
Abstract
Sociology can no longer avoid engagement with biological ideas, but it can incorporate them where they are useful. Most biologically inspired explanations of sociological processes from outside the discipline are simple and, moreover, too reliant on biological rather than sociological models of social processes. Yet, it is possible to engage these efforts by developing sociological concepts and theories that meet those using evolutionary theory from biology. This paper argues that the heavy reliance on Darwinian natural selection limits sociological explanations, although this approach can help sociologists understand the evolved behavioral propensities of humans as evolved apes. These behavioral propensities cannot, however, explain the evolution and dynamics of the layers of sociocultural phenomena studied by sociologists, and efforts to do so with Darwinian notions of natural selection on individual organisms will always be inadequate. As an alternative, we propose that there are other types of natural selection inherent in the organization of what Herbert Spencer termed superorganisms. We label these Durkheimian, Spencerian, and Marxian selection, and they explain what Darwinian selection cannot: the dynamics and evolution of sociocultural phenomena.
Introduction
Perhaps the most significant trend in the social sciences has been efforts to bring biology back into the analysis of sociocultural systems. Among the social sciences, sociology has often been a reluctant participant in this intellectual movement, viewing efforts by sociobiology and evolutionary psychology as threats to the integrity of sociological analysis. Coevolution models have fared better (cf. Durham 1991; Richerson and Boyd 2005), but these, too, arouse suspicions among sociologists about the biological half of coevolution. Indeed, many generations of sociologists have held to the view that social structure and culture are emergent phenomena, requiring their own mode of analysis that is not reducible to either psychology or biology. What this emphasis tends to ignore is that a distinctly sociological analysis is not obviated by drawing from biology but, instead, such analyses are made even more robust. The best examples supporting this conclusion are urban ecology (Hawley 1986; Park 1936) and organizational ecology (Hannan and Freeman 1977) that have greatly enriched the analysis of two key types of corporate units—communities and organizations. These approaches view, respectively, selection as a driving force in (1) the differentiation of communities into distinctive areas of activity and (2) the rates of survival and death of organizations in resource niches inhabited by distinctive populations of organizations. The sociology of these approaches is sustained, but new insights are achieved by introducing selection as a key social process.
There is, however, some merit in the concern that approaches from outside sociology tend to ignore sociology. For instance, many approaches posit that Darwinian natural selection and the other three forces of biological evolution create cognitive, motivational, and behavioral propensities—for example, kin selection, altruism, reciprocity, reciprocal altruism, and so on—that become institutionalized in the structure and culture of institutional systems in societies (e.g., Alcock 2001, 2015; Williams 1966; Wilson 1975, 1978). But these efforts to explain this institutionalization in terms of evolved behavioral propensities cite very little if any of the large literatures in sociology on the dynamics of social systems. Furthermore, the agent-based simulations that sometimes accompany these “explanations” rarely specify the sociocultural mechanisms and processes by which such institutionalization occurs (e.g., Fowler 2005; Gintis 2000; Nowak 2006; Skirms 2000). With or without the simulation, most of these explanations become, in essence, “just so” stories about how behaviors installed by Darwinian natural selection cause the emergence of institutional systems. In Figure 1, we model in simple form this explanatory strategy. The last arrow to institutionalization is generally a “just so” story of how a deeply seated biological propensity (e.g., Dunbar and Barret 2007) or, in the case of evolutionary psychology (Barkow, Cosmides, and Tooby 1992; Boyer 2001; Buss 2004, 2011; Dunbar and Barret 2007), how a new brain module, causes the emergence of a meso- or macro-level sociocultural unit. While these stories are often interesting and provocative, they do not offer explanations that are interesting to most sociologists. They are highly discursive and, at best, speculative; they do not provide an explanation beyond telling a good story. Thus, while it is certainly true that Darwinian natural selection and other forces of evolution have instilled or enhanced in humans cognitive, motivational, and behavioral capacities, it is equally true that humans have used these capacities, purposively, to build up culture and social structure—a point echoed in Robert Boyd and Peter Richerson’s (1992) work on micro-evolution.
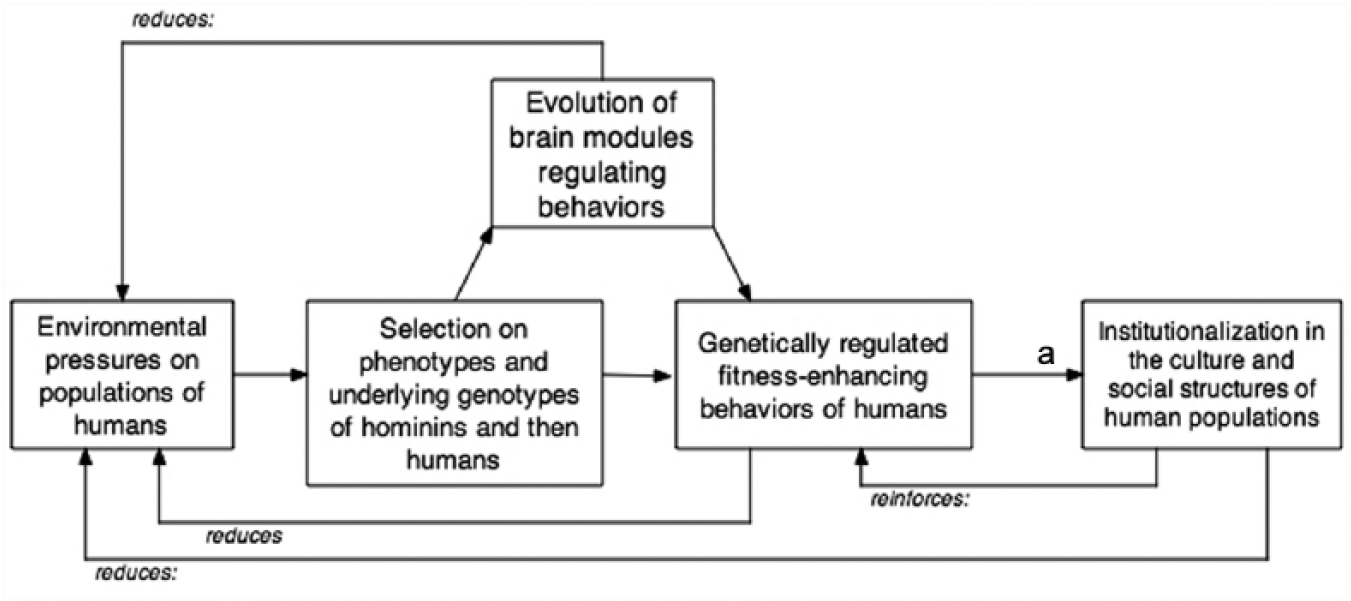
Limitations in the models of sociobiology and evolutionary psychology.
The issue thus becomes this: can we sustain an emphasis on selection as a key dynamic even when the variations on which selection works are no longer exclusively biological? There has been a tendency among those outside sociology to answer affirmatively, but many others inside sociology, even those highly sympathetic to a Darwinian explanation, recognize that something more is needed (e.g., Crippen 2015; Hopcroft 2015; Lopreato and Crippen 2001; Sanderson 2015). Moreover, many social scientists argue that the Darwinian image of natural selection working on variations in phenotypes or organisms (and underlying genotypes) needs some adjusting when applied to sociocultural phenomena (e.g., Boehm 2008, 2012; Boyd and Richerson 1992, 2005;; Richerson and Boyd 1999, 2001, 2005). To be sure, a number of early sociologists made much more than minor adjustments (e.g., Durkheim 1893; Spencer 1897), while an even larger number of contemporary sociologists (e.g., Abrutyn 2009, 2014b, 2015a, 2015b; Abrutyn and Lawrence 2010; Abrutyn and Van Ness 2015; Carneiro 2000, 2015; Chase-Dunn and Hall 1997; Runciman 1983-97, 2009; Turner 1995, 2010; Turner and Maryanski 2008b) and some cultural evolutionists (cf. Laland et al. 2011; West, El Mouden, and Gardner 2011) have made major adjustments and, as a result, have posited, often somewhat implicitly, the operation of new types of selection dynamics in social systems. In much the same vein, some nonsociologists who have entered the field have done the same thing by arguing for multilevel selection, while soft-pedaling the likelihood that multilevel selection may also mean multiple types of selection beyond Darwinian natural selection (e.g., Turchin et al. 2013; Turchin and Nefedov 2009).
In the end, it becomes evident that strict fidelity to Darwin’s view of natural selection cannot be sustained because Darwinian evolution is nonteleological, whereas the evolution of human sociocultural formations is often teleological and that goals are not wholly biologically wired in the genotype of individual humans. And although many have rejected consideration of other types of selection, Darwin himself considered teleological forces as important (see chapter 5 in later editions of his Descent of Man), and so much of the contemporary resistance to considering additional types of natural selection is far more dogmatic than Darwin ever was. Darwin and many sociologists recognize that sociocultural phenotypes are emergent phenomena, revealing their own distinctive dynamics as, in Herbert Spencer’s words, superorganisms. Some of these dynamics are parallel to natural selection among organisms, as long as we are willing to admit that the process of selection is very much affected by individual and collective actors who have capacities for articulating and pursuing goals, and moreover, for using their capacities for agency to create new variants on which another type of natural selection operates. In our view, then, natural selection is not only multilevel, but the very nature of natural selection varies as we move from the evolution of a species of organisms (such as humans) to the analysis of species of superorganisms composed of constituent parts that have capacities for formulating goals and for exercising agency to achieve these goals and, thereby, remaking not only the structure and culture of superorganisms but also the environments to which superorganisms and their constituent structures must adapt.
In Figure 2, we outline what Richard Dawkins (1976) termed survivor machines for genes but have drawn out the multilevel implications of his argument. The phenotype of the human body is a survivor machine carrying a precious cargo, genotypes, and by extension, human phenotypes are protected by the morphology of societies as they build ever-more complex layers of social structures: groups, organizations, communities, institutional domains, societies, and intersocietal systems. Natural selection operates at all levels of the embedded structures outlined in Figure 2, but once we leave the realm of human phenotypes, the selection working on the morphology of human superorganisms is no longer Darwinian. New types of selection begin to operate; and our goal in this paper will be to outline different types of selection dynamics, while maintaining some continuity with Darwin’s view of natural selection. To highlight the differences in types of natural selection, we have chosen to name them by invoking social scientists whose work overlaps with Darwin’s. We will thus distinguish among Darwinian, Spencerian, Durkheimian, and Marxian selection as a means of highlighting both the convergence and divergence of Spencerian, Durkheimian, and Marxian natural selection from Darwinian natural selection.
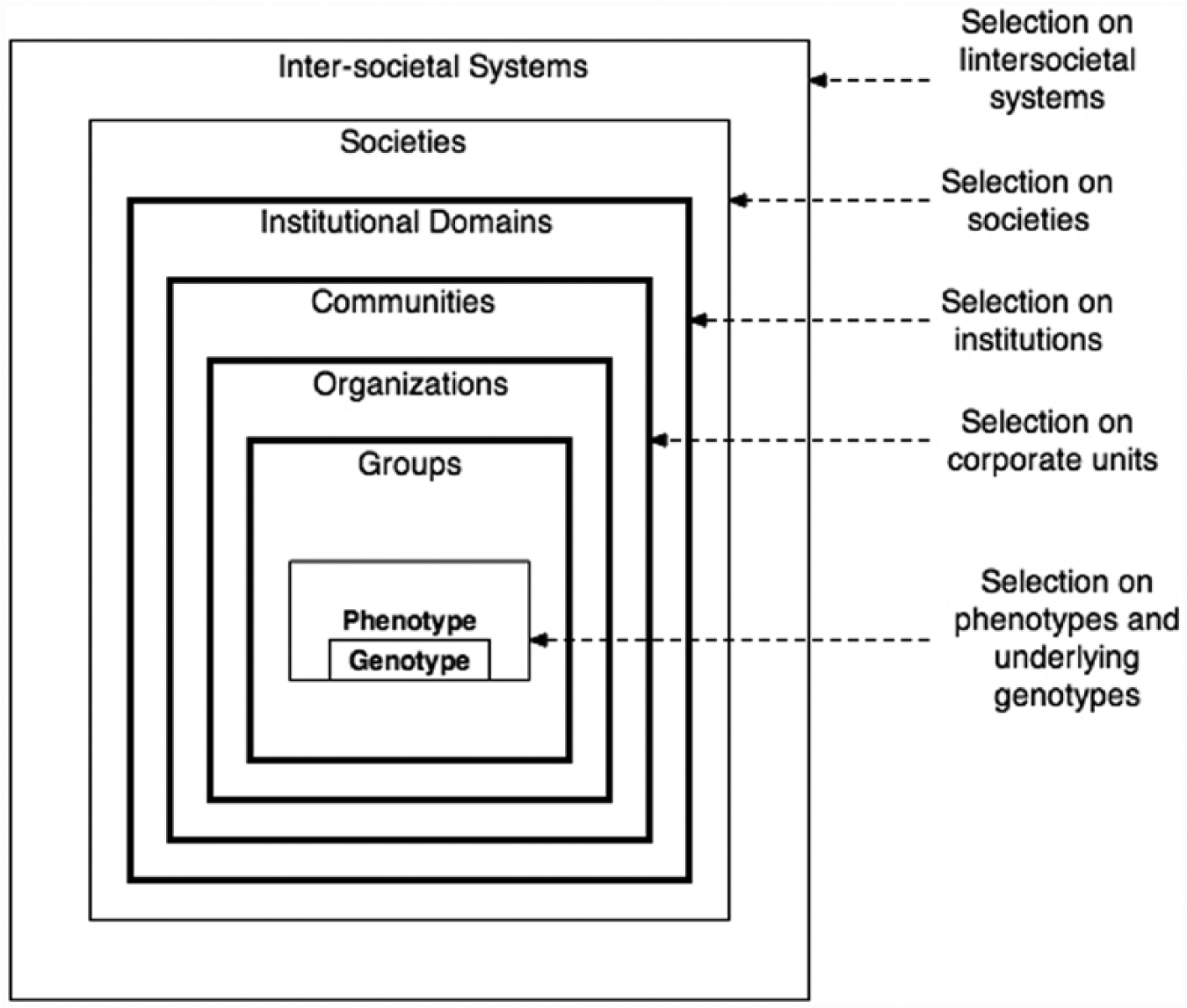
Sociocultural survival machines beyond biological phenotypes.
Types of Natural Selection
Darwinian Natural Selection
In the Modern Synthesis, the “forces of evolution” are defined as (1) natural selection, (2) mutations, (3) gene flow, and (4) genetic drift, with each affecting the distribution of genes in a population’s gene pool (cf. Mayr 2001). Darwinian natural selection is the process whereby the phenotypes of living organisms are selected upon by environments in which life forms seek to survive and reproduce. 1 If the environment changes, selection begins to work on those phenotypes that reveal features that enable organisms to survive and reproduce in the new environment, while selecting out those phenotypes less compatible with the environment. Thus, by nonteleological selection, the “fitness” of variations in phenotypes among members of a population is sorted out by selection, and from this process, the population of organisms evolves. Figure 3 lays out Darwinian selection as a distinctive process.
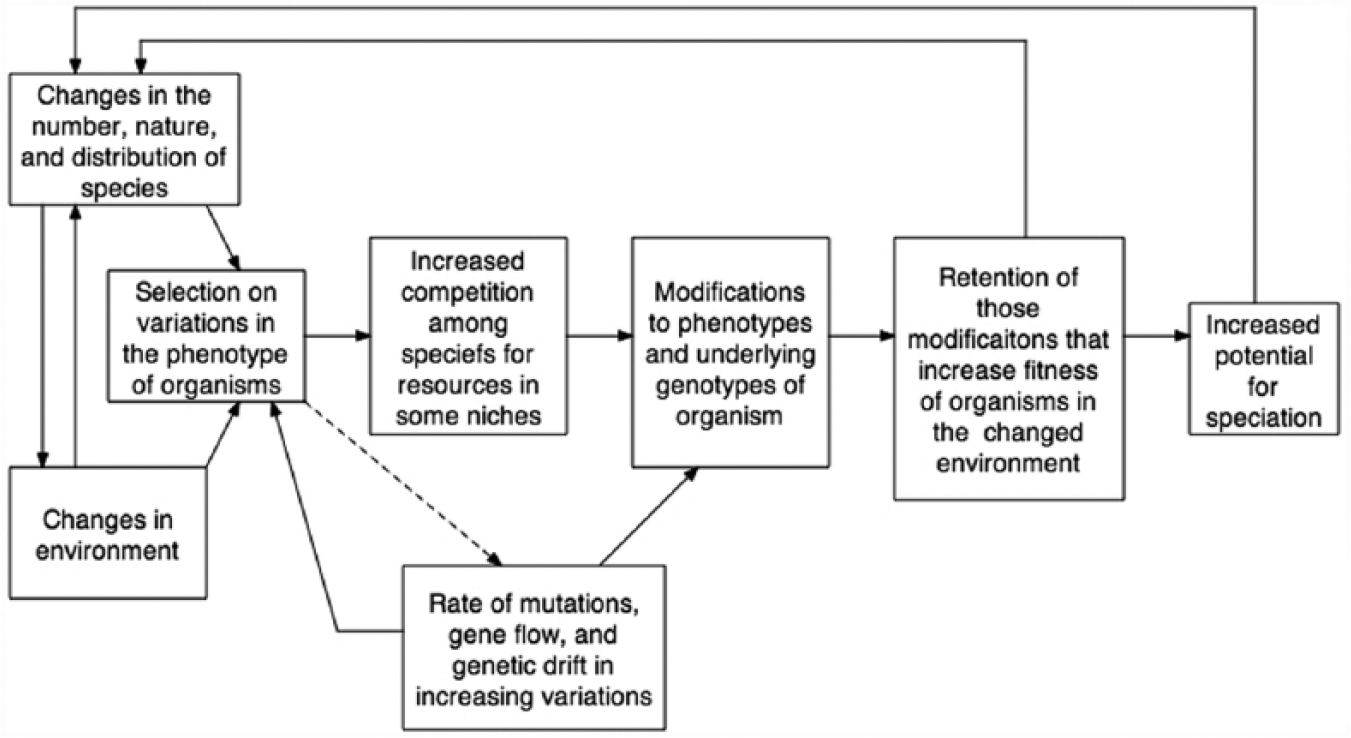
Darwinian selection as conceived by the Modern Synthesis.
The central force driving natural selection is environmental change that leads to more intense competition among conspecifics over resources and favors those phenotypic variations in members of a population that increase rates of survival and reproduction. While selection works on individual phenotypes, biology’s emphasis is on the evolution of the population as a distribution of phenotypes within a given resource niche in the environment. This last point is important because the unit of evolution in Darwinian theory, and the Modern Synthesis, is the population defined as a distribution of (1) phenotypes among members of the population and (2) genes in a “gene pool.”
For the last 150 years or so, social scientific research has shown that cultural transmission simply does not work like genetic transmission (Richerson and Boyd 2005; Runciman 2009; Steward [1955] 1972; Turner and Maryanski 2008a). In addition, the unit of evolution is not the population of phenotypes but, instead, various levels of sociocultural formation (see Figure 2). Hence, fitness—which is perhaps an ill-fitting word—is defined not by passing on genes or even something as fanciful as “memes” (Dawkins 1976), but by the capacity of a sociocultural formation to survive over time in an environment. Evolution for sociocultural systems involves changes in the nature, type, number, distribution, and integration of corporate units, defined as units revealing divisions of labor in pursuit of goals found in the cultural systems (beliefs, norms, technology, texts, etc.). As is evident in Figure 2, corporate units are of three basic types: groups, organizations, and communities. Thus, as one moves to higher levels of social organization outlined in Figure 2, Darwinian processes recede, and Durkheimian, Spencerian, and Marxian selection dynamics become increasingly evident as structural and cultural elements become the things upon which selection works.
Durkheimian Selection
Any particular corporate unit can be seen as potentially subject to the three types of non-Darwinian selection, but of particular importance is selection on organizations and communities because of their ability to coordinate and control the behaviors of relatively large groups of people. Corporate units tend to be embedded inside each other, as one mode of integrating them (Abrutyn and Turner 2011). Thus, groups are the building blocks of organizational systems creating divisions of specialized labor to realize a wide variety of goals, and, in turn, organizations are typically embedded in communities, which are also a type of corporate unit organizing geographical space into specialized zones, districts, and functions performed by corporate units (families, businesses, churches, governments, sports, recreation, etc.).
Both individually and collectively, as mutually embedded sociocultural systems, these corporate units are subject to selection forces on variants that enhanced their fitness or ability to persist as part of the basic structure of a society or intersocietal system. At the level of corporate units, selection reveals many Darwinian-like features revolving around the number and distribution of corporate units in various environmental niches where they seek resources; and with change in environments or increases in the number of corporate units in a niche, competition among such units increases, thereby selecting those with morphologies (or sociocultural phenotypes) that allow them to continue to secure existing resources, or find resources in new niches. So, Darwinian-like processes are in play, but the nature of the units of selection is very different, as is the process of selection on variants of corporate units. The persons inside these units, plus the units as a whole, all reveal the capacity for agency, which means that they can remake themselves (in a more Lamarckian than Darwinian way) and thereby create variations that allow them to sustain themselves over time. In addition, as Durkheim recognized, competition does not always lead to “death” of organizations, as agency can lead to specialization, diversification, and the construction of new resource niches that expand the carrying capacity of environments (McPherson 1981; McPherson and Ranger-Moore 1991). Indeed, goal-seeking corporate units in complex and dynamic societies can create not just new resource niches but new environments to which a population of corporate units must adapt.
Implicitly or explicitly, the twentieth century Chicago School employed Durkheimian logic in urban ecology: urban space represented an ecosystem in which humans, much like plant life or organisms, distribute themselves over local areas where they can secure resources (cf. Park 1936). This type of selection has found strong empirical support and greater theoretical precision (than the Chicago school could provide; see Irwin 2015) in organizational ecology, which studies the evolution of populations of organizations by Durkheimian selection (e.g., Aldrich and Ruef 2006; Chaves 2004; Hannan and Freeman 1977; Hannan, Polos, and Carroll 2007; McPherson and Ranger-Moore 1991) and by diverse sets of sociologists studying various types of sociocultural formations (Abrutyn 2015b; Eisenstadt 1964; Runciman 1983–1997, 2009; Turner and Maryanski 2008a). Durkheim, however, was the first to explicitly incorporate Darwin’s basic model to sociological analysis and, thus, we label sociocultural selection among units of human social organization Durkheimian selection.
As such, Durkheimian selection is characterized by four principles. First, the units on which selection works are organizations or, more generically, corporate units revealing internal divisions of labor (e.g., families, schools, businesses, governmental agencies, art galleries, newspapers, etc.). Second, consistent with Darwinian selection, emphasis is on the evolution of the population of organizations and corporate units and, therefore, their organizational forms (e.g., structure and culture) in a particular resource niche. Third, the dynamics driving selection are much like those specified by Darwin: population growth, increased density, escalating competition, selection, and death to less fit organizations, although Durkheim also argued that because of their capacity for agency, corporate units can often move to new niches, or even create new resource niches in which they can survive. Fourth, competition and Durkheimian selection always occurs within the boundaries of an institutional framework that, to some degree, shapes and limits how competition manifests itself (Fligstein and McAdams 2012). In Figure 4, we present a visual model of Durkheimian selection.
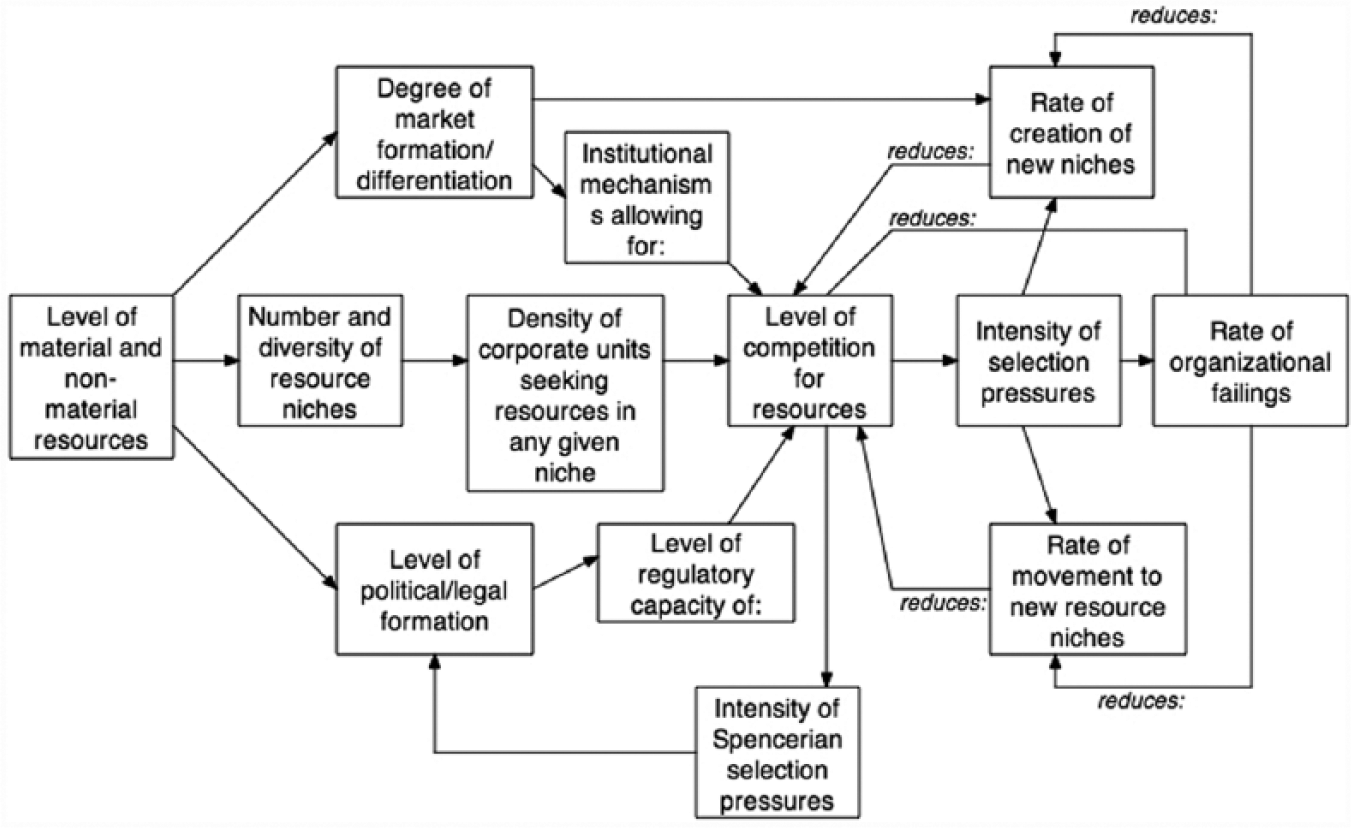
Durkheimian selection processes on organizational corporate units.
Spencerian Selection
Type 1 Spencerian selection
Long before Durkheim, Herbert Spencer formulated another view of selection that is essential to the sociological analysis of evolution of sociocultural systems. Although many of his ideas were converted to functionalism in the mid-twentieth century, if we go back to Spencer’s (1874–1896) original work, we can see Spencer’s formulation of an important type of selection that is essential in understanding human superorganisms. Spencer argued that, as societies evolve, they become more complex by elaborating different types of social structures (and corresponding cultures) along four fundamental axes: (1) production of resources needed for humans to survive and build social structures; (2) reproduction of individuals and the sociocultural formations organizing their activities; (3) regulation through the (a) consolidation of power, (b) codification of belief systems, and (c) formation of structural interdependencies through markets; and (4) distribution through the expansion of (a) infrastructures of moving resources, information, and people across territories, and (b) markets facilitating exchanges of resources, information, and even people (e.g., slave markets, labor markets). This emphasis on axes of differentiation is, however, a much more important contribution for conceptualizing sociocultural selection. In fact, Spencer was emphasizing selection pressures that are just the opposite of what neo-Darwinians or Durkheim emphasize because Spencer was arguing that under some conditions, it is the absence of existing variants of social structures or cultural codings capable of dealing with exogenous or endogenous exigencies that generate pressures for individuals and corporate actors to innovate organizationally, normatively, and symbolically. That is, under these conditions, selection does not sort among existing variants but pushes actors to create new variants that can deal with a real or perceived problem revolving around production, reproduction, regulation, and distribution. Spencerian selection, therefore, is Lamarckian in that it is driven by needs, motives, interests, and power relations to “acquire” somehow, perhaps anyhow, new sociocultural phenotypes (Abrutyn 2014a, 2015a, 2015b; Eisenstadt 1964; Runciman 2009; Turner 1995). Figure 5 outlines the essentials of Spencer’s argument for what we are terming Type 1 Spencerian selection. 2
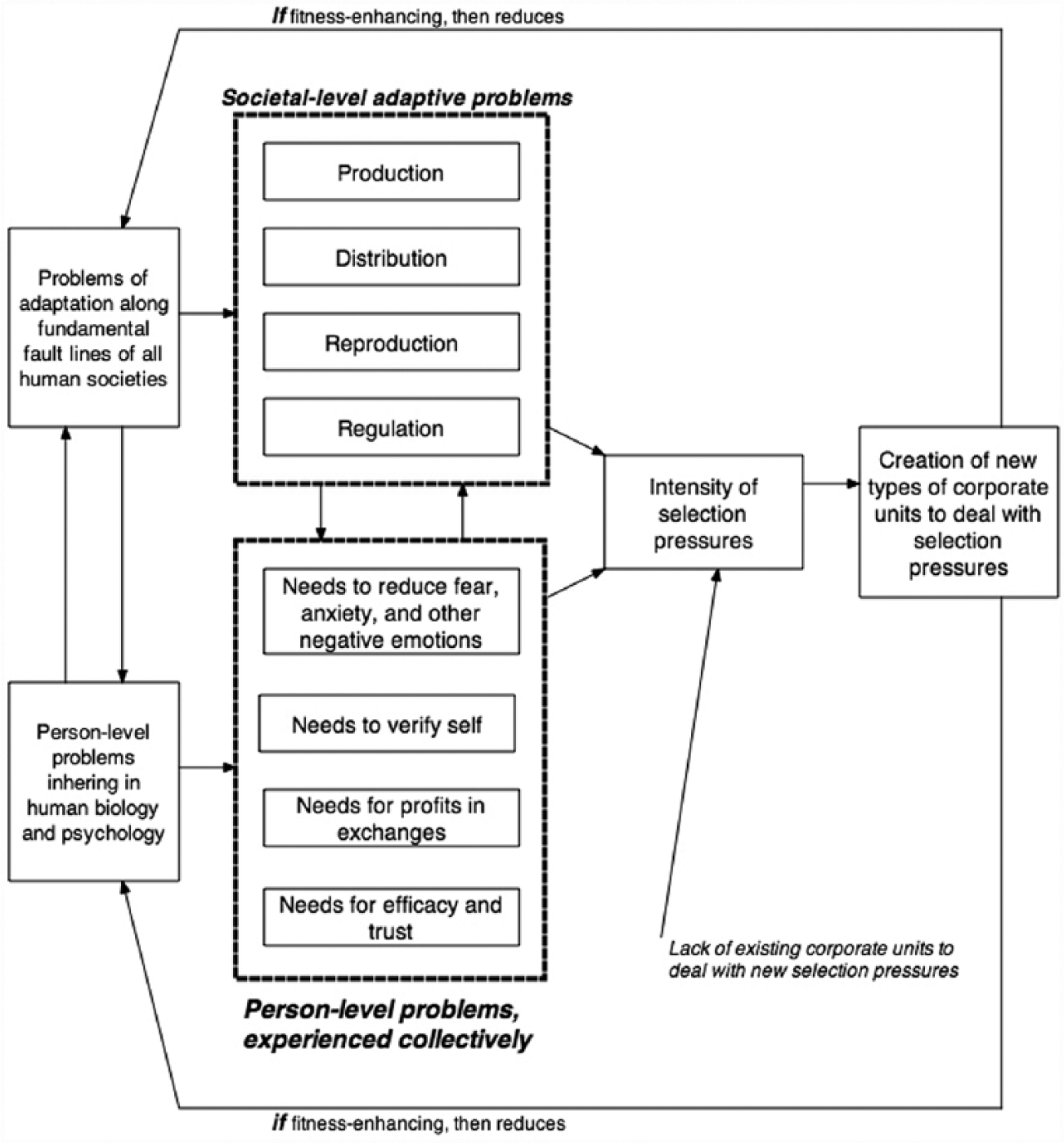
Type 1 Spencerian selection.
We can add another element to Spencer’s argument on what we are terming Type 1 Spencerian selection, one that he did not fully outline but that is important, particularly in the analysis of religion (see later discussion): the selection pressures created by basic human needs (Abrutyn 2009; Turner 2002, 2010). When a large proportion of a populations’ fundamental need-states are consistently not realized by existing institutional systems, these collective needs generate selection pressures on key actors in various institutional domains (e.g., polity, economy, religion) to create new forms of institutional systems. It is not unreasonable to assume, for example, that humans have fundamental needs to verifying self, for receiving “profits” in exchange relations with others and corporate units, for experiencing a sense of efficacy in at least some situations, for feeling a sense of trust in others, and perhaps for other such needs (Turner 2002). When large segments of a population cannot meet these needs, the negative emotional arousal—for example, fear, anxiety, shame, anger, frustration, sadness, and so on—can become collectively experienced, thereby arousing intense emotions that exert pressures on actors to find solutions to decrease this arousal—lest more Marxian selection pressures and intrasocietal conflict are generated (see below). Again, the pressure is not so much to select among existing variants of sociocultural formations, but to create some new variants of structures and cultures within institutional systems to reduce these selection pressures. Indeed, as will be outlined later, one of the most important sources of selection pressures for the evolution of religion has come from the failure of collectivities of individuals to meet basic human needs (see Turner 2014,2015 for analysis of these need-states).
Type 2 Spencerian selection
Spencer saw the evolution of societies from simple to more complex forms as very much driven by warfare, and as he turned to sociology in the 1870s, he increasingly employed his famous phrase (Spencer 1888 [1850–1851]), “the survival of the fittest,” to denote selection between societies when they go to war (Spencer 1874-96). Societies with larger populations from which to draw military forces, with greater consensus over values, ideologies, and beliefs, with higher levels of economic and military technology, with a polity that is centralized and yet legitimated (often by religious beliefs and elites), with an economy that produces large economic surpluses, with superiority in military organization, and with other advantages can typically win a war against societies revealing lower levels of all these capacities.
As one society conquers another, the diffusion of the dominant society’s culture and many of its organizational forms begin to generate another level of selection pressure on the indigenous cultures and social structures of a conquered society, particularly if domination rather than co-optation are employed to control members of a conquered population. Thus, the selection process inhering in war revolves around more-fit societies forcing some degree of consolidation of less fit societies into their institutional structures, thereby making, for the time being, an even larger superorganism. Much like genetic drift, but with purpose and intention, culture travels along with war and conquest, sometimes extinguishing local cultures, but more often combining in new structural and cultural variants. Over time, the institutional systems of the conquered change by a combination of Type 2 Spencerian selection and perhaps Durkheimian selection (see Figure 4). Thus, warfare and these associated dynamics have historically been one of the key forces driving “descent with modification” at the level of societies and intersocietal systems, and the internal cultures of even subordinated societies in the new geopolitical formation that evolves. The dynamics of this Type 2 Spencerian selection are outlined in Figure 6.
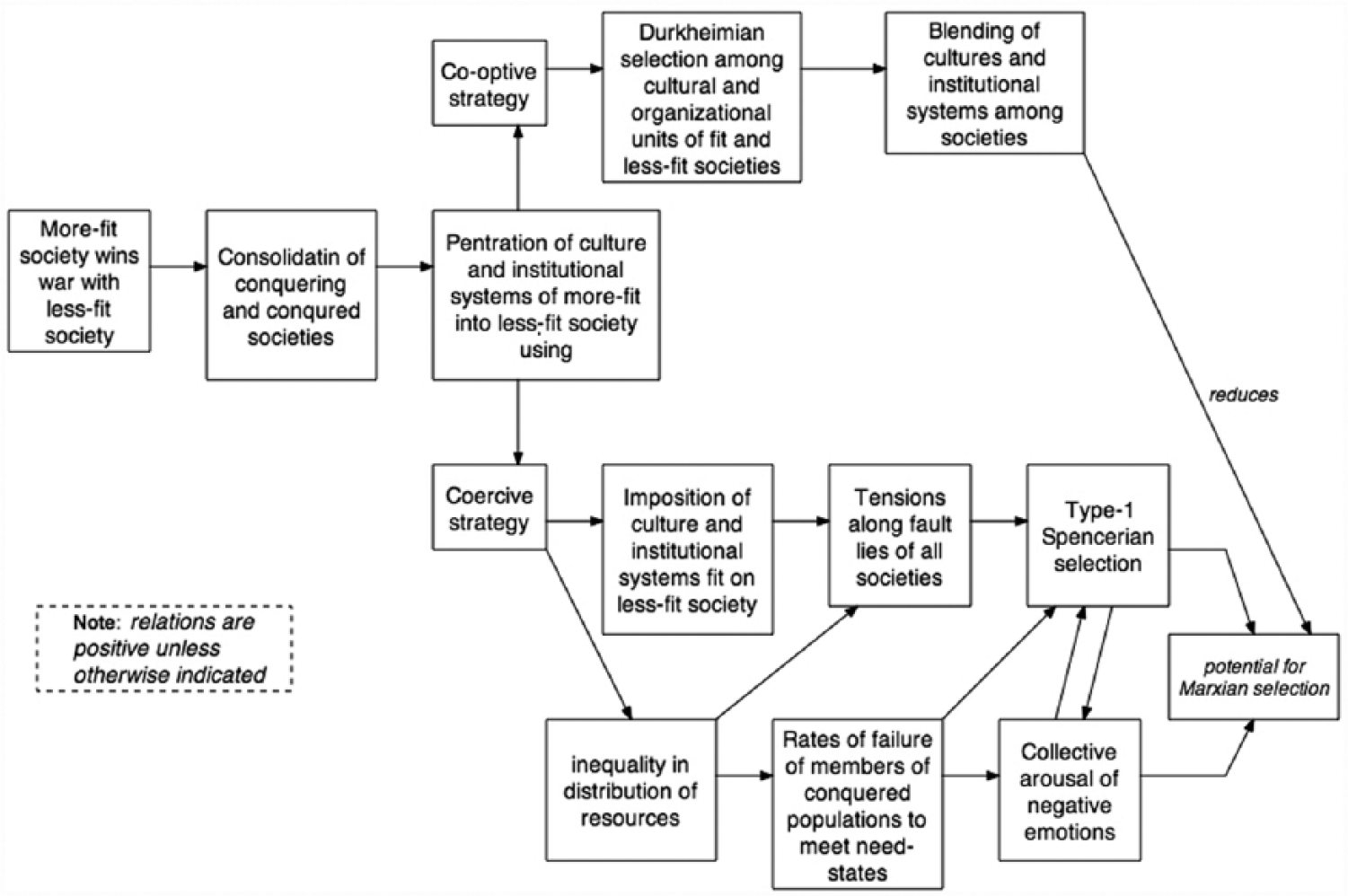
Type 2 Spencerian selection.
One force emphasized in Figure 6 is that conquest generally increases inequalities in the new geopolitical formation composed of the conquered and their conquerors. Inequalities always generate Type 1 Spencerian selection pressures, especially pressures for new forms of regulation but also distribution, production, and reproduction of the new institutional systems. Threat of revolts and other potential lines of resistance represent selection pressures on the institutional systems—polity, law, economy, and even religion—and often lead to adaptive responses that may work in the short run (e.g., coercive repression) but that themselves create a new level of new Type 1 selection pressures from those not receiving resources, not having opportunities, and not meeting fundamental human needs (installed by Darwinian selection). The stage is thus set for a round of what we view as Marxian selection in the midst of Type 1 and Type 2 Spencerian selection, often generated by the unfulfilled needs (that are part of the legacy of Darwinian selection) of the subordinated in societies revealing high levels of inequality.
The advantages to viewing selection as Spencerian, whether Type 1 or Type 2, is that evolution is neither inevitable nor unilinear. As the literature on entrepreneurship (Abrutyn 2014b; Eisenstadt 1964) and social movements (Snow and Soule 2010) demonstrates, agents rarely achieve their stated goals as articulated. Instead, adjustments and accommodations are typical (Abrutyn and Van Ness 2015), and incomplete success and ongoing efforts to maintain and expand success are the rule. In fact, societies are almost always in at least a mild state where Spencerian selection pressures from (1) fundamental fault lines of all patterns of social organization (i.e., production, reproduction, regulation, and distribution) and/or from (2) unfulfilled need-states create Type 1 pressures for new types of corporate units that can respond to these pressures. When these creative efforts prove fitness enhancing, they become the core of new institutional domains, as core actors continue to develop these successful corporate units; and as they do so, they also seek modes whereby they become more integrated. In fact, until mechanisms of integration are in place, the fitness-enhancing effects of new corporate units cannot be fully realized. All of the institutions of human societies have evolved this way, and continue to evolve in response to Type 1 and Type 2 Spencerian selection pressures.
Marxian Selection
Marx’s emphasis on the “contradictions” that always exist in the organization of production (save for his communist utopia) is another way of arguing for Spencerian selection. But what makes his analysis distinctive is that often these pressures cannot be resolved because institutional systems—for example, economy, polity, law, religion—reveal rigidities that keep them from innovating new solutions to problems of inequality (although the historical record, thus far, shows that these institutional systems were able to innovate more than Marx anticipated). As a result, those who are subordinate or marginalized and who do not perceive that they receive a fair share of resources will experience a variety of deprivations that accumulate into a set of grievances that, initially, are only vaguely articulated but that, nonetheless, arouse intense negative emotions (Goodwin and Jasper 2006; Jasper and Owens 2014; Turner 2010). The conflict process begins here, and under a number of conditions, leads subordinates to develop counterideologies against not only the legitimating ideologies of the political economy but also the institutional structure of a society (Summers-Effler and Kwak 2015; Turner 2010). As leaders articulate these counterideologies, they coalesce into an increasingly coherent ideology for revolution in which a class “in itself” becomes organized as a class “for itself,” willing to engage in conflict against the organizational structures and the ideologies imposed by elites that legitimate these structures (Snow 2004). This conflict can be violent at some point and lead to an overthrow of the existing political regime, with a new regime responding to Spencerian selection pressures for regulation and control, thereby imposing new systems of economic relations and political control that may lead to new types of Spencerian selection pressures that, if not managed, can lead to a new round of Marxian selection.
While some of Marx’s specific predictions may not be empirically valid, the key point is that certain conditions lead to conflicts that fall outside the bounds of Durkheimian selection because the conflict is over the very institutional systems—economy, markets, polity, law, and religion—that often regulate Durkheimian competition over resources. His ideas also go beyond Spencer who tended to emphasize warfare between societies, whereas conflict-based selection outlined by Marx occurs within and between societies. For much of sociocultural evolution has involved conflict as a selection process between mobilized corporate actors, with the parties to the conflict most able to secure and mobilize resources, the most likely to prevail and, thereby, to structure or restructure new variants of sociocultural formations. Figure 7 outlines the elements of this type of selection.
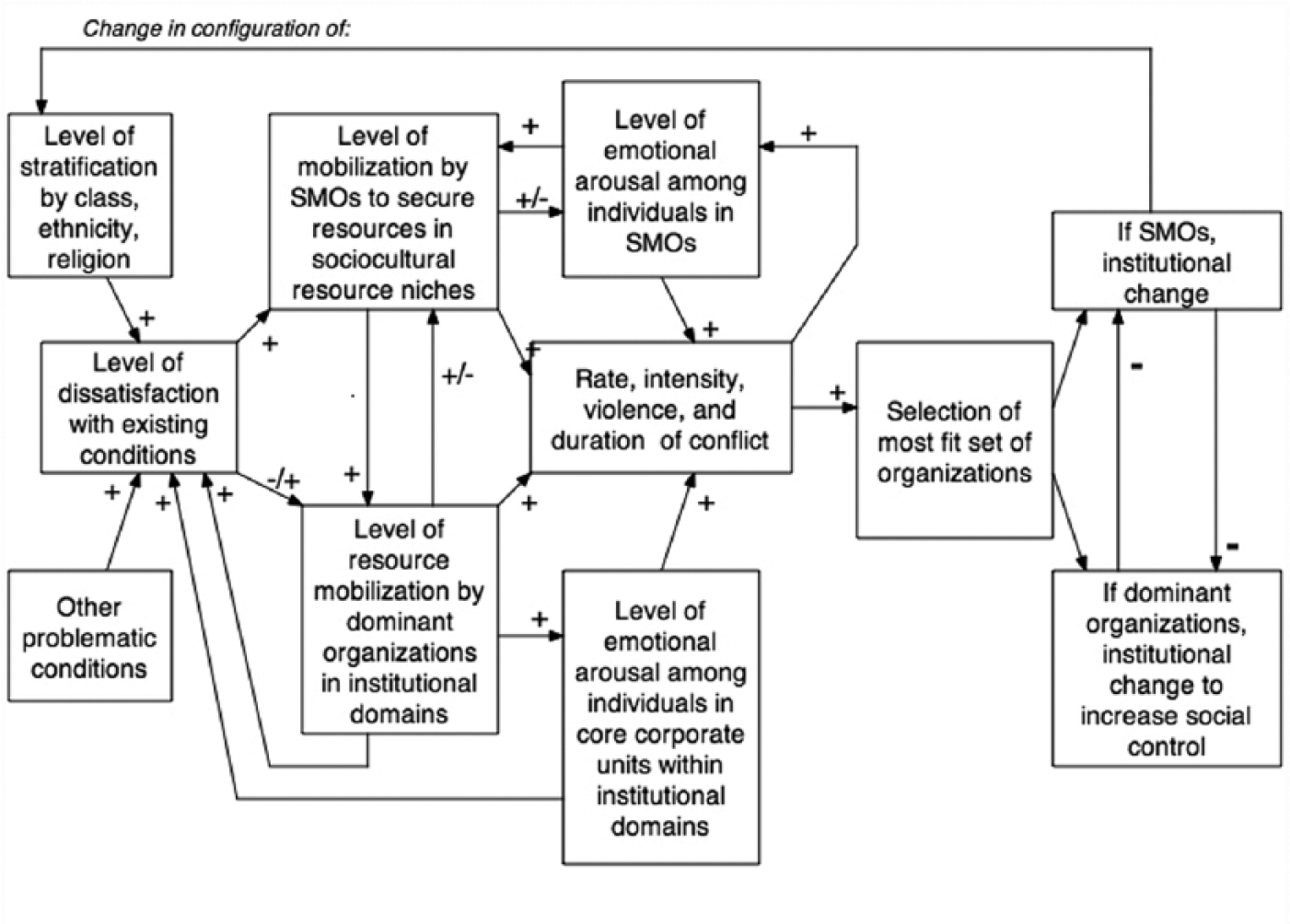
Marxian selection.
Notably, Marxian ideas can be easily blended with organizational ecological arguments in the literature on social movements, and they thereby make clear that Marxian theorizing is still a selectionist argument. The social movement literature emphasizes the formation of social movement organizations (SMOs) that begin to organize in response to grievances against the existing institutional systems in a society (Snow and Soule 2010). 3 Emerging SMOs seek cultural or symbolic resources, often by framing their quest as an effort to realize the general values and moral codes in a society that have not been equally applied or in some way have been abrogated to narrow interests sustained by the existing institutional systems; in addition, they may create these resources where they are unavailable (Summers-Effler 2010; Summers-Effler and Kwak 2015)—a sort of Spencerian entrepreneurship (Abrutyn and Van Ness 2015). Along with these ideological efforts that draw upon cultural resources, SMOs begin to seek a variety of additional resources, including members and volunteers willing to act, material resources such as money, organizational resources such as existing organizations (like churches or unions) and/or their templates for mobilizing individuals, networks with other organizations and with kinships, occupational, and community structures as conduits for recruits, money, and organizational liaisons. They may also seek technological resources that can be employed to recruit members, sustain networks, and broadcast their cause to the larger population. The success of a revolutionary SMO, capable of Marxian evolutionary transformation, is a function of (1) their success in securing sufficient resources and in co-opting or eliminating other SMOs focusing on the same or similar issues, (2) their capacity to reach a large proportion of the population and neutralize potential resistance, and (3) their ability to change institutional structures—for example, political, legal, economic—generating the grievances that initiated the formation of SMOs. At this point, the environment to which many corporate units must adapt has changed, sometimes in ways that affect many diverse types of corporate units in differentiated institutional domains (e.g., economy, polity, law, education, perhaps religions, etc.). Thus, because Marxian selection involves mobilization for institutional change through conflict, it drives the formation of new resource niches and environments to which actors in a superorganism must adapt. Other types of selection can do the same but, in the case of Marxian selection, the evolution of the environments to which actors must respond can be dramatic and rapid.
In anticipating the next section, we should perhaps emphasize that religion has often been at the center of these more Marxian-type processes because it already has achieved what evolving SMOs always seek: (1) a highly moralized belief system that can be used to charge up emotions, (2) a collective memory that roots social ties and obligations in a sacred time, and (3) an existing organizational structure with leaders and a division of labor ready to act and recruit additional resources to pursue conflict. Indeed, religious organizations have advantages over other types of organizations in that their rewards are intangible and long term and, thus, commitment is intensified, especially where a singular, transcendental deity comprises the supernatural (Norenzayan 2013; C. Smith 1996; Stark and Bainbridge 1996). In addition, religious collectives have networks that cross-cut other categories of stratification, as well as other types of corporate units—for example, kin units, neighborhoods, and communities. As we will outline in the next section of the paper, religion is a very appropriate illustration of our effort to visualize four distinctive, although related, types of natural selection. By using a very brief sketch of how these types of selection can add to explanations for the emergence and subsequent evolution of religion, we can hopefully make a stronger case for a revival of evolutionary thinking in sociology.
On the Origins and Evolution of Religion: By Means of Natural Selection
The origins of religion is a question that has been central in the social sciences since their beginnings (e.g., Durkheim [1915] 1995; Morgan [1877] 1996; Spencer 1897), while becoming ever-more prominent in modern psychological and biological evolutionary analysis (Atran 2002; Boyer 2001; Dawkins 2006; Geertz 2013; Norenzayan 2013). From a sociological perspective, the evolution of religion begins with Darwinian selection working on the great-ape-like hominin neuroanatomy to give humans the capacity to imagine a supernatural universe inhabited by beings and forces, coupled with the need to make ritual appeals to the powers of these supernatural forces. Without the rewiring of the basic great-ape brain over the six to seven million years of hominin evolution, religion could never have evolved in human societies. But the capacity and even the propensity or need to engage in religious behavior cannot explain how and why religion became institutionalized in early hunting-gathering societies and, once institutionalized, how it evolved as societies became increasingly larger and more complex. That is, Darwinian evolution driven by natural selection on the neuroanatomy of humans’ hominin ancestors needs to be supplemented by analysis of how Type 1 and Type 2 Spencerian selection institutionalized religion and, then, how Spencerian, Durkheimian, and Marxian selection have all driven subsequent religious evolution. Let us begin, then, with the Darwinian part of the story on the evolution of religion.
Darwinian Selection and the Origins of Religion
The question of why humans are religious is long-standing but, recently, a variety of scholars has sought to develop explanations for its origins (see Geertz 2013 for a compilation). Much of this work has been highly speculative; even that by biologists and social scientists. As such, Turner along with a network of colleagues (Turner et al. 2016) has recently sought to develop a much more robust explanation, emphasizing Darwinian selection on the basic neuroanatomy of extant apes as a proxy for the behavioral capacities of the last common ancestor to present-day apes and the hominin line of evolution leading to Homo sapiens (via a methodology from biology termed cladistic analysis). This analysis turns first to considering the hardwired behavioral propensities of apes, which leads naturally to a comparative analysis of neuroanatomy. By comparing the differences in neurological structures between ape and human brains, we have a good proxy for evidence of what Darwinian selection did to the basic ape brain that the last common ancestor to humans (and present apes) possessed some six to seven million years ago. The habitat of extant great apes remained arboreal, even as the forest began to recede in Africa around 10 million years ago, whereas the habitat of hominins changed, and dramatically so. Hominins along the clade of the human line of evolution were essentially great apes but, unlike current great apes today, they had to adapt to niches in the entirely new habitat: the open-country, predator-ridden African savanna without the protection afforded by the forest. The hominin ancestors of humans were, thus, under intense selection pressures compared with humans’ current ape cousins, and it is the changes wrought by Darwinian natural selection on savanna-dwelling hominins that eventually led to the evolution of Homo sapiens.
Genetically controlled behavioral propensities
To begin, great apes and, in some cases, all primates and most mammals, evidence what appear to be innate behavioral capacities that were wired into the great-ape line during their 40 million year period of evolution in the arboreal habitat. The specific data sources for these behavioral propensities are too numerous to summarize here but can be found in a series of articles that cite in detail the primary sources (Turner 2012, 2013, 2014, 2015; Turner and Maryanski 2015a, 2015b). The basic finding of this large complex of data on primates can be summarized as follows: great apes are visually dominant and are disposed to rely upon the gestures of face and body for rather subtle forms of communication, often organizing instrumental cooperative action; great apes can role take and establish a state of empathy with conspecifics; apes can recognize themselves in a mirror and, when asked, define who they are; apes can calculate reciprocities and fairness in exchanges; apes can read primary emotions almost at birth and can exhibit more variants of primary emotions than any other primate except humans; apes can learn human language and express themselves via visual sense modalities; and apes can use rituals to open and close interactions, while falling into rhythmic synchronization with conspecifics during the course of face-to-face interaction. What is evident from this list is that many of the behaviors of humans in interaction are simply extensions of these behavioral capacities and propensities of great apes. Thus, they were available for natural selection to work on if their elaboration would enhance the fitness of hominins having to adapt (or die out) as the forest shrunk and the new terrestrial habitat began to open up some 10 million years ago.
Great ape social structures
Alexandra Maryanski (1986, 1987, 1992, 1993) conducted a review of the literature on apes and monkeys to determine the nature of their respective social structures and the underlying behaviors that generate and sustain these structures. Among great apes, there are very few strong or permanent social ties. If there is any kind of social structural orientation among great apes, it is to the larger regional population or community that can be as much as 10 square miles. Hence, great apes are not highly social or group oriented.
The weak-tie, nongroup organizational structure for great apes was the result of natural selection working to limit the number of individuals at a given place in the arboreal habitat. Apes had lost out to monkeys in the competition for the core areas of the trees, where there is food, room, and structural support for branches. They were pushed to the terminal feeding areas at the tops of the arboreal habitat where there is less food, less space, and little structural support from thin branches. In such niches, permanent groups would be maladaptive because of these limitations, as large groups of apes would soon exhaust the food supply, causing death by starvation, and/or overwhelm the weak branches, thereby causing death by gravity.
Using these data on social ties and networks, Maryanski conducted cladistic analysis to determine the social ties and network structures of the last common ancestor to contemporary great apes and the hominins on the human line. Cladistic analysis consists of outlining traits that are evident among species that are presumed to have evolved from a common ancestor. If all members of this set of species reveal behavioral capacities, propensities, and similar social structures (as determined by their networks of relations), then it can be assumed that the last common ancestor of these species of great apes and humans also possessed these capacities, propensities, and structures. Variation from these common traits, then, is assumed to have evolved independently among members of the set of related species as they adapted to their somewhat different niches in various habitats. The result of Maryanski’s cladistic analysis was that the last common ancestors of present-day great apes and, for our purposes, the hominin line that broke away from the ape line were virtually solitary. The only strong tie among the last common ancestor, as with contemporary orangutans, was mother-offspring ties, which are virtually universal among mammals. However, this one strong tie would have been severed in the last common ancestor when offspring emigrated away from their natal community and their mother, never to return—just as great apes do today. Otherwise, there are no strong or permanent ties among any conspecifics among orangutans and probably the last common ancestor as well, save for the short-term courtship of females by males during mating (with this tie not lasting more than a couple of weeks, with the male then leaving the female to raise their offspring alone). Therefore, humans are not nearly as social as is often presumed, and no ape, except humans, forms nuclear families much less any other kind of permanent group.
What, then, does the above have to do with the evolution of religion? The answer to this question resides in the selection pressures on hominins as they separated from their last common ancestor with present-day great apes. On one hand, the behavioral propensities and capacities of all great apes briefly listed earlier, coupled with preadaptations for language, would seem to indicate that there was something on which natural selection could “select” if increased sociality would facilitate survival on the savanna. On the other hand, given the fact that nearly every species of ape, save for five, is now extinct, these traits were apparently insufficient to enable great apes to do something that is not very natural to them as a set of species: to form stronger, more permanent social ties among conspecifics and to form more permanent groups with higher degrees of solidarity. Without strong ties or groups, most apes were doomed to extinction on the savanna, and this is exactly what happened. How, then, did hominins survive?
Comparative neuroanatomy
If we compare great ape to human brains, they are very similar and reveal the same basic structures. But there are two significant differences of degree. First, the human neocortex is three times the size of that of a great ape, controlling for the effects of body size. This difference gets most of the press in the social sciences because it accounts for human intelligence and capacities for culture. Equally significant and, in fact, far more significant in an evolutionary sense, is the fact that subcortical areas of the human brain where emotions are generated are, on average, twice the size of those among great apes (see Eccles 1989 for a short review of the precise measurement involved; see also Turner 2000). Natural selection was thus growing the emotion centers of the hominin brain long before significant increases in the size of the neocortex occurred (more on the significance of this fact shortly). Second, the connectivity within the neocortex and older subcortex as well as between these two basic regions of the brain is dramatically enhanced in the human brain, particularly neuro-nets between the prefrontal cortex (where decisions are made) and emotion centers in the subcortex. If there is a “smoking gun” as to what Darwinian natural selection was doing, these comparisons of ape and human brains are strong evidence.
Hence, with hominins, natural selection blindly hit upon a solution that had eluded most species of savanna-dwelling apes: enhance emotions and thereby supercharge those behavioral capacities and propensities listed earlier that would seem to increase sociality. The first language among hominins was likely a visually based “language of emotions” built upon the (1) preadaptations that all great apes have for language and (2) expanded range of emotions that hominins could experience to the point where social bonds and groups could form (Turner 2000). These were, however, laid over the older ape propensity for weak ties and nongroup formations, and so, human groups and, indeed, human social relationships in general are always difficult because they are not hardwired in bio-programmers but, rather, dependent upon emotional neurological capacities that must be invoked and manipulated to forge bonds (Lawler, Thye, and Yoon 2009), a process fraught with potential for breaches and arousal of negative emotions. But these emotional enhancements were just enough to get hominins sufficiently organized to survive and reproduce.
Indeed, the importance of emotions for social life cannot be overemphasized. Consider the number and diversity of interpersonal mechanisms shaped by emotions (Turner 2002, 2007): role taking and role making; verifying status and roles; using gestural rituals for opening, closing, structuring, and repairing breaches during the course of interaction; keying and rekeying of frames; self-presentations and other identity dynamics; interaction rituals generating solidarities through rhythmic synchronization of talk and body language and the emotional entrainment of individuals; emotion management to meet expectations; and assessing relevant status beliefs, norms, and expectations states. Ultimately, natural selection was working to enhance sociality through emotions and, at the same time, generating as many “hooks” for social bonding as is possible.
The neurological and behavioral capacity for religion
There is one more element of neurology that must be introduced: growth in the neocortex would have no fitness-enhancing value without the prior expansion of hominins’ emotional capacities. And, indeed, we can see this fact in the measurement of cranial capacities of hominins over the last 7 million years. There was a modest increase in size from 375−450 cc for great apes to 475−525 cc for Homo habilis (3.0 million years ago), although some now argue that habilis is just an early Homo erectus. In either case, the neocortex—the fountainhead of human culture and language—grew very little for perhaps as much as 7 million years, where, during this same period, the subcortical areas of the hominin brain doubled in size. The neocortex portion of the brain only began to grow significantly with later Homo erectus, and the dramatic variations in the size indicated by cranial casts of Homo erectus suggest that the brain of Homo erectus was about that of Homo habilis at 500 cc at 2.0 million years ago (the same is true in the-yet-undated find of Homo naledi), but by 1.0 million years ago, it was rapidly growing to its maximum size of 950 cc, which overlaps with the lower range of the human neocortex, which is about 1,250−1,350 cc on average.
The reason for this late evolution of the neocortex is that for all animals on earth, cognitions can only be remembered, retrieved, and used in decision-making if they are tagged with emotions. Without emotions attached to cognitions, we cannot remember anything for very long, nor can any animal make a decision (Damasio 1994). Hence, for intelligence to grow, it is necessary for the range of emotional tags to expand so that cognitions can be remembered, stored, and retrieved with nuance, subtlety, and complexity. Hence, intelligent animals are always emotional animals, and because humans are the most intelligent animals on earth, they are also the most emotional.
Religion is a by-product of this wiring for interpersonal bonding; it is not the product or by-product of specific genes or modules, as was once proposed by evolutionary psychologists, but rather religion is a consequence of a much more general process of selecting on a wide range of preadaptations—say, for language facility and a larger palate of emotional states—evident in contemporary apes and certainly the last common ancestor of contemporary apes and humans. It is possible to conclude, then, that religion is little more than an interaction process of animals that can now, with their large neocortex, think about a universe that cannot be seen—dead relatives and ancestors, sacred forces, and supernatural deities—and toward whom it is possible to make highly ritualized appeals to use their sacred powers for their benign intervention in the mundane world. Given the close relationship between the emotion of disgust and the process of morality/moral decision-making (Haidt and Algoe 2004), it is clear that positive and negative rituals oriented toward recently or long-dead relatives or other supernatural forces occurs in much the same way as all interaction among humans occurs: through emotionally charged rituals toward “others” where self, roles, relative status, cultural codes, and all of the other emotionally charged-up interpersonal practices discovered by micro sociology are brought into play to enlist the help or to stave off the punishments of the sacred forces and beings in the supernatural realm, as well as invoke real or imagined beings that come to symbolically represent the group as a mode of communicating about the group (Durkheim 1915 [1995]). It was not a dramatic transformation that made human religion possible, but rather, a simple extension of what Darwinian natural selection did to make hominins increasingly social and group oriented.
The Institutionalization of Religion by Spencerian Natural Selection
At first glance, it would seem on the surface that the capacity for religion would not be very essential to the organization of simple hunting and gathering bands consisting of two basic types of groups: (1) nuclear kinship units within (2) bands of perhaps five to eight nuclear families. Would not the interpersonal mechanisms honed by Darwinian selection be sufficient to keep these groups together? We must remember, however, the results of Maryanski’s review of social bonds and networks among primates, and what these data suggest for the last common ancestor. Even after six to seven million years toward enhancing emotions to increase the permanence of social bonds and groups through emotional attachments, humans are still great apes at their core. If we reflect for a moment on contemporary groups of any sort, and indeed all social relations among humans, they are highly vulnerable to disruption by the same emotional enhancements that allow for social bonds and groups (cf. Abrutyn and Mueller 2014). At the neurological level, negative emotions far outnumber the positive emotions that humans can use during interaction (indeed, four of at least the five primary emotions that have been elaborated by hominin neurology are negative [anger, fear, sadness, disgust] whereas one is positive [happiness]). The result is that maintaining a social relationship and peace within groups is not automatic; rather, it is something that must be worked at, constantly (Boehm 1999). What is true today in groups was doubly so in groups living on the African savanna because disintegration of groups could easily lead to death. And so, as outlined in Figure 5, there were probably Type 1 Spencerian selection pressures emerging along at least the fault lines of regulation and reproduction that may have led early humans to institutionalize religion within kin and band units, and eventually as a distinctive set of corporate units constituting a clearly differentiated or autonomous institutional domain. The same premises shaped the rapid evolution of the Ghost Dance cult among the Sioux: under intense exogenous pressure, and the need for new mechanisms of integration and coordination, a ritual emerged around a set of beliefs that it would make the invincible against the white man’s bullets (Mooney 1973).
Person-level Spencerian selection pressures
As outlined in Figure 5, another type of Spencerian selection pressure comes from the subtle but persistent pressure from collectivities of persons whose need-states are not realized and who, as a result, experience a range of negative emotions. No single person can exert much selection pressure on social structures, but when a population is viewed as a whole, even in the small populations of hunter-gatherers, the emotions and need-states of individuals can line up and create pressures on the formation of particular types of structures that deal with collective emotional states and with common psychological needs. These kinds of Type 1 Spencerian pressures would be particularly intense, it can be hypothesized, in smaller societies like that hunter-gatherer system that dominated human social organization for at least 90 percent of Homo sapiens’ time on the planet. Three clear sources of person-level but also population-level pressures come from (1) anxiety reduction for an animal wired for emotions and with a big neocortex that can think in the past, present, and future, and who are painfully aware of their own mortality; (2) need-states that may be universal among humans as part of their inheritance from apes as this inheritance was modified and perhaps intensified by Darwinian selection; and (3) self-control in micro social contexts. Below, we expand upon each of these person-level, and then turn to macro-level examples of both Type 1 and Type 2 Spencerian selection pressures.
(1) Fear is an emotion that was inherited from reptiles that probably only experienced fear and anger from their only emotion center: the amygdala, which is well over twice as big in humans than great apes, and which, curiously, has areas for pleasure grafted onto this ancient area for fear and anger (Turner 2000, 2013). An emotionally charged-up big brain, with emotions biased toward the negative side, is capable of worrying a great deal about what happens now, in the past, or in the future. Humans appear to be quite deft at satisfying needs by exchanging with each other or using rational, natural solutions whenever possible, but, as Rodney Stark and William Sims Bainbridge (1996) have argued, some problems—such as the anxieties surrounding the dead and impending death—would have no natural solutions. It may well be that the interaction rituals that charge up emotions to forge solidarities (Collins 2004) began to be used to make appeals to supernatural forces to intervene and alleviate what can easily be a wide variety of anxieties, or, as Durkheim suggested, generate powerful forces—such as awe, which is a mixture between happiness and fear—that feel external to each individual’s person and, thus, demand attribution and classification.
(2) Almost all sociological and indeed social theory posits fundamental human needs. For example, symbolic interaction theory sees self-verification as a need that must be met; exchange theory argues for needs for profits that are fair and just; and conflict theories implicitly posit a need for efficacy or control of one’s actions and fate. Many theories view a sense of group inclusion and belonging as fundamental (see Turner 2002, 2007 for reviews). As individuals collectively experience these needs and become emotionally aroused by success or failure in meeting some or all of them, the collective emotional arousal exerts selection pressures on social structures and culture, with new organizational, symbolic, and normative patterns emerging out of rituals and crystallizing into external social reality (Abrutyn 2014b). In turn, these external structural and cultural elements act back on the group as emotions “leap” from the people to objects that come to represent the group and, thereby, become sacred and serve as reminders to all members of their shared identity, history, and sense of destiny.
More generally, we find that religious evolution is very likely to be driven by a real or perceived collective sense of anomie rooted in what Max Weber (1922) observed as the central underlying reason for religion: the need for meaning. Peter Berger (1969:22) famously argued that “the ultimate danger [humans face] is the danger of meaninglessness” born of separation from our group(s), and the seemingly coherent “nomos” religion offers humans acts as the “ultimate shield against the terror of anomie” (Berger 1969:26–7). Anomie, although often vaguely defined, appears in Durkheim’s work in two specific ways: (1) as a chronic sociopsychological state of separation from integrative social bonds and regulative moral order, and (2) as an intense sense of shame when one’s status is rapidly disrupted and social bonds are threatened or believed to be severed. In both cases, the social bond—either its consistent absence or its loss due to failing to meet the other’s expectations—is the center of psychological discomfort. Not surprisingly, the former has been consistently found to be at the root of motivation to convert or join religious movements (Abrutyn 2015b; Almond, Appleby, and Sivan 2003; Lofland and Stark 1965).
(3) Self-control in humans is very much related to two emotions that appear to be unique to Homo sapiens. Apes (and, presumably, the last common ancestor to apes and hominins) do (and did) not experience shame and guilt (Boehm 1999), whereas among humans, these emotions evolved as the neocortex was growing and, with this growth, enabled humans to articulate cultural and moral codes. People experience guilt if they feel that they have violated a moral code, and they will experience shame when they perceive self as not meeting the expectations of others and is deficient. These are very powerful emotions, and they convert social control by others into self-control, where the person is motivated to avoid shame and guilt by meeting expectations and abiding by moral codes, and while moral codes are social codes, concretizing the source of authority—albeit an “invisible” supernatural source—provides a source of sanctioning in societies lacking politically differentiated actors, and an animal capable of experiencing chronic shame and/or guilt will be highly receptive to these potential sanctions and guidance from “on high” supernatural beings and forces.
Macro-level selection pressures
Type 1 Spencerian processes can clearly be seen in cases such as the ancient Israelites (Abrutyn 2014a, 2015a, 2015b). When Assyria destroyed the northern kingdom of Israel, a wave of refugees traumatized by the loss of land and property faced the metaphysical question of why their god(s), Yahweh, had forsaken them (Finkelstein 2001; Schniedewind 2004) but, more importantly, forced them to begin to mobilize in the face of Spencerian selection pressures. From the archaeological record, the kingdom of Judah was a political backwater (Finkelstein 2013), and in all likelihood would not have been structurally or culturally prepared to handle the sudden selection pressures wrought by (1) the tripling of the population brought on by the flood of refugees, (2) the existential crisis faced by the loss of the northern kingdom—both a culturally close cousin and a geographic barrier to Assyrian imperialism, (3) the lack of administrative capacity necessary to handle being thrust into the geopolitical and economic demands of the Assyrian economic system, and (4) the heterogeneity generated by the more cosmopolitan northerners and by the sudden, sharp rise in stratification due to the rapid growth of a feudal system and, thus, a big class of dispossessed folks (Abrutyn 2015b). Needing a scribal class that simply could not have existed given the small size of political complexity in the south, Hezekiah and then later Josiah would have drawn from the human capital of the north; the northerners, carrying traditions that could easily be manipulated to elevate the south over the north, would be authorized to resolve the practical and metaphysical problems the Judean polity faced, which gave them the space and support necessary to innovate.
The result: the outlines of a new religious system. Symbolically, Jerusalem was moved to the center of the cult through the embellishment of the David-Solomon story (Finkelstein 2001), while the Torah and textual authority was elevated over the oral word as Deuteronomy was “found” in a temple cleaning, which placed the written traditions at the center of the king’s and scribes’ claims to power (van der Toorn 2007). Normatively, sacrifice was banned everywhere but the Jerusalem temple as the scribes and king sought to physically and temporally disrupt the local cult, force the people to pilgrimage once a year, and thus forge a national bond (M. S. Smith 1997). Technologically, a new set of scribal hermeneutical techniques that could tolerate contradictions and external changes emerged (Levinson 1997). These techniques lent flexibility to interpretation, reinterpretation, glossing and editing, and redacting extant traditions. It also allowed future scribes, removed from the cultural trauma the northerners felt, to adapt their experiences to future experiences (Abrutyn 2015a)—for example, the Babylonian exile (Smith-Christopher 2002) or, several centuries later, the Roman destruction of the Second Temple and the Second Diaspora (Neusner 2004). All of these innovations were the product of elite-driven innovation. Other examples abound: the elevation of early Christianity by Constantine and the formalization of Catholic dogma and doctrine, the elevation of Confucianism by the Han dynasty and the emergence of the Academy and a standardized philosophy, the elevation of Buddhism by the Mauryan empire in ancient India.
Recall, Type 2 Spencerian selection occurs when a more dominant culture creates pressures on a subordinate, conquered culture to either adapt by assimilating a certain degree of the dominant culture or face absorption into the dominant culture and, thereby, potential extinction. Two brief examples of Type 2 Spencerian selection can be offered. The first is a top-down strategy employed by the Catholic Church to first reduce the conflict between local parishes and their efforts to control personal status issues across Frankish and Germanic villages, and then later to do the same for missionaries in the Americas. The Church recognized the difficulties in converting pagans wholesale, so they canonized local supernatural beings, turning them into patron saints (Sharot 2001). Catholicism evolved in building an extensive set of supernatural beings beside the Trinity, while imposing a centralized ecclesiastic culture on the people who could continue to worship their patron saints—or, the Mother Mary—for immediate needs such as luck, fertility, and the like, while their everlasting salvation shifted to the transcendent Father and his sacrificed Son. A second and, perhaps, more obvious example is Islam (Hodgson 1958). One of the most innovative religions in that it fused with political entrepreneurship early on, Islam spread through conquest. To be sure, the story is not as simple as absorption, as Islamic conquerors allowed the conquered to retain their religious affiliation for a price: substantial taxation. Nevertheless, the rapid spread of Islam led to various local solutions. In Indonesia, for instance, a syncretic blend of Hinduism and Islam persists. In addition, distinct ethno-religious strands emerged differentiating Arab Muslims from many of their North African, Moorist, southeast Asian, and even Persian counterparts. Not merely superficial, each represents the uneasy blending of local cultural elements infused with historical idiosyncrasy and a relatively universal, dominant religious strand carried by conquest. The result is institutional change, but often this change stands in a very uneasy state of high potential for further conflict, and perhaps even conflict revolving around more Marxian dynamics, as are outlined in Figure 7.
Religious Evolution by Durkheimian Natural Selection
Once institutionalized, religion is subject to more Durkheimian selection processes revolving around competition among corporate units organizing communities of worshipers that share beliefs about the supernatural and their ritual appeals to the forces of the supernatural realm. Competition and even open conflict among religious organizations have typified all societies; and while these conflicts can erupt into something more like Marxian selection (see below), they often occur within the constraints imposed by polity, law, and markets—thereby making them more Durkheimian. Moreover, if conflicts become too extreme to be managed within existing institutional systems, they also generate Spencerian selection pressures along the regulatory axes to create mechanisms of social control. Yet, if such control is not forthcoming, then conflicts can morph into Marxian selection in which competition turns violent and, in the process, the entire institutional system of a society comes under attack by organizations willing to challenge the state, economy, and other institutional domains. Thus, the ecology of religious evolution can generate Durkheimian, Spencerian, or Marxian selection pressures. And, as conflicts move into Marxian selection, they still continue to generate Spencerian selection along the regulatory and perhaps reproductive fault line, which eventually will lead to a new pattern of institutionalization of religion among members of a population.
In a penetrating analysis, Mark Chaves (2004:143ff.) has illustrated the ecological dynamics of American religious markets. At the heart of his study are two key resources: money and members (cf. Iannaccone and Bainbridge 2009). Because most congregations rely on their members’ donations for funding, most congregations—besides the small percentage of “mega-churches”—live month-to-month while competing with other churches in resource niches shaped by the size and density of the population from which any given congregation can draw potential members, the population’s socioeconomic factors, and the number of competing churches and, of course, nonreligious secondary associations. These congregations adopt strategies to draw more members and money, such as the provision of social services (e.g., 57 percent of congregations in the United States, comprising 75 percent of the congregants in his survey, claim to provide social services such as food programs).
Where resource niches have reached a real or perceived carrying capacity, two strategies drive organizational evolution. The first strategy is isomorphic: “emergent religions positions themselves relative to already existing groups such that their worship is different, but not too different, from prevailing practice . . . resembling an ecological, evolution of species” (Chaves 2004:151–52); commonly, this strategy is employed by Evangelical sects breaking away from older, more established ones. This strategy provides the church with a marketing message that appeals to maintaining ontological certainty, but encourages greater collective effervescence and, thereby, meaning making in rituals (Collins 2004). That is, its doctrine and dogma fit diffuse cognitive schema, but it advertises decay and anomie as found in highly formalized rituals as a way of providing congregants with a shield against those (Lofland and Stark 1965); moreover, in reducing the “costs” of membership by making worship less formal (e.g., relaxed attire) while making the benefits visceral and affectual, the “terms of exchange” between the supernatural and humans is far more palatable (Stark and Bainbridge 1996).
The second strategy is equally Durkheimian: where the broader population is large, dense, and relatively rich, but cultural forms are fixed, new religious movements “pursue a legitimation and differentiation strategy further out on the leading edge” (Chaves 2004:158), situating “themselves partially in a cultural practice space that already is occupied and partially in a space that is not yet occupied by established religious organizations” (Chaves 2004:155). While we often think of cults as exemplar, Mormonism reflects a sort of middle-ground approach to both strategies. Chaves, importantly, adds a Lamarckian element noting that religious entrepreneurs may make decisions to move further away from established cultural forms because of espoused normative or theological reasons, but ultimately do so as a means of tapping into either a subpopulations’ tastes or preferences, cultivating these tastes and preferences, or in articulating a movement frame that emphasizes the unjust and immoral nature of the existing cultural options, while highlighting the charismatic revelation offered by their symbolic, normative, and organizational innovations.
Yet another form of Durkheimian argument comes from Robert Wuthnow’s (1987) analysis of the “moral order,” which presents an argument for both past and present religious competition. At different points in religious history, especially in the United States, those ideologies that reduce uncertainty and order self, roles, programs, constraints, and social control (perhaps in response to Spencerian selection) are more likely to attract a following, especially if they can, in response to Durkheimian selection pressures, secure viable resource bases (material, networks, rituals, organizational footings, and leadership). Particularly important is the capacity of organizations to gain legitimacy with respect to existing values and legal rules, as well as political authority, while revealing a high number of weakly connected, but flexible, moral codes. Organizations meeting these criteria are more likely to survive in the competition for members.
Religious Evolution by Marxian Natural Selection
At many points in history, and as is certainly evident in the present-day Middle East, competition gives way to more violent conflicts often among sects of the same religion, and equally often, this conflict evolves into a struggle for not only religious dominance but political domination of a society. Certain general conditions work to push the ecology of religious competition toward this more violent societal and intersocietal pattern of religious warfare. Once competition challenges the institutional order in societies or even a system of societies, selection moves to a more Marxian profile and, whether successful or unsuccessful, leads to significant transformation in the institutional systems of a society or, potentially, a system of societies.
In some cases, the violence is aimed at nonreligious elites, such as the Iranian revolution, which radically and qualitatively transformed the Iranian state, replacing the political and economic elite with a religious elite who put sacredness and piety first and power and money second (Moshiri 1991). Similarly, fundamentalism has been the principal source of antisystemic movements across the globe and across religious systems aimed at capitalism, western democracy, Christianity, and other sources of modern “corruption” (Almond et al. 2003). At other times, the aspiring or already positioned religious elite draw political elite into the conflict (or simply benefit from clashes between political elite and upstarts), as the latter’s unique monopoly over violence allows them to destroy competitors. The Pharisees were “selected” in that their unique brand of Judaism, which would eventually become Rabbinic, won out over their central rivals, the Sadducees, as well as others such as the Essenes and Zealots whose religious variants exist only as artifacts; they positioned themselves and adopted strategies meant to accommodate the imperial Roman state, whereas the other factions failed to (Neusner 2004). More recently, the Soviet state used force to wipe out the Orthodox Church (Froese 2009). Although it did not succeed in its goal of total eradication, it did allow other religions to “flourish” in the vacuum.
Commonly, however, violence is unleashed on other religious competitors. Saddam Hussein, and many other Muslim states such as Turkey, have used force in an effort to eradicate the Kurds; in addition, the Sunnis and Shiites are militarized within and between states. Of course, as the Islamic State of Iraq and the Levant (ISIS) has shown, this struggle can be generalized against all groups that do not seem orthodox by whatever criteria are employed. Other historical examples abound. The Catholic Church routinely used force or the threat of force to silence “heretics,” and would have used that force against Luther had he not been protected by a German prince. The Crusades, although not totally driven by religious motives, were driven, in part, by the encroaching presence of Islam on the peripheries of Europe and a growing population of second-born sons with no recourse to status given the dictates of primogeniture (Le Goff 2005). Christian missions, especially during the earliest days of colonialism, sought to convert heathens with force. In the case of the Aztecs, their religion was “wiped out,” becoming a museum religion, while most other indigenous South and Central American religions were not lost, but rather absorbed into Catholicism in ways similar to the adoption of saints when Catholicism encountered Germanic resistance (Sharot 2001).
Conclusion
Our hope is that by using religious evolution as a protracted but still very rushed example, the merits of emphasizing selection as a basic force become evident (see more extensive book-length analysis in Turner et al. 2016). But, as we have stressed, Darwinian selection on individual organisms in evolving populations of organisms is only one type. Emphasis on Darwinian selection pulls sociological theory into biology and the analysis of how “human nature” evolved, but in a much more sophisticated way than early speculations on human nature—as we sought to demonstrate with our brief review of hominin evolution under Darwinian selection pressures. Type 1 Spencerian selection allows us to take what was useful about functionalism and throw the rest out, and thus enables us to see why particular institutional domains such as religion emerged in the first place; that is, as a response to the selection pressures emanating from the fault lines of all organizational structures and from the failure of larger numbers of individual to meet basic human need-states, and negative emotions aroused such is the case. Type 2 Spencerian selection adds a geopolitical dimension to religious evolution by emphasizing that intersocietal conflicts and conquest also lead to the flow of the culture and institutional systems into conquered societies that won the war. And, particularly, would such be the case because religion is so often critical in legitimating political domination. Durkheimian selection blends organizational ecology with the analysis of social movements into a useful model of competition among corporate units for resources within niches that are, to varying degrees, regulated by markets, law, and polity. Institutional systems evolve as the mix of corporate units within a domain changes as a result of this regulated competition. By adding a more Marxian form of ecological analysis, it becomes possible to understand how more revolutionary movements get started and move to change institutional systems within and even between societies. And, with a certain irony, it appears that religion in the history of societal evolution has been more “revolutionary” in this ecological sense than social class. For, unlike any other organizational system in societies, religion has already achieved what most revolutionaries and social movement leaders see: an organization or set of organizations with a steady input of all necessary resources, with a highly moralized ideology that can be adapted to more secular causes, a set of ritual practices that arouse emotions and reinforce moral codes, and an existing leadership structure and division of labor that can initiate or be pulled into conflicts that often turn violent because they are so moralized. Whether evolution change by Durkheimian selection or more revolutionary change by Marxian selection, institutional systems and the mix of corporate units and mechanisms integrating these units change, as can be seen today in the violent conflicts of the Middle East.
While we have endeavored to demonstrate how other types of selection may help to explain the evolution of religion, both through the creation of the necessary neuroanatomy as well as in sociocultural institutionalization, we believe these processes can be brought to bear on other types of sociocultural formations. For instance, Seth Abrutyn and Kirk Lawrence (2013a; Abrutyn and Lawrence 2010) have applied Spencerian and Durkheimian selection models to the evolution of the polity, exploring the emergence of the earliest autonomous states. In addition, Seth Abrutyn (2009) has suggested that Spencerian processes can better account for the evolution of the unique western legal system during the Gregorian Reformation in the eleventh century C.E. While both of these examples lack systematic treatment, it is not difficult to discern the potential power these models have in specifying how institutional spheres evolve, or, as S. N. Eisenstadt’s (1964; cf. Abrutyn and Van Ness 2015) work highlights, how institutional entrepreneurs contribute to non-Darwinian selection.
In sum, selection, as we have outlined it, does not explain everything, and may well prove to be less useful than alternative theories in explaining evolutionary processes. But, sociologists have been making selectionist arguments since the very beginning of the discipline, as is evident in the analysis of organizational ecology and social movement literatures. We have just made these arguments more explicit, while indicating that by emphasizing selection processes that are “natural” to superorganisms, a continuity is created with biological sciences but, unlike many current efforts to bring biology into sociology, biologically inspired ideas are adjusted to the fact that sociology seeks to explain the dynamics of populations of superorganisms, revealing many layers of social structure and culture, all of which operate under selection dynamics that are different than those posited by the Modern Synthesis in Biology. And to emphasize, again, the crucial point: these other types of selection dynamics are just as natural as those Darwinian selection dynamics used to analyze populations of organisms.
Footnotes
Acknowledgements
The authors would like to thank the editor and reviewers for important suggestions and criticisms on previous drafts.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.