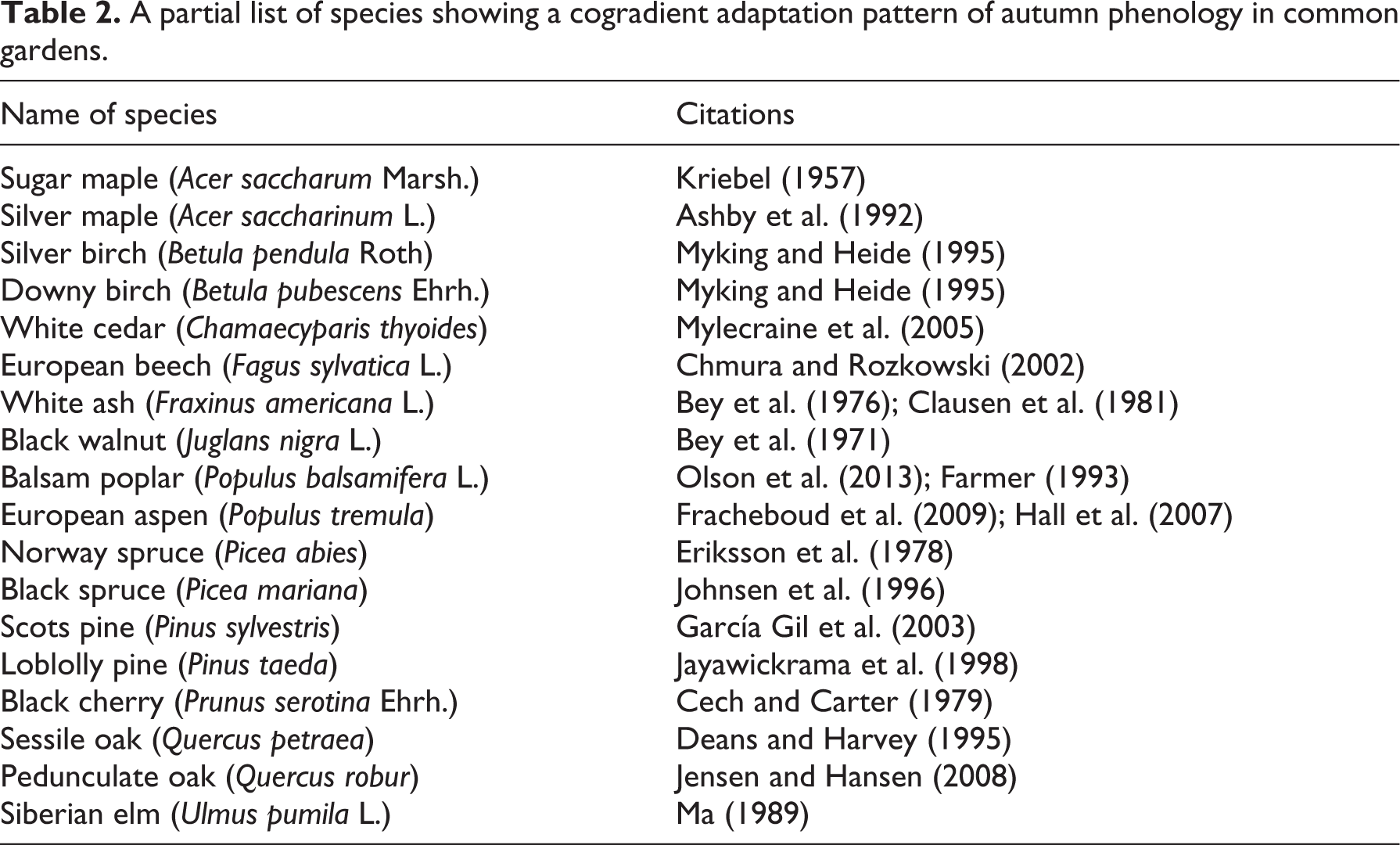
For all species surveyed, akin to an earlier review of North American trees (Nienstaedt, 1974), leaf fall, and leaf coloration almost always start from high-latitude populations, and end with low-latitude populations in common gardens. Such a cogradient pattern in autumn phenology is observed for many tree species provided in the partial list in Table 2. The large number of works on a cogradient pattern in fall phenology bears eloquent testimony to the existence of a strong photoperiod-sensitive adaptation mechanism. A diagram summarizing geographic adaptation patterns of both spring and fall phenology along a latitudinal gradient is provided in Figure 3.

Geographic adaptation patterns in temperate plant phenology along a latitudinal gradient in the Northern Hemisphere for both spring and fall seasons. The dash-line shape indicates a hypothesized but unobserved pattern.
A partial list of species showing a cogradient adaptation pattern of autumn phenology in common gardens.
IV Adaptation patterns revealed using cloned plant phenology
In addition to the common garden approach, geographic adaptation patterns in phenology can be derived by removing cloned plants-based geographic response variability from observed patterns (cf. Table 1, Figure 1). The spatial variation in the amount of difference between the concurrently observed common and cloned plants (e.g. departure of budburst dates) can be used to infer geographic patterns controlled by adaptive differentiation. Studies employing this approach are very rare, with only one published study to date (Liang and Schwartz, 2014). The dataset used in this study is for first leaf and first bloom of cloned lilac (Syringa x chinensis ‘Red Rothomagensis’), which has been observed within certain regional networks in the US since the 1950s (Schwartz et al., 2012). This effort has now been continued by the USA-National Phenology Network (USA-NPN) since 2009. The USA-NPN also collects phenological data of many other common plant species and included an additional cloned plant, cloned dogwood (Cornus florida ‘Appalachian Spring’). These data will undoubtedly increase the opportunities of using the cloned plant approach to investigate plant geographic adaptation patterns in the future.
Ideally, cloned plants should be compared with common plants of the same species, but such data are not currently available. A drawback of comparing cloned plants with common plants of a different species is that the absolute difference between the phenological timing of the two does not carry direct biophysical meaning. But given that cloned plant phenology is not influenced by genetic variations, the geographic gradients (not absolute dates) of phenology of different clones would reflect similar bioclimatic gradients. Therefore, the spatial gradient (not the absolute values) of the differences between the phenologies of cloned and common plants of different species can still reflect spatial adaption differentiation of the latter, hence indirectly revealing its geographic adaptation pattern. Indeed, the first leaf and first bloom dates of cloned lilac showed significantly steeper latitudinal gradients than that of common plants in the temperate eastern US (Liang and Schwartz, 2014). This finding implies that cold climate-originated natural populations are adapted to lower thermal forcing requirements, hence with relatively (vs. absolutely) earlier phenology with respect to cloned plants at the same locations, and vice versa for natural populations growing in warmer climates. This climatic adaptation leads to dampened latitudinal gradients for common plant phenology than that of cloned plant phenology, which reflects geographic response only. This relationship concurs with that of a countergradient pattern in common gardens due to differential forcing. The role of chilling is not obvious in cloned plant phenology studies given that all observations were taken in situ and chilling requirements were likely fulfilled very early in most occasions.
V Implications, applications, and future directions
1 Climate change impact on spring and fall phenology over space
The forcing-based countergradient and chilling-based cogradient patterns in spring phenology reveal two interwoven pathways of climate change impact on temperate trees. The countergradient pattern suggests that, in general, the same amount of spring temperature increase may trigger larger earlier shifts in spring phenology of cold climate-adapted plants than warm climate-adapted plants, contingent upon an early chilling fulfillment. A study using data from the USA-NPN predicted that the mean budburst date will likely advance more in the north than the south in the Northern Hemisphere under projected future climate scenarios (Jeong et al., 2013). On the other hand, contrary to a popular understanding of earlier phenology triggered by climatic warming, insufficient chilling may instead delay spring phenology of many species (Cook et al., 2012). However, current and anticipated magnitudes of climate warming (e.g. <2°C) are far less than climate change simulated by common garden experiments (e.g. 10–20°C, as in a space-for-time substitution scenario), making chilling shortage less likely to occur for locally grown plants on a regular basis. This generalization does not exclude cases when chilling fulfillments of certain species or vegetation in particular regions are hampered by anomalously warmer winter weather (Yu et al., 2010). Indeed, over the past century, climate change in the US, for example, has shown variability both seasonally and across regions, with temperature increase being more prevalent in the north and during the winter season (Capparelli et al., 2013; Menne et al., 2009). In addition, both observational and predictive studies have suggested that the spring phenology of northern forests is more likely affected by a chilling deficit during years with mild winters (Kaduk and Los, 2011; Liang and Zhang, 2016). Therefore, while the forcing-based countergradient mechanism is most applicable under the current climate regime, at least for specific regions and species that have relatively higher chilling requirements, it is necessary to consider the influence of an underlying chilling-based cogradient mechanism to avoid overestimating warming-induced spring phenological advances. In either case, the spring phenology of plants in warmer parts of the temperate climate may be less affected by global warming given their generally greater demands for heat energy as well as lower requirements for chilling.
Fall phenology, however, is affected differently across space by climate change given its unique environmental cues (Way, 2011; Xie et al., 2015). Although in addition to photoperiod and temperature, other factors, especially water stress, may trigger early leaf senescence (Leuzinger et al., 2005), precipitation is not a routine driver of fall phenology in temperate forests (Dragoni and Rahman, 2012; Liu et al., 2016). Hence, while environmental stresses may cause changes in the fall phenology of temperate trees at local to regional scales in specific years, the majority of the spatial variations in fall phenology is still governed by photoperiod and temperature. In fact, for the humid temperate forests in the eastern US and Japan, amid the general delaying effect of climate warming on the end of the growing season, northern tree populations and vegetation tend to be less sensitive (with smaller interannual variations) to temperature increase (reduced summer-to-fall chilling) than those in the south (Doi and Takahashi, 2008; Dragoni and Rahman, 2012). This latitudinal trend of climate change impact is opposite to that of spring phenology, and has important implications about the relative roles of photoperiod vs. temperature in regulating the timing of fall phenology. In the light of a consistent cogradient adaptation pattern in autumn phenology for most temperate tree species, this latitudinal difference in temperature responsiveness suggests that photoperiod gradient regulates the timing of growth cessation and leaf senescence predominantly. This is supported by the fact that in high-latitude regions with greater early fall frost risks, more rigid photoperiod control vs. temperature response appears to be a favorable adaptive trait. Whereas in the south, plants may be less constrained by photoperiod and, therefore, are more flexible in responding to temperature fluctuations, perhaps allowing some extra extension of the growing seasons. The larger and faster photoperiod change at the higher latitudes may also contribute to forming a stronger photoperiod cue in driving fall phenology in colder regions. There are, however, exceptions to this general relationship for certain species (Tanino et al., 2010). Nonetheless, given the leading role of photoperiod in regulating fall phenology according to the known physiology of many species (Allona et al., 2008; Rohde and Bhalerao, 2007; Way, 2011), the impact of climate warming on temperate trees in the autumn season is likely more constrained, and is probably more so towards higher latitudes.
2 Building spatially explicit phenological models
In the absence of clearly defined geographic adaptation patterns, predicting phenological variations induced by spatial heterogeneity in plants is hardly possible. Delineation of intrinsic adaptation geography in phenology, therefore, comprises an essential step towards building phenological models that are capable of predicting spatially explicit phenological responses to climate change. The existing phenological models, which are developed primarily for spring phenophases (leaf-out or flowering) of temperate woody species, have commonly focused on the instantaneous climatic effects on a few specific plant populations (Chuine et al., 2013; Hanninen and Kramer, 2007). To achieve a broad geographic coverage, one can utilize cloned plant phenology to produce standard responses to meteorological variables at the continental scale, as represented by the widely used spring indices models (Schwartz, 1997; Schwartz et al., 2006, 2013). Alternatively, more physiologically specific models (e.g. unified models) may be fitted for discrete natural populations of selected species across various locations independently (Chuine, 2000; Xu and Chen, 2013). The latter approach using multiple spatially independent models partly addresses the need to enable spatially explicit prediction capability, but is technically inefficient and lacks the power to extend the predictions to where data and models are not available. On the other hand, cloned plants-based models are advantageous in a sense that they offer continuous spatial coverage (limited only by the availability of weather data). Yet in both cases, an underlying spatial structure, as can be furnished by the geographic adaptation patterns defined in this study, is missing.
Moving forward from these previous developments, a way to construct spatially explicit phenological models, therefore, lies in combining the strengths of the above two approaches through incorporating the geographic adaptation patterns. The goal is to produce a single model over a region of interest for a given species and event (e.g. spring leaf-out) that is driven by both climatic and genotypic variables. Traditional models use fixed parameters fitted for specific populations. In a spatially explicit model, however, the parameters are geographically dependent. To better illustrate this idea, I offer the following conceptual model, using a simple 2-phase (chilling and forcing) sequential structure for spring leaf-out phenology as an example. A traditional approach is embodied in the following equations:

where Cc is the critical amount of chilling needed for breaking endodormancy (deep dormancy maintained internally by plants; Rohde and Bhalerao, 2007); t1 is the time when chilling starts to accumulate (may be arbitrarily set as an constant, e.g. October 1 of prior year), and t2 is when chilling requirement is met and endodormancy is broken; Rc is the rate of chilling, which is calculated from hourly or daily temperatures (T), according to specific base temperatures and response functions (Chuine et al., 2013).

where Cf is the critical amount of forcing needed after endodormancy break to complete ecodormancy (shallow dormancy maintained by environmental conditions) and trigger leaf budburst; t2 is the time of endodormancy break, and t3 is the date of budburst; Rf is the rate of forcing, which is similarly calculated from hourly or daily temperatures (T), according to specific base temperatures and response functions, in the simplest scenario, as growing degree days (Chuine et al., 2013).
In a spatially explicit model, the chilling and forcing requirements will vary across populations/locations, as can be predicted by specifically defined geographic adaptation patterns for a particular species, rather than independently for each population/location. Therefore, this additional level of variability needs to be incorporated into the previous equations as embedded functions, potentially in the following forms:
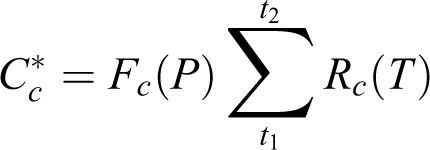
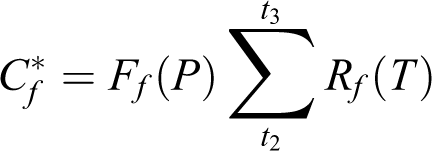
where
Given the diversity of phenoclimatic mechanisms of varied species, the spatially explicit phenological models may take on alternative forms with more or less complexity. For species that do not show obvious chilling requirements (likely satisfied early in winter), model structures may be simplified to include the forcing component only (Schwartz et al., 2013). Or, for species with more intricate environmental cues and physiological processes, more involved parameterization and model structure formulation are required (for a review of the different types of models, see Chuine et al., 2013). These cues and processes involve species with strong photoperiod responsiveness (Basler and Körner, 2012), differential dormancy induction dates (Caffarra et al., 2011b), parallel and alternating relationships between chilling and forcing (Hanninen and Kramer, 2007; Murray et al., 1989), and sensitivities to synoptic weather events (Schwartz and Marotz, 1988). Customized models need to be fitted to data for respective species according to the best known phenoclimatic relationships. Clearly, implementing these ideas and models requires substantial future work, not only because the detailed geographic adaptation patterns for particular species are yet to be accurately quantified, but also because our knowledge about the exact phenoclimatic mechanisms of plants remains scarce. At the current stage of developing spatially explicit phenological models, the generalized geographic adaptation patterns serve to substantiate the theoretical feasibility (i.e. proof-of-concept) and provide a geographic framework for more detailed works in the future.
In addition, modeling of fall phenology, though often neglected and perceived to be more complex than spring phenology (Gallinat et al., 2015), may turn out to be rather straightforward in a geographic context, given its highly consistent cogradient adaptation pattern. Improved fall phenology modeling is certainly needed for better constraining the growing season length. In addition, it may be necessary for accurately predicting spring phenology of certain species, given the necessity to account for different dormancy induction dates (Caffarra et al., 2011a). Apparently, winter does not always reset the calendar of perennial woody plants with respect to the cycles of growing and non-growing seasons. Unless there is a sufficient temporal interval between endodormancy break and the starting date of thermal accumulation, as in the case when chilling requirement is fulfilled early in winter before temperature rises above a critical threshold to allow for effective forcing, the timing of fall phenology in the previous year may well be a part of the equation for predicting the timing of spring phenology in the following year. A similar carry-over effect is reported for the spring-to-fall period (i.e. throughout the growing season) as well (Keenan and Richardson, 2015). Hence, fall phenology modeling should be given more attention and ideally implemented in tandem with spring phenology. Spatial structures akin to the conceptual model provided for spring phenology may be used, with photoperiod (incorporated according to the cogradient adaption pattern) and temperature as primary predictor variables. Overall, phenology as a precursory sign of more profound changes in the biosphere (details follow) should be more fully used in prognostic applications. Knowing that geographic diversities of phenological timing are not random and discrete over space, but are governed by traceable patterns predefined by long-term environmental gradients, allows us to enable spatially explicit capability for phenological forecasting, thus to bring this effort to a new level.
3 Tracking ecological processes and species distribution
The ecological consequences of phenological change in plant species are far reaching and occur in geographically explicit contexts. As a crucial component of ecological processes and functions, plant phenology regulates carbon exchange and primary productivity (Richardson et al., 2010), synchronizes or desynchronizes trophic interactions (Rafferty et al., 2015), and limits species distributions (Chuine, 2010; Chuine and Beaubien, 2001). In the face of climate change, the capacity for temporal shifts in organismal life cycles via phenotypic plasticity is not unlimited. Surpassing the limits of plastic response through timing changes of growth and reproductive events will ultimately affect the survival and development of organisms. Consequently, local extinction and/or microevolution of certain species are likely to follow, leading to species range shift and/or community reconfiguration (Chuine, 2010; Gienapp et al., 2008). The temporal adjustment of plants via phenotypic plasticity is thus intrinsically linked to the spatial adjustment of species in response to climate change (Chuine, 2010; Chuine and Beaubien, 2001). Such sequential change demonstrates that changes to a particular ecosystem in the temporal domain via phenological shift are precursory and early-warning signs of further and more drastic changes in spatial and genetic domains. Especially for sessile plants that cannot migrate as individuals and whose dispersal abilities are limited, the impacts of phenological change on community and ecosystem functions may be substantial. In addition, different rates of phenological change in related species that exist in predator–prey, herbivore–host, and mutualism relationships will lead to trophic mismatch and asynchrony in their life processes (Kudo and Ida, 2013; McKinney et al., 2012; Thackeray et al., 2010). Hence, drawing a clear picture of the intrinsic geographic adaptation patterns in plant phenology and building spatial models accordingly can potentially help determine the time and location of the subsequent transitions from phenotypic plasticity-supported timing shifts to more fundamental changes in genetics, distribution, and interaction of species.
In particular, the distribution of species is affected by phenology given that the survival, growth, and reproduction, hence, the fitness of plants, is closely tied to their ability to successfully fulfill the annual cycle of growth in a given climate (Chuine, 2010; Chuine and Beaubien, 2001). Previous studies hypothesized that at the northern limit, the fitness of populations of a temperate species is limited by frost damage and short growing seasons, while at the southern limit the restrictions are set by unfulfilled chilling requirements. Certain models have used phenology as a predictor of species distribution in line with these hypotheses (Morin et al., 2007, 2008). But these hypotheses and associated models have not taken into account the varied climatic requirements of plant populations within a species’ range, as reflected in the identified geographic adaptation patterns. For instance, southern populations of certain species may have lower chilling requirements with the result that the impact of warming on spring phenology is likely smaller compared to northern populations under the same magnitude of temperature change, as pointed out previously. Hence, there is a need to improve these phenology-dependent species distribution models by supplying explicit spatial parameters that represent the within-range genotypic variability of species. As a further step beyond developing spatially explicit phenological models, range-wide predictions of phenological change based on specific geographic adaptation patterns of a given species can be used as a critical input to improve the performance of species distribution models. In addition, integrating spatial models of phenology and species distribution may facilitate simulation and evaluation of the spatial variability of species interactions (Liang and Fei, 2014; Rafferty et al., 2013). In the long run, this will potentially bridge the temporal and spatial aspects of biospheric responses to climate change in a prognostic modeling framework. Such an integrated system with temporal shifts being precursory and range shifts being consequential components, for instance, may provide a more comprehensive and cohesive approach to the ecological impacts of climate change.
4 Monitoring inherent vegetation changes and biological invasions
In addition, given that the adaptive geography of phenology provides an intrinsic view of genotypic characteristics of plants that are free from instantaneous environmental influences (e.g. year-to-year weather fluctuations), it can be utilized to track inherent changes in vegetated landscapes. Remotely sensed land surface phenology has been employed for forest health monitoring using anomalous phenological departures as signals of disturbance (Hargrove et al., 2009), and mapping ecosystems based on diversified timing behaviors of seasonal land covers (Loveland et al., 1995). The current synthesis focuses on adaptation patterns at the population and species levels. But given that phylogenetic conservatism is expressed in the species’ geographical co-occurrence patterns (Davies et al., 2013), especially through environmental filtering (Webb et al., 2002), the geographic adaptation patterns in phenology are likely applicable at the community and ecosystem levels as well (Kaduk and Los, 2011). A recent study estimated the underlying climate adaptation patterns of temperate vegetation at the ecosystem level, using land surface phenology standardized by predictions from cloned plants-based models (Liang et al., 2016). These ecosystem-level geographic adaptation patterns in phenology, once periodically derived on a regular basis, may be used to more accurately detect non-weather-related vegetation changes, such as those caused by succession and disturbances.
The geographic adaptation patterns are applicable mostly to native and naturalized plants and communities. In a world with its biota being increasingly dispersed and rearranged over space, the phenological patterns determined at the ecosystem level are subject to modifications at specific locations. For example, many exotic and/or invasive plant species (mostly understory) often take advantage of vacant phenological/temporal niches in relation to canopy trees to increase their fitness, such as via prolonged growing seasons (Fridley, 2012; Polgar et al., 2014; Smith and Reynolds, 2015; Wolkovich and Cleland, 2011). Agriculture further adds to the uncertainties of applying the derived patterns to specific regions because crop composition and planting time are modified by human management, thereby missing a large part of the adaptive variation from natural vegetation. Hence, both exotic species and land cover/land use are expected to complicate the geographic adaptation patterns manifested, especially when looking at specific locations. On the other hand, however, departures from the predicted geographic adaption patterns can be used as a tool to gauge the degree and extent of human modification to natural landscapes. This idea echoes back to the rationale of using geographic adaptation patterns to detect inherent vegetation changes, as elaborated previously.
Furthermore, knowledge of adaptive geography in phenology can be potentially used to track biological invasions in novel ways. In particular, matching adaptive phenological traits of an introduced population with that of populations in the species’ native range can help reveal the geographic origin of the introduced population. This is a reversed application of the concept of common garden, in which the source locations of populations are known. Practically, this can be done by comparing the difference between phenologies of exotic and native populations of the species with that of a cloned plant, if they are observed in the field. Information from the exotic species alone (i.e. without a clonal reference) cannot achieve this because phenological timing varies across environments via phenotypic plasticity. The cloned plant phenology approach is, therefore, needed in this circumstance to identify the degrees of adaption for both exotic and native populations. But if resources allow, putting the introduced and native populations into a common garden or controlled experiment setting may achieve the same (probably better) result. Conceptually, this comparative phenology approach may also be used to discover invasive populations at various stages of naturalization/adaptation, hence to retrospect histories and pathways of specific introduction/invasion scenarios. Given that phenology is one of the most conspicuous functional traits of plants, knowing the linkages between its variations and the underlying genotypic differences is useful for quickly identifying intrinsic relationships among plant populations and species across geographical space.
5 Future directions
Continued and improved common garden phenology observation and analysis are still needed to determine detailed adaptation patterns for selected species. Many of the phenological observations recorded in literature were for juvenile plants during the initial years after the plantations were established. Young plants and adult trees, however, have shown different spring phenology due to ontogenetic changes, with the former generally leafing out earlier (Vitasse, 2013). This age effect on phenological timing matters for evaluating climate change impact on trees in growth chamber experiments using seedlings. But with respect to detecting the geographic adaptation patterns, the spatial relationships found for juvenile plants should still hold, because the provenances were observed at the same time and same ontogenetic stages. The consideration of ontogenetic differences of seedlings vs. trees, however, is needed for improving future phenological observations in common gardens. Especially, instead of establishing new common gardens, phenological observations in historically established ones (hence, with matured trees) are preferred to better match observations to responses of canopy trees in natural stands. Ideally, more dedicated phenological observation protocols with concurrent onsite environmental measurements should accompany this effort (Liang, 2015). An awareness of the value of historical common gardens for climate change impact and adaptation research has been increasing (Alfaro et al., 2014; Leites et al., 2012; Mátyás, 1996). However, many previously established (but later abandoned or not actively used) provenance test sites are yet to be discovered and properly maintained to allow access and data collection.
Further, to directly measure the phenoclimatic requirements of different plant populations, thus to more accurately specify the geographic adaptation patterns of certain species, controlled experiments may offer the best odds. To date, determination of chilling and forcing requirements of particular species have relied on indirect estimation using statistical methods (Ashcroft et al., 1977; Luedeling et al., 2013; Pitacco et al., 1992; Schwartz, 1997). Certain models are more constrained by known physiological processes, but still require statistical fitting (Chuine, 2000; Hanninen and Kramer, 2007). Certainly, the experimental approach used together with common gardens is labor- and resource- intensive, but it could yield significant information for precisely parameterizing spatially explicit phenological models. This effort is challenging and moves towards reaching the “holy grail” of linking phenological variations across physiological and climatological scales (Pau et al., 2011). Given the age effect noted above, cuttings from mature trees (vs. seedlings) are preferred in such experiments (Vitasse and Basler, 2014).
Along the lines of cloned plant phenology, further work is needed to include additional species and develop new methods to extract geographic adaptation patterns. This work is becoming more promising with the growing datasets from the Nature’s Notebook program of the USA-NPN, which covers both cloned indicator plants and naturally adapted species (Denny et al., 2014; Liang and Schwartz, 2014; Schwartz et al., 2012). In spite of the limitations in relation to measurement accuracy and precision that typically come with crowd-sourced data (remediable with effective quality control procedures), as well as mixed signals from horticultural plants, the USA-NPN dataset offers broad geographic coverage and a rich variety of species, and, therefore, provides unique opportunities for developing and testing spatially explicit phenological models.
Moreover, expanding the principles of geographic adaptation to areas beyond the temperate regions in relation to spatial gradients of additional environmental factors, such as precipitation, should not be ruled out in future studies. While more detailed work is still needed to further our understanding of the patterns in temperate plant phenology, and there is generally a lack of similar literature in support of a synthesis as such in other geographic domains, information from satellite remote sensing (along with available ground-based observation) may help initially explore ecosystem-level geographic adaptation patterns in additional biomes.
Finally, the dynamic interactions between life and climate, over both time and space, and across multiple spatial and ecological scales, converge at and are highlighted by the study of the geography of plant phenology. Continued exploration of the intrinsic geographic patterns in plant phenology, on fronts of both theoretical development and practical applications, will undoubtedly expand our scope of knowledge along this line of research. Beyond the Bioclimatic Law, which summarized the apparent geographic pattern in plant phenology, the current work proposes a further step to establish the underlying geographic principles, as embodied in the derived geographic adaptation patterns.
VI Conclusions
Phenology represents the biological aspect of seasonality, and its changes and geography vividly reflect the importance of unifying temporal processes and spatial patterns. This article synthesized knowledge concerning the intrinsic genotype-based variations of temperate plant phenology and identified corresponding geographic adaptation patterns. These patterns are characterized as being either countergradient or cogradient in common gardens relative to the in-situ observed geographic gradients in plant phenology. Both a cogradient pattern and a countergradient pattern are inherent to the spring phenology of many temperate tree species. The former is caused by different chilling requirements of populations, and the latter by different forcing requirements, either of which can be more dominant for particular species and/or under specific climatic conditions. Fall phenology of most tree species demonstrates a consistent cogradient pattern that is tied to a latitudinal photoperiod gradient.
The current work facilitates linking the temporal domain of plant activities more explicitly to its spatial domain, and provides meaningful guidance to important research topics in relation to bioclimatic interactions of organisms. Practically, the defined geographic adaptation patterns are useful for building spatial models that can track geographically distinct phenological responses to climate change, and to potentially improve the accuracy of predicting species’ range shift, among other associated ecological changes. Additionally, adaptive geography in phenology offers novel tools for monitoring inherent vegetation changes and biological invasions. More work is still needed to specifically quantify the geographic adaptation patterns for selected species, mainly through improved common garden observations and controlled experiments, and furtherance of cloned plant phenology research. Overall, a better understanding of the adaptive geography of plant phenology lays the foundation for tracking spatially variable interactions between climate and life. In the face of the environmental challenges in relation to climate–life relationships, phenology will play a more effective role with better-developed geographic theories and models. This synthesis serves as an initial step to make the way for accomplishing these tasks, and hopefully will contribute to furnishing a stronger and more clearly defined geographic structure in phenological studies.
Footnotes
Acknowledgements
I am grateful to my colleagues for their support in completing this work. Jonathan Phillips, Tony Stallins, Stan Brunn, and Tad Mutersbaugh reviewed the early draft of this manuscript and provided valuable comments and editorial help. Jonathan Phillips copy-edited the final draft of the manuscript. In addition, I appreciate input on literature review from Alison Donnelly. Finally, I thank the two anonymous reviewers for their insightful and constructive comments.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.