
Abstract
Natural killer (NK) cell activation and function are controlled by highly complex transcriptional programs. To leverage NK cells against human immunodeficiency virus and other diseases, a more complete understanding of signaling and metabolic dynamics is needed. In this study, we utilize a rhesus macaque (RM) model to define the transcriptomic profile of NK cells during acute lentiviral infection and to map the changes in this profile as infection progresses to the early chronic phase. Using a novel summary gene set for NK cell cytokine stimulation, we report synchronous metabolic and signaling transcriptional patterns that underlie established features of NK cell dysfunction. Assessing gene expression induced by the SARS-CoV-2 challenge allowed us to confirm a set of core conserved gene pathways central to RM NK cell activation across diverse acute viral contexts.
Body
Due to their dominant role in combating most viral infections, natural killer (NK) cells are a critical component of the early immune response to human immunodeficiency virus (HIV) and the related simian immunodeficiency virus (SIV). 1 Before manifesting as a systemic chronic illness, HIV/SIV evades innate and adaptive immune defenses at the site of infection and continue to withstand immune pressures throughout a symptomatic acute phase that lasts several weeks, during which the virus establishes itself within CD4+ T cell reservoirs. 1 An understanding of NK cell signaling dynamics during the narrow eclipse phase of early lentiviral infection could enable the successful leveraging of NK cell potency and may benefit the development and improvement of preventative and therapeutic HIV modalities. 1
NK cells are activated by several cytokines present during acute infection, including interleukin (IL)-12, IL-15, IL-18, IL-21, tumor necrosis factor (TNF)-α, and type I interferon (IFNα/IFNβ). 2 Type I interferon in particular plays a major role in limiting the virus before the chronic phase of HIV, with abundance during the pre-chronic phase negatively correlating with virus loads. 3 Activated NK cells function by producing additional antiviral cytokines (IFNγ, TNF-α) and engaging in direct cell killing via antibody-dependent cellular cytotoxicity, killing of infected cells that downregulate MHC, NKG2D-mediated killing via stress ligand upregulation, or engagement of natural cytotoxicity receptors. 2 Importantly, activation requires a complex metabolic remodeling process. NK cells increase glycolysis and mitochondrial capacity during acute infection downstream of the master regulator mTORC1 and its control of the transcription factor c-MYC, 4 a process initiated by the uptake of leucine and other amino acids recognized by mTORC1. 5 NK cells, which display dysregulated metabolism due to overactivation or nutrient depletion, as indicated by mitochondrial fission and diminished glycolytic capacity, exhibit attenuated effector responses.4,6
While the acute phase primes NK cells for viral clearance, chronic lentiviral infection induces a dysfunctional phenotype associated with mitochondrial fission and attenuated oxidative phosphorylation. 6 This dysfunctional state is at least partially attributable to persistent signaling by type I interferon, despite its beneficial role in early stages of infection. 3 Meanwhile, IL-15 has been shown to reverse mitochondrial dysfunction induced by chronic HIV and restore NK cell function in vitro via the mTORC1 pathway. 6 The mechanisms underlying cytokine activation and exhaustion are evidently complex, necessitating the use of broad expression patterns to untangle the dynamic factors that drive this dichotomy.
To better understand NK cell transcriptional dynamics across different stages of lentiviral infection, we performed RNA-seq on bulk-sorted peripheral NK cells from two cohorts of rhesus macaques (Macaca mulatta; RM) infected with distinct SIV strains via different routes. One cohort received SIVmac239 intravenously (IV), and NK cells were sorted as previously reported. 7 The second received SIVmac251 intrarectally (IR), as reported, 8 and NK cells were sorted using the same methodology as in the SIVmac239 cohort. 7
For several time points per SIV-infected RM cohort, we performed gene set enrichment analysis (GSEA) to assess enrichment of Hallmark gene sets post-infection (p.i.) compared with pre-infection (Fig. 1A,B). We found that acute SIV infection was associated with consistent peripheral NK cell transcriptomic signatures related to interferon-stimulated genes (ISGs), proliferation, and metabolism (Fig. 1A,B). Hallmark gene sets for MYC and mTORC1 signaling were enriched on days 7 through 14 p.i. in SIVmac239 IV, but not in SIVmac251 IR, possibly owing to differences in rates of disease progression between routes of infection (Fig. 1B). Type I interferon signaling was consistently upregulated across SIV cohorts following day 3 p.i., a trend maintained in chronic timepoints. Notably, Hallmark gene sets for oxidative phosphorylation, MYC signaling, and mTORC1 signaling were significantly negatively enriched at both day 70 and day 74 p.i. in SIVmac251 IR compared with pre-infection, providing evidence for a transcriptional program underlying NK cell dysfunction during chronic disease. Damaging reactive oxygen species (ROS) represent another established component of NK cell mitochondrial dysregulation, 9 and it should be remarked that genes associated with ROS control were downregulated in the chronic phase compared with NK cells pre-infection (Fig. 1B).
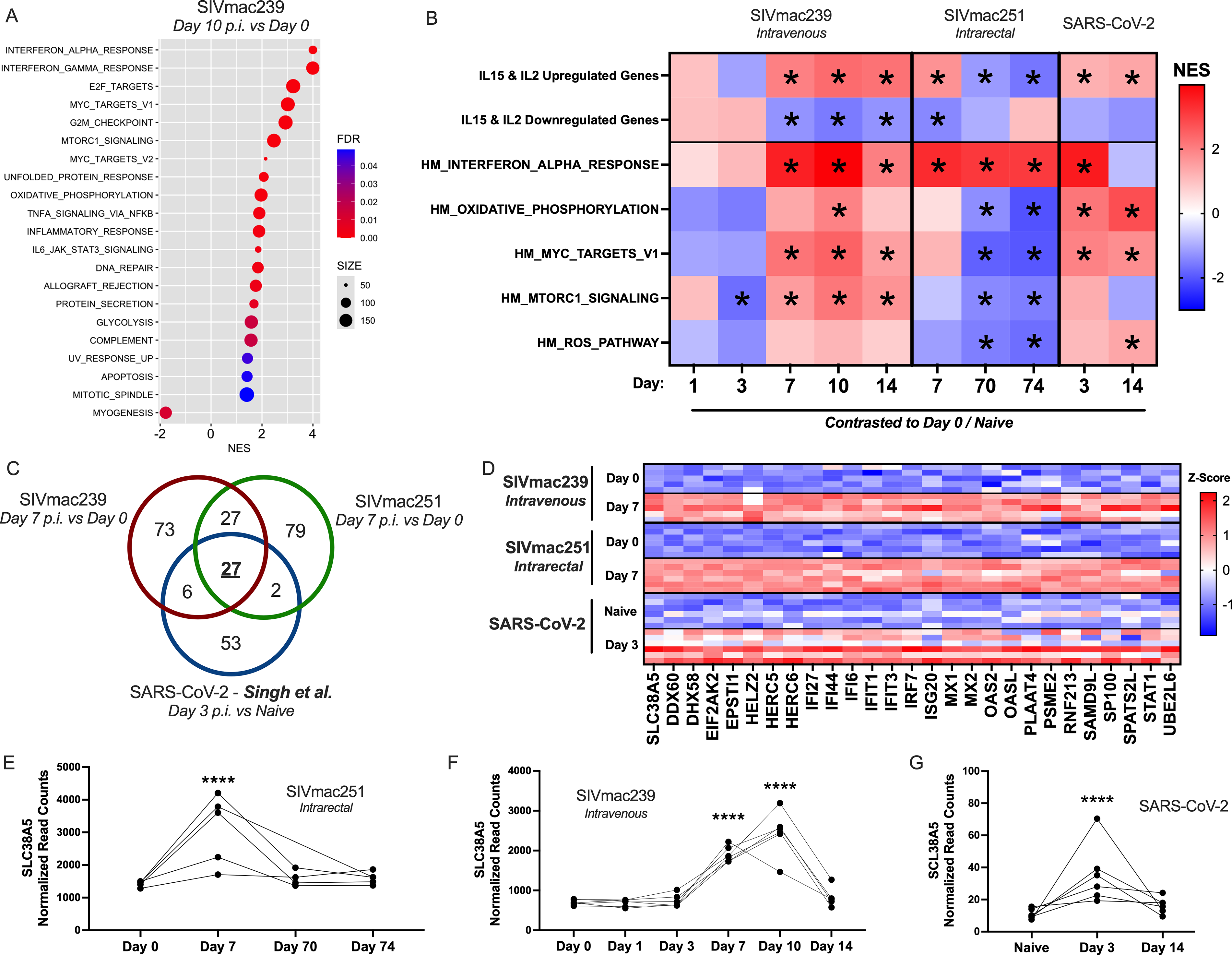
Transcriptomic signature of rhesus macaque (RM) natural killer (NK) cells during acute viral infection. Peripheral NK cells were bulk sorted from six RM intravenously infected with SIVmac239 and from five RM intrarectally infected with SIVmac251. Public scRNA-seq data (GSE190659) were accessed from a study in which six RM were infected with SARS-CoV-2; in the present study, NK cells were subset from total bronchoalveolar lavage immune cells based on immune marker expression and assessed by pseudobulk analysis.
The fact that metabolic pathways central to NK cell effector function were downregulated in chronic infection compared with baseline, while genes activated by type I interferon remained highly expressed, led us to consider alternative gene sets as indicators of NK cell activation and function. We consolidated ImmuneSigDB gene sets, which characterized genes up- and down-regulated by in vitro stimulation of NK cells with IL-15 or IL-2 (GSE7764, GSE22886, and GSE12198) to produce a gene set of those upregulated by IL-15 and/or IL-2 (663 unique genes) and those downregulated by these cytokines (696 unique genes).10–12 GSEA on these summary gene sets revealed that genes upregulated by IL-15 and/or IL-2 were consistently and significantly enriched during the acute phase in both SIV cohorts, and unlike the hallmark interferon response genes, were downregulated during chronic infection (Fig. 1B). This pattern agrees with mTORC1 and MYC pathway expression and likely represents a transcriptional program more indicative of NK cell function than ISG expression alone, a “functional activation signature”. Meanwhile, as expected, genes downregulated by IL-15 and IL-2 incubation were negatively enriched during the acute phase of SIV (Fig. 1B), though this inverse cytokine signature was not upregulated during chronic infection.
To evaluate the broader applicability of these findings related to acute virus infection, we next explored the transcriptional dynamics of RM NK cells in a non-SIV viral context. Using the Gene Expression Omnibus, we accessed 10× single-cell RNA-seq (scRNA-seq) data pertaining to RM bronchoalveolar lavage-derived immune cells before and after SARS-CoV-2 challenge (NCBI GEO, GSE190659). 13 We then employed the R Seurat package to subset NK cells based on immune marker expression as defined by the Human Primary Cell Atlas. 14 GSEA on normalized read counts comparing SARS-CoV-2 pre- and post-infection time points revealed enrichment of interferon signaling at day 3 p.i., which was no longer significant at day 14, by which point the animals had cleared the virus. Conversely, IL-15- and/or IL-2-upregulated genes, coinciding with several metabolic signatures of oxidative phosphorylation and MYC signaling, remained enriched at day 14 p.i., suggesting that these transcriptional states may be maintained for some time after viral clearance (Fig. 1B).
According to GSEA, NK cell transcriptomics at day 3 of SARS-CoV-2 infection aligned mainly with that of NK cells during acute SIV infection. We next evaluated differentially expressed genes (DEGs) that were higher in acute than in pre-infection timepoints for each cohort and identified which viral response genes were shared between cohorts. We identified 27 DEGs common to all three groups, accounting for roughly 30% of genes transcriptionally upregulated in NK cells during SARS-CoV-2 infection (Fig. 1C,D). Of these, all but one were reported to be interferon-inducible in the literature. The remaining gene, not yet associated with interferon signaling, is SLC38A5 (SNAT5), a glutamine transporter with no known function in NK cells. However, the related glutamine importer SLC1A5 is known to drive NK cell function by indirectly facilitating the import of leucine and mTORC1-activating amino acids. 5 Therefore, SLC38A5 may represent a glutamine importer with a similar role in RM. As an alternative conjecture, the presence of SLC38A5 on human dendritic cell lysosomal membranes suggests a role in lysosomal acidification in NK cells. 15 Either function would explain the upregulation of SLC38A5 during acute infection (Fig. 1D,E) and highlight the importance of metabolic transporters during antiviral activity.
By investigating NK cell expression patterns across various infection models, we have constructed a more comprehensive map of RM NK cell antiviral programming, characterized by cytokine signaling and altered metabolism. We have traced NK cell transcriptional changes from acute to chronic lentiviral infection. This timeline coincides with a critical shift in immune cell function and identifies diminished gene expression in oxidative phosphorylation, mTORC1/c-MYC pathways, and IL-2/15 signaling as infection progresses. It is likely that these effects of an evolving lentiviral infection on peripheral NK cells not only indicate but also inform the dysregulation of NK cells following the acute phase, thereby contributing to loss of viral control. Future NK cell-oriented HIV therapeutics must fine-tune activating signals to avoid exhaustion, a difficult task given the extreme complexity of the in vivo cytokine milieu and intracellular apparatus; however, the specific metabolic and cytokine pathways emphasized in this study represent useful indicators of NK cell function and dysfunction and may serve as targets for future therapeutic strategies.
Data Availability
The datasets generated for this study using the SIVmac251 cohort can be found in the NCBI GEO, accession GSE329639. The datasets generated for the SIVmac239 cohort can be found under accession GSE329638. In addition, data regarding the naïve timepoints for the SIVmac39 cohort have previously been made available under accession GSE148290. 7
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by National Institute of Health research grants R01AI161010, UM1AI164570, and R21AI184422 (to R.K.R.).