
Abstract
Background:
The emergence of highly pathogenic avian influenza A (H5N1) clade 2.3.4.4b in dairy cattle and human cases raises urgent pandemic concerns. A critical question is whether seasonal influenza vaccines elicit cross-reactive immunity against this novel zoonotic strain, potentially contributing to pre-existing population immunity.
Methods:
We combined structural bioinformatics and serological analysis. Hemagglutinin (HA) protein sequence and structural conservation were assessed between World Health Organization WHO-recommended 2024–2025 vaccine strains (H1N1 A/Victoria/4897/2022, H3N2 A/Thailand/8/2022) and bovine H5N1 (A/Texas/37/2024). Cross-reactive antibodies were measured in serum from 46 vaccinated individuals using ELISA against A/Texas/37/2024 HA. Endpoint titers were the highest reciprocal dilution with absorbance >2.1-fold background. Statistical analyses included Pearson correlation (age, dose, time) and Wilcoxon rank-sum/Chi-squared tests (group comparisons).
Results:
Structural analysis revealed 79.3% amino acid identity in the HA2 subunit between H1N1 and H5N1, with conserved epitopes in the stalk domain. Serologically, 41.3% (19/46) of vaccinated individuals had cross-reactive HA-binding antibodies with titers ≥1,280. No significant associations were found with sex, vaccine type, brand, or number of doses. A significant positive correlation existed between age and antibody titer (Pearson’s R = 0.51, p < 0.001); individuals over 60 years had higher titers than younger groups.
Conclusions:
Seasonal influenza vaccination is associated with cross-reactive HA-binding antibodies against bovine H5N1 clade 2.3.4.4b in a substantial proportion of individuals, with responses increasing with age. This suggests pre-existing immunity from vaccination or prior exposures may influence responses to this zoonotic threat. However, the functional neutralizing capacity and protective efficacy of these antibodies are unproven. These findings highlight the potential immunological footprint of current vaccines against emerging strains and support further investigation into cross-protection. They also reinforce the importance of conserved HA epitopes as targets for next-generation universal influenza vaccines.
Introduction
In March 2024, the highly pathogenic avian influenza (HPAI) subtype A H5N1, clade 2.3.4.4b, were identified in dairy cattle within the United States (Baker et al., 2025; Caserta et al., 2024), leading to an infection in a farm worker in Texas (Uyeki et al., 2024). Following this initial detection, the virus has been confirmed in dairy farms across 16 states. Up until July 7, 2025, a total of 71 human cases of H5N1 infection have been reported in the country, with two fatalities recorded among these cases (CDC US, 2025). These cases have sparked concerns over a potential public health emergency, given the risks of the virus spreading to humans via infected cattle, milk, and the food supply chain, as well as the possibility of human-to-human transmission (Lin et al., 2024; Rosenke et al., 2025). Vaccination may control its spread, but the effectiveness of existing H5N1 vaccines against circulating strains, especially clade 2.3.4.4b, remains uncertain. Consequently, the development of effective vaccines and therapeutics targeting the HPAI H5N1 is deemed a public health priority.
Receiving the influenza vaccine recommended by the World Health Organization (WHO) prior to the start of the flu season is currently the foremost strategy for preventing seasonal influenza. Antibodies directed against hemagglutinin (HA), the major surface glycoprotein of the influenza virus, are essential for providing immunity against infection (Subbarao and Joseph, 2007). Broadly neutralizing antibodies that target specific epitopes on the influenza virus HA have the potential to offer near-universal protection against influenza virus infections (Paules et al., 2017; Sun et al., 2024). Particularly, antibodies targeting multiple conserved epitopes on HA, such as membrane-proximal anchor epitope and a distinct site overlapping the receptor binding site, can induce broadly cross-reactive antibodies effective against H1 and H5 strains (Guthmiller et al., 2022; Li et al., 2022, 2023). This capability underscores their significant role in providing comprehensive immunity against diverse influenza strains. Therefore, due to the emergence of this HPAI H5N1 virus and its widespread circulation in birds and mammals (including dairy cattle), with the potential to spread to humans, it is important to determine if any of the licensed influenza vaccines elicit antibodies that are cross-reactive against the currently circulating HPAI H5N1.
In this study, we conducted a systematic comparison of the HA sequence composition and structural conservation between current influenza vaccine strains (A/Victoria/4897/2022 [H1N1] and A/Thailand/8/2022 [H3N2]) (WHO, 2024) and the HPAI H5 N1 strain (A/Texas/37/2024). Additionally, serum samples were collected from 46 vaccinated individuals. Using the ELISA method, we analyzed the binding characteristics of antibodies in these sera to the HA of A/Texas/37/2024, aiming to evaluate the presence of potentially cross-reactive antibodies. Understanding the cross-reactivity of antibodies generated by current vaccines against emerging HPAI H5N1 strains is crucial for assessing public health preparedness and guiding the development of more broadly protective vaccines. This research contributes vital insights toward mitigating the threat of potential influenza pandemics by enhancing our vaccine strategies.
Materials and Methods
Ethics statement
Ethics approval was granted by the Youjiang Medical University for Nationalities Ethics Committee (the ethics code is 2025011601). This study was performed in strict accordance with the recommendations described in the principles of the Declaration of Helsinki. All patients or the parents of minors were provided with an information notice explaining the nature and purpose of the collected data. In cases where a patient or parent objected to data usage, the corresponding data were excluded from the research database.
Study design and data collection
We collected serum samples from healthy individuals who had received the influenza vaccine, including trivalent influenza split vaccine (SINOVAC) and quadrivalent influenza split vaccine (HUALAN and SINOPHARM), from the affiliated hospitals of Youjiang Medical University for Nationalities. Exclusion criteria for the samples are as follows: (1) no symptomatic influenza infection in the past 3 months; (2) no immunodeficiency diseases or use of immunosuppressive medications; (3) vaccination against influenza between January 1, 2024, and December 31, 2024. Meanwhile, we anonymously collected basic information from volunteers, including gender, age, number of doses received, and vaccination dates.
ELISA
HPAI H5N1 (A/Texas/37/2024) HA-specific antibodies were determined in sera by ELISA. Briefly, ELISA plates were coated overnight with 2 μg/mL of HA (H5N1 A/Texas/37/2024) protein in 0.05 M carbonate–bicarbonate buffer, pH 9.6, and blocked with 5% fat-free milk in PBS. Serum samples were first diluted 20-fold and then serially diluted two-fold before being added to each well. Plates were incubated with goat anti-human IgG-HRP (1:5,000) antibody (Bioss, USA) for 1 h at 37°C and developed with 3,3′,5,5′-tetramethylbenzidine substrate. Reactions were stopped with 2 M HCl, and the absorbance was measured at 450 nm using a microplate reader (PerkinElmer, USA). The endpoint titers were defined as the highest reciprocal dilution of serum to yield an absorbance > 2.1-fold of the background values. Antibody titer below the limit of detection was determined as half the limit of detection.
HA similarity analysis
The HA protein sequences of A/Victoria/4897/2022 (H1N1), A/Thailand/8/2022 (H3N2), and A/Texas/37/2024 (H5N1) were obtained from the NCBI database (https://www.ncbi.nlm.nih.gov/). Multiple sequence alignment were performed using Clustal W v2.0 (Larkin et al., 2007) to calculate the similarity between sequences. Conservative sequence analysis and visualization were conducted using ESPript v3.0 (Robert and Gouet, 2014). Similarity analysis of the tertiary and quaternary structures of the HA protein were conducted using UCSF ChimeraX (Meng et al., 2023), with the H5 hemagglutinin from the HPAI H5N1 virus (A/Texas/37/2024) serving as the reference (https://www.rcsb.org/structure/9DIP).
Statistical analysis
Data analysis was performed using R v4.4.0. Descriptive statistical analyses were calculated using gtsummary v2.0.4 (Sjoberg et al., 2021). The inter-group difference analysis of endpoint titers for HA cross-reactive antibodies in serum was conducted using the Wilcoxon rank-sum/Chi-squared tests, and the correlation analysis was performed using the Pearson method. All plots were generated using ggplot2 v3.5.1.
Results
High amino acid similarity of the HA2 between HPAI H5N1 and H1N1 virus
To assess the possibility of cross-reactive antibodies being generated between the seasonal influenza vaccine [A/Victoria/4897/2022 (H1N1) and A/Thailand/8/2022 (H3N2)] and A/Texas/37/2024 (2.3.4.4b clade bovine H5N1), we performed identity analysis on the primary, secondary, tertiary, and quaternary structures of the major immunogenic protein HA from both the influenza vaccine strains and the 2.3.4.4b clade bovine strains. The analysis results indicate that the HA protein of HPAI H5N1 has a higher amino acid sequence similarity to the HA protein of H1N1. The full sequence identity value is 61.2%, with the HA1 subunit showing an identity of 51.3% and the HA2 subunit exhibiting a sequence identity as high as 79.3% (Fig. 1A). The secondary and tertiary structure alignment results show that there are two β-sheet-rich conserved regions at the N-terminus and C-terminus of the HA1 subunit (residues 17–84, 290–337) (Fig. 1B,C), while the HA2 subunit contains several conserved α-helices (α4–α7) and β-sheets (β30–β34) (Fig. 1B,D). Spatial structure identity analysis of the HA protein indicates that the HA proteins of the H1N1 and HPAI H5N1 strains may share multiple conserved antigenic epitopes, particularly in the neck region of the HA protein (Fig. 1E).
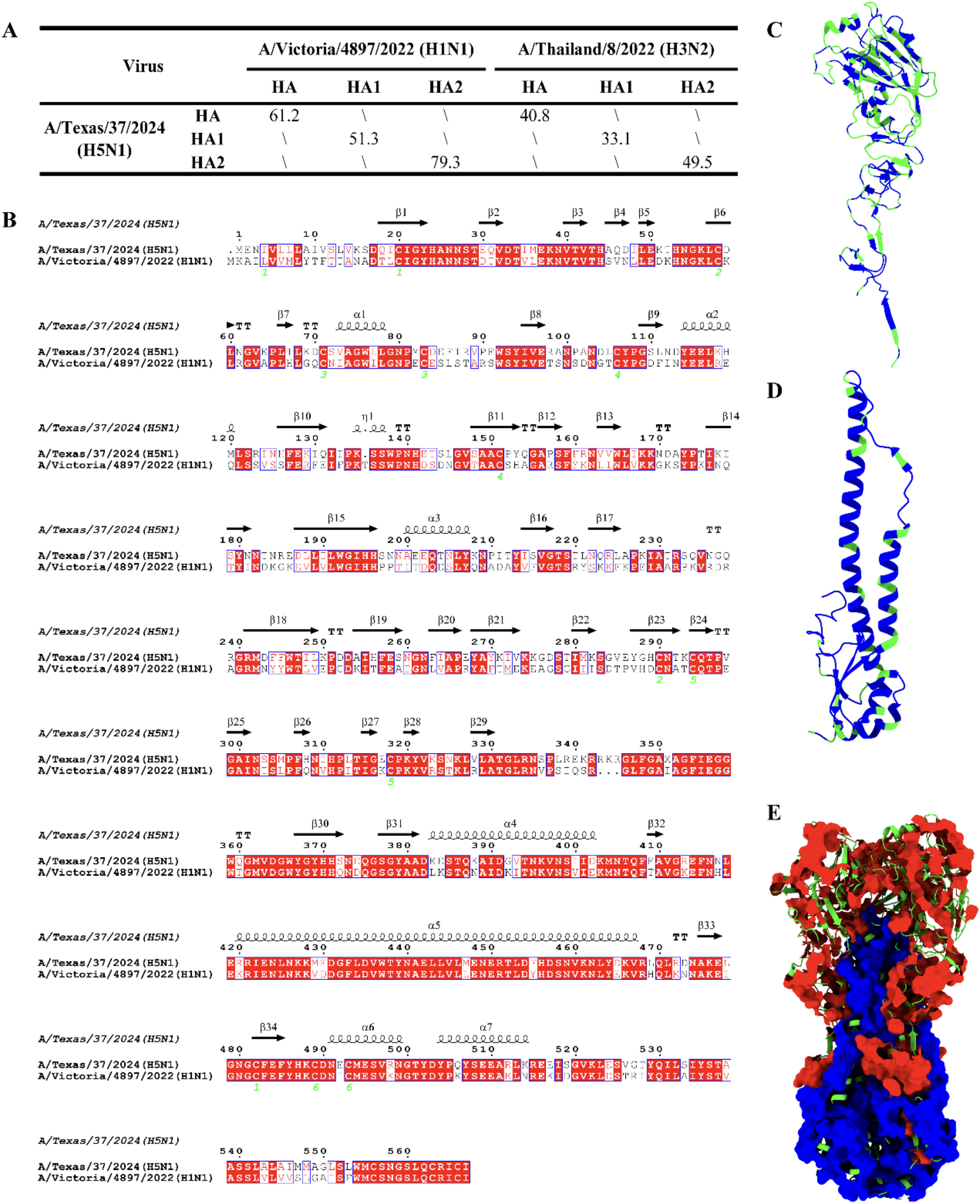
Conserved sequence of HPAI HA and A/Victoria/4897/2022 HA may elicit cross-react antibodies.
Characteristics of the study population
To evaluate whether the present influenza vaccines from A/Victoria/4897/2022 (H1N1) and A/Thailand/8/2022 (H3N2) strains generate cross-reactive binding antibodies against the HPAI H5N1 strain, we analyzed postvaccination serum samples collected from 46 individuals who were inoculated with the influenza vaccine (Table 1). Individuals were vaccinated with either trivalent or quadrivalent split vaccines, with both vaccines containing the Group 1 influenza strains A/Victoria/4897/2022 (H1N1) and A/Thailand/8/2022 (H3N2). The primary populations were individuals under 19 years old and over 54 years old (44/46, 95.65%). Females accounted for 65.22% of the participants. The majority had received one dose of the vaccine (30/46, 65.22%), while some individuals had been vaccinated more than five doses. ELISA evaluation showed that 41.30% (19/46) of the individuals had an HPAI H5N1 HA binding titer of 1,280 or higher in their serum.
Characteristics of Individuals Included in this Study

Pearson’s Chi-squared test; Wilcoxon rank sum test; Fisher’s exact test.
Antibodies of vaccinated individuals cross-react with the HA protein of the HPAI H5N1 virus
To understand the characteristics of cross-reactivity between serum antibodies in vaccinated individuals and the HA protein of the HPAI H5N1 virus, as well as related factors, we conducted various inter-group difference analyses and correlation analyses. The endpoint titers of serum antibodies against the HA protein were mainly distributed above 320, with the highest titer reaching up to 2,560 (Fig. 2A). There was no significant difference in endpoint titers among different genders, trivalent and quadrivalent vaccines, varying doses, or different brands (Fig. 2B,C, 2D, 2F). The binding antibody titers of serum HA proteins in individuals under 9 years old and those aged 9–18 were significantly lower than in individuals over 60 years old (Fig. 2E). Correlation analysis showed a trend toward a negative correlation between the number of vaccine doses received and the endpoint titer (R = −0.24, p = 0.11) (Fig. 2G), while age showed a significant positive correlation with serum HA binding antibody titers (R = 0.51, p < 0.001) (Fig. 2H). There was no significant correlation between vaccination time and serum HA binding antibody titers (Fig. 2I). These results suggest that individual age may be an important factor affecting the production of cross-reactive antibodies against HPAI H5N1 HA.
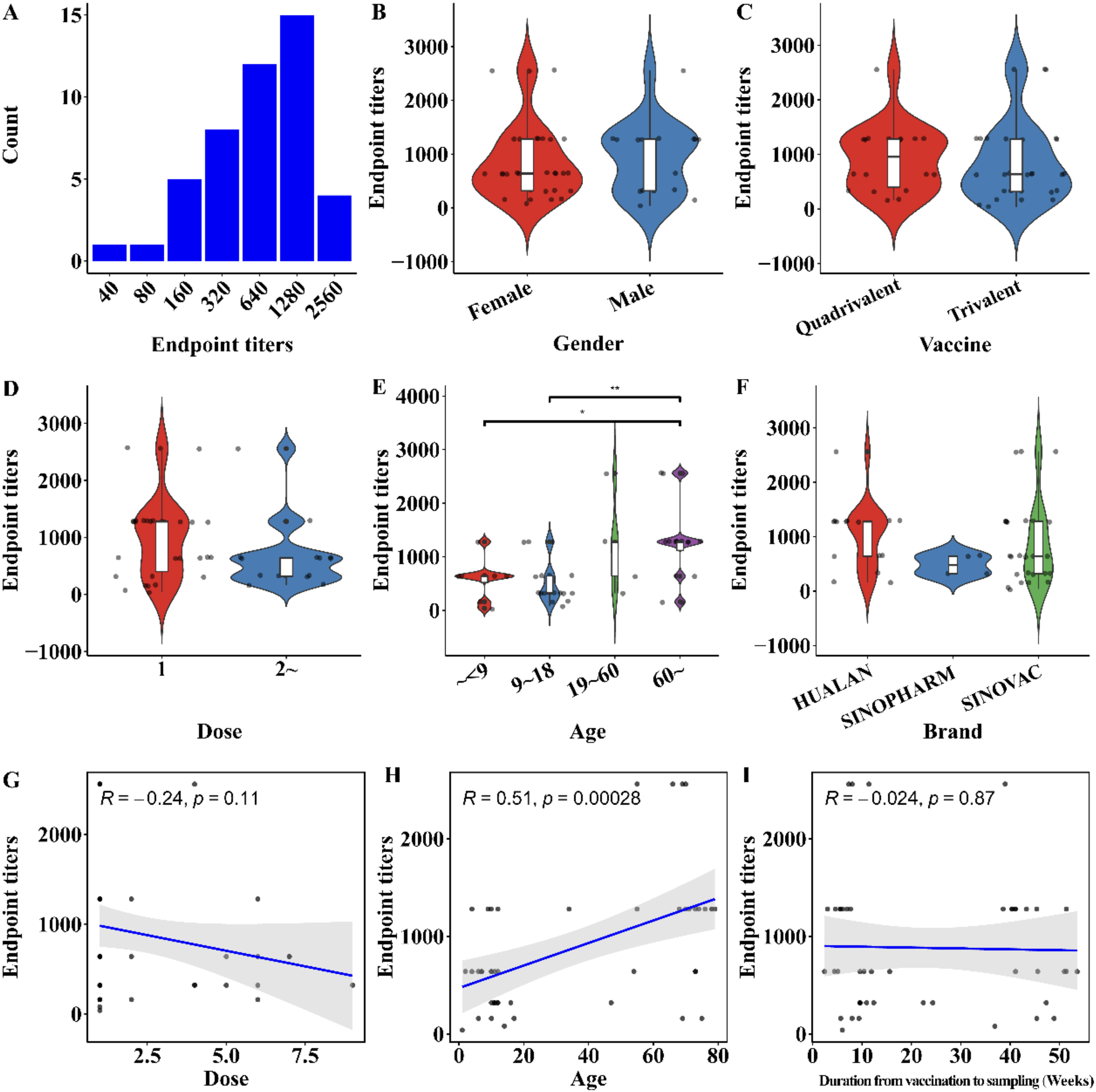
Antibody cross-react with HA of HPAI H5N1 after vaccination of individuals with season influenza vaccines.
Discussion
Historically, newly emerging HPAI virus strains that occasionally transmit to humans have resulted in significant morbidity and mortality. This is largely due to the absence of pre-existing antibodies against these viruses in human populations. Specifically, clade 2.3.4.4b HPAI H5N1 viruses, which have been detected in various mammalian species including domestic animals such as cattle, have also caused infections in humans (Peacock et al., 2025). Official reports document a total of 70 human cases, with one reported fatality. Human infections attributed to exposure to infected poultry or dairy cattle have exhibited a wide spectrum of clinical presentations, ranging from mild conjunctivitis to severe and potentially fatal pneumonia (Garg et al., 2025; Jassem Agatha et al., 2025). Previous studies have demonstrated that certain H1, H3, and H5 subtype influenza viruses share common epitopes capable of inducing cross-reactive neutralizing antibodies. These antibodies can prevent viral entry or mitigate disease severity (Li et al., 2022; Uno and Ross, 2024). Given this, it is crucial to evaluate whether the currently WHO-recommended vaccines, primarily targeting H1N1 and H3N2, can elicit antibodies that are cross-reactive with HPAI H5N1 2.3.4.4b. Such an assessment is vital for informing effective influenza control strategies. Our analysis of the sequence and structural conservation of the HA protein revealed approximately 80% sequence identity between the HA2 subunits of H1N1 (A/Victoria/4897/2022) and HPAI H5N1, along with multiple potential shared epitopes within their three-dimensional structures, particularly in the stem region. These conserved regions suggest a molecular basis for the induction of cross-reactive antibodies against HPAI H5N1 HA through H1N1 influenza vaccines.
Our investigation into the presence of HPAI H5N1 HA protein cross-reactive antibodies in the serum of influenza vaccine recipients revealed that 39.58% individuals exhibited notably high levels of these antibodies (endpoint titers ≥ 1280). This finding indicates that vaccines currently administered for influenza can induce cross-reactive antibodies against H5N1 in vaccinated individuals. Such a discovery is significant, as it suggests potential broader protection conferred by existing vaccines beyond their primary targets.
Interestingly, our data also suggest a trend toward decreasing cross-reactive antibody titers with an increase in the number of vaccine doses received by individuals. This observed trend may reflect the complex dynamics of immune memory upon repeated antigen exposure (Arevalo et al., 2020; Lee et al., 2019). As a result, repeated vaccinations may not always enhance or maintain high levels of cross-reactive immunity against divergent strains such as H5N1. Further research is warranted to elucidate the mechanisms behind this observation and to determine optimal vaccination strategies that maximize cross-protection.
Our observation that individuals over 60 years old exhibit significantly higher HPAI H5N1 HA antibody titers compared with those under 18 years of age aligns with the broader understanding of age-related differences in immune responses. This finding is particularly significant given that younger individuals have been reported to suffer higher mortality rates from H5N1 infections (Cowling et al., 2013; Qin et al., 2015). The presence of more abundant cross-reactive antibodies in older adults can be partly attributed to the immune imprinting effect, where individuals born before 1968 were exposed to H1N1 and H2N2 strains during childhood, potentially conferring enhanced resistance to H5N1 through pre-existing immunity (Gostic et al., 2016). In contrast, children and young adults, who have had fewer early exposures, might benefit significantly from timely influenza vaccination, potentially providing them with crucial protection during an H5N1 outbreak. Furthermore, our analysis revealed that the HA stalk region shared between H5 and H1 subtypes contains numerous conserved epitopes. These conserved regions are crucial for inducing stalk-reactive antibodies. Previous studies have demonstrated that these stalk-reactive antibodies, induced by historical exposures to H1N1 viruses, can provide a degree of cross-protection against divergent influenza strains, including H5N1 (Garretson et al., 2025; Nachbagauer et al., 2016; Yassine et al., 2018), which is similar to our findings. This underscores the importance of the HA stalk as a target for broadly protective vaccines. Understanding the basis of this enhanced response in older individuals may provide insights into developing more effective vaccines tailored to different demographics.
In conclusion, our findings highlight the potential for current influenza vaccines to elicit cross-reactive antibodies against HPAI H5N1, offering hope for broader protection against emerging strains. However, the nuanced relationships between vaccine dose frequency, age, and antibody response underscore the complexity of influenza immunity and the need for further studies to optimize vaccine formulations and schedules. Given the significant implications of these findings, it is crucial to emphasize the importance of widespread vaccination. Strengthening surveillance systems, increasing the transparency of data sharing, and fostering international cooperation are essential steps toward enhancing global preparedness against HPAI H5N1. By working together and ensuring that accurate and timely information is available, we can better protect public health and respond effectively to emerging infectious diseases.
Limitations
Our study has several key limitations: First, the analysis was conducted on a small sample size of 46 serum samples, which may limit the generalizability of our findings. This restricted sample set might not fully represent the broader population’s immune response variability. Second, although ELISA confirmed the presence of cross-reactive antibodies against HA of HPAI H5N1, it cannot determine their neutralizing capacity or protective efficacy. Due to experimental constraints, virus neutralization assays were not performed. These functional studies are essential to assess the actual antiviral capabilities of the detected antibodies and should be included in future research. Finally, we were unable to ascertain the proportion of cross-reactive antibodies derived from early antigen exposure versus those induced by vaccination. This uncertainty would affect the accuracy of our results.
Conclusion
Our findings demonstrate that seasonal influenza vaccination, particularly with H1N1-containing vaccines, can induce antibodies in a substantial subset of individuals (approximately 40%) that cross-react with the HA protein of the emerging bovine H5N1 clade 2.3.4.4b virus. This cross-reactivity is supported by a high degree of sequence and structural conservation, particularly within the HA2 stalk domain, between the H1N1 vaccine strain and the bovine H5N1 strain. Notably, the levels of these cross-reactive binding antibodies were significantly correlated with increasing age, suggesting a role for prior immune exposures and antigenic imprinting in shaping this heterosubtypic response. While these results provide evidence of a pre-existing immunological footprint against this zoonotic H5N1 variant within the vaccinated population, it is crucial to emphasize that our study only assessed antibody binding, not neutralizing or protective capacity. Therefore, the extent to which these cross-reactive antibodies confer actual protection against H5N1 infection or disease remains unknown and requires validation through functional assays, such as virus neutralization tests. Nonetheless, this study highlights the potential for existing seasonal vaccine platforms to contribute to broader immune preparedness against emerging pandemic threats by eliciting responses against conserved viral epitopes. It underscores the importance of considering individual factors such as age and immune history in pandemic risk assessment and vaccine strategy. Future research should prioritize evaluating the functional activity of these cross-reactive antibodies and investigating whether seasonal vaccination can modulate disease severity in the event of H5N1 exposure. Enhancing global surveillance and fostering international collaboration remain paramount for effectively monitoring and responding to the evolving threat of highly pathogenic avian influenza.
Authors’ Contributions
Y.L. and D.C. analyzed the data, and drafted and revised the article. Xinyue Z., Xuan Z., and Xiaoying Z. performed the antibody detection experiment. L.T. and Z.P. collected and processed the samples. Z.H, J.L., and D.C. supervised the study and approved the final article. All authors have agreed to the published version of the article. All authors read and approved the final article.
Footnotes
Acknowledgments
The authors thank Dr. Tianjiao Hao (National Institute of Biological Sciences, Beijing) for the generous gift of HPAI bovine H5N1 (A/Texas/37/2024) HA protein. They also thank for the support of Construction of Key Clinical Specialties in Guangxi (Guiweiyifa [2022]-NO.21) and the Key Laboratory of Molecular Pathology of Guangxi.
Author Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This study was supported by the National Natural Science Foundation of China (62162063, 32560016).
Availability of Data and Material
All data have been presented in this article.