
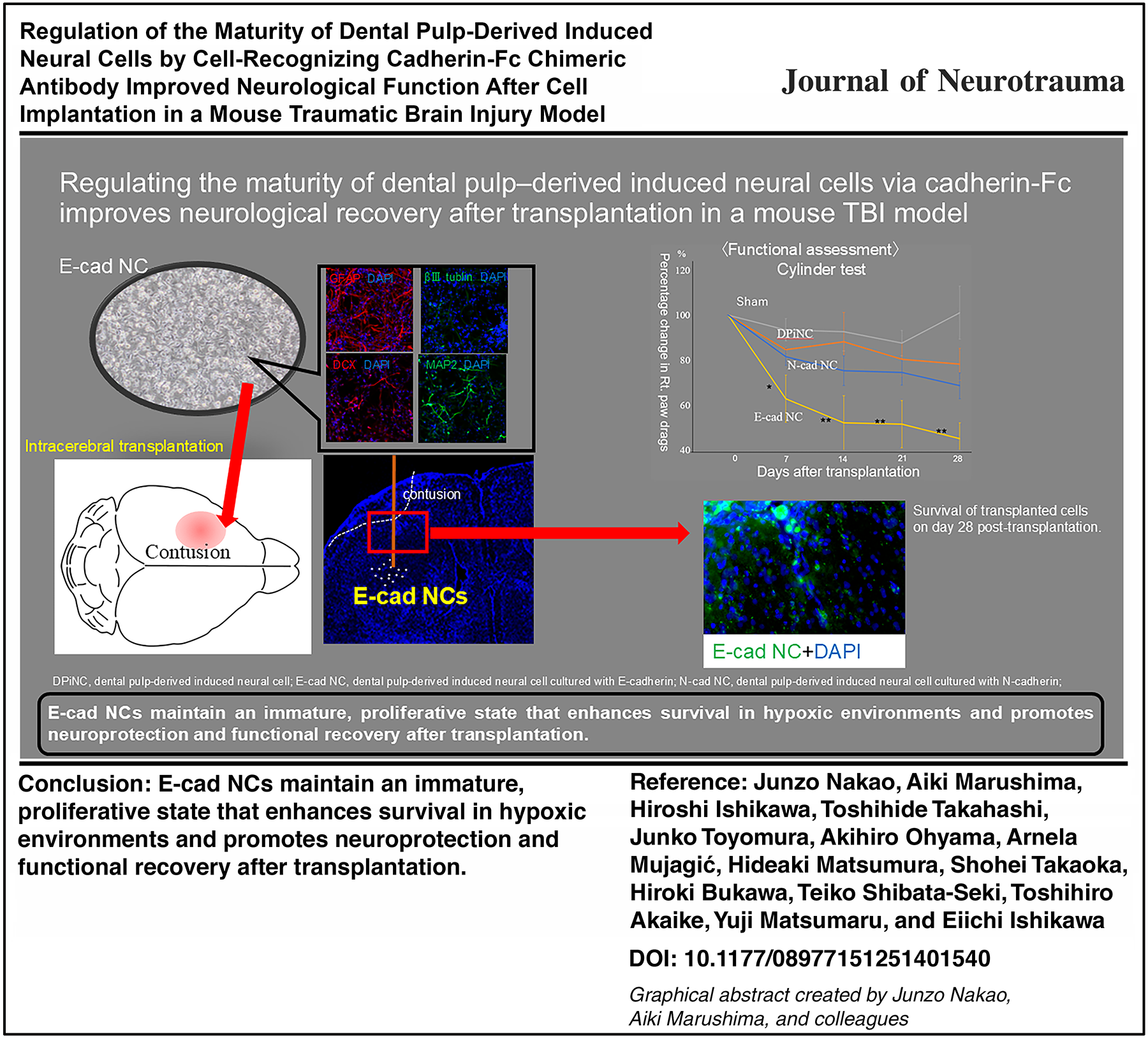
Abstract
Biomaterial-based neural cell transplantation has displayed promise in regenerative medicine. E-cadherin and N-cadherin are cell–cell adhesion proteins with important roles in neuronal maturity. Therefore, cell culture on Petri dishes coated with a fusion protein combining the human E-cadherin or N-cadherin extracellular domain with an immunoglobulin G Fc region chimeric antibody (E-cad-Fc and N-cad-Fc) can regulate cell maturity through the process of neuronal differentiation in stem cells. This study explored the efficacy of dental pulp-derived induced neural cells (DPiNCs), which were cultured in dishes coated with cell-recognizing E-cadherin-Fc chimeric antibody (E-cad NCs) or N-cadherin-Fc chimeric antibody (N-cad NCs), in promoting neural regeneration in a traumatic brain injury (TBI) model. In vitro, DPiNCs featured axon-like projections with positive GFAP expression, weak positivity for DCX and βIII-tubulin, and positivity for MAP2 and vascular endothelial growth factor, indicating that the cell population included a mixture of mature and immature neurons. E-cad NCs exhibited staining for DCX and βIII-tubulin but reduced MAP2 staining, indicating immaturity. N-cad NCs displayed weaker DCX and βIII-tubulin staining but increased MAP2 staining, indicating maturity. In an oxygen–glucose deprivation/reoxygenation model, which simulates secondary brain injury, E-cad NC cells maintained higher βIII-tubulin and DCX expression and extended axons, denoting resistance to hypoxic stress. N-cad NCs displayed stronger MAP2 staining and axon extension, indicating resilience and maturity. In vivo, TBI model mice were transplanted with each cell type. Regarding motor function, the wire hang test revealed a significant improvement in the E-cad NC group with the other cell groups and sham group on day 28 post-transplantation. Additionally, in the cylinder test (right paw drags), a significant improvement was noted in the E-cad NC group compared with the DPiNC and sham groups from day 14 to day 28 post-transplantation. The E-cad NC group exhibited the highest rate of improvement on the Revised Neurobehavioral Severity Scale. Dual immunostaining on day 28 confirmed that the number of surviving transplanted cells was higher in the N-cad NC group than in the DPiNC group. Our study demonstrated the potential of cadherin-modified DPiNCs in TBI treatment, suggesting that controlling cell maturity through cadherin expression can significantly improve outcomes by promoting cell survival, integration, and neural regeneration. This study offers promising insights into regenerative medicine for acute-phase TBI, but further research is needed to clarify the long-term efficacy and the mechanisms underlying cell integration and neural network reconstruction.
Graphical Abstract
Keywords
Introduction
Traumatic brain injury (TBI) is a major cause of mortality and functional impairment.1–4 TBI results in brain dysfunction attributable to damage to neural structures, including mechanical destruction of the brain parenchyma, cranial nerves, and cerebral blood vessels. 3 Primary brain injury occurs from direct damage to neural structures caused by external forces. 5 Conversely, secondary brain injury follows primary brain injury, and it is the consequence of a series of pathophysiological changes, including reduced blood flow, cerebral edema, hypoxia, inflammation, and cell death. These complex biochemical cascades further exacerbate brain damage.6–8
Regenerative medicine has emerged as a novel approach for TBI treatment. Specifically, neural cell transplantation into the brain has displayed promise in treating primary brain injuries.9–12 Nevertheless, the low survival and engraftment rates of transplanted neural cells represent significant challenges. 13 Additionally, it is critical that the transplanted cells can appropriately differentiate within the niche of the disease site. This ability is attributed to the maturity of the transplanted neural cells, resulting in a scarcity of precursor cells and adverse conditions during the acute phase of TBI. Transplanting mature neural cells is believed to result in low engraftment and proliferation rates. 14 Conversely, transplanting proliferating neural precursor cells into spinal cord injury model mice confirmed the survival of these cells and their differentiation into neurons, along with improvements in motor disorders. Furthermore, mature neural cells have high demands for nutrients and oxygen, and they do not undergo cell division. Therefore, immature nerve stem cells might be effective in transplantation given their ability to divide and appropriately differentiate within the disease niche and their lower oxygen and nutrient requirements. 15
In neural cell transplantation for TBI, it is critical to maintain stable proliferation while keeping neural cells in an immature state. However, stably maintaining an immature state while achieving a large population of neural cells is challenging. 16 To address this issue, we explored the potential of cadherin molecules, especially E-cadherin (E-cad) and N-cadherin (N-cad). E-cad plays significant roles in cell adhesion, tissue preservation, the maintenance of pluripotency, and cell maturity and differentiation.17–21 N-cad, known for its crucial roles in neural cell development, projection growth, synapse formation, and neural circuit construction, enhances cell adhesion and improves the engraftment of transplanted neural cells. 22 Leveraging the effects of E-cad and N-cad in cell culture for transplantation provides both control over cell maturity and enhances cell engraftment and post-transplantation survival.
Techniques have been developed that apply cadherins to cell culture. E-cad-Fc is a chimeric antibody in which E-cad is fused with the Fc fragment. Fc fusion proteins typically include the antibody Fc region (located at the lower part of the IgG molecule, playing a crucial role in receptor binding and the regulation of immune responses) at the N-terminus of the protein, and they have garnered attention as cell-recognition antibodies in the field of biomaterials.23–26 Neural stem cells can be cultured on plates coated with E-cad-Fc.16,24 A similar chimeric protein, N-cad-Fc, has also been developed, and it has been found to promote the differentiation of neural precursor cells in vitro.27–30 Additionally, in experiments aimed at developing treatments for neonatal brain injuries in mice, gelatin sponge scaffolds coated with N-cad-Fc promoted nerve regeneration and functional recovery, suggesting the potential of N-cad to significantly aid in cell migration in regenerative medicine. 31 Cells cultured on plates coated with these cadherin chimeric proteins adhere in the presence of Ca2+, facilitating attachment and proliferation, and detachment is achieved using chelating agents. These biomaterials support the maintenance of an undifferentiated state, which is critical for effective transplantation.
Dental pulp stem cells (DPSCs) are pluripotent cells capable of multilineage differentiation.32,33 Because DPSCs are derived from the neural crest, they are prone to differentiate into neural cells. DPSCs, with their ability to differentiate into both neural and glial cells, hold promise for repairing and regenerating neurons in central nervous system disorders.34,35 Additionally, DPSCs exhibit immunomodulatory and anti-inflammatory properties, enhancing their therapeutic potential.12,36
Therefore, we employed dental pulp-derived induced neural cells (DPiNCs), a mixture of neural and glial cells derived from DPSCs, with demonstrated efficacy in neurological recovery and cell engraftment in TBI and acute stroke models,34,35 and assessed their efficacy in the treatment of TBI using a mouse model. We hypothesized that manipulating the maturity of transplanted cells would enhance their survival and integration, thereby facilitating neural regeneration. Our study investigated the influence of cadherins, which are critical molecules in morphogenetic processes such as synapse and vasculature formation,5–7 in an effort to develop a regenerative medicine approach using DPiNCs regulated by E-cad-Fc and N-cad-Fc to control cell maturation.
Materials and Methods
Induction of DPSCs
Human dental pulp was harvested from adult third molars designated for extraction without caries, with the approval of the Ethics Committee of the University of Tsukuba Hospital (approval number: H30-181), and processed under sterile conditions. After extraction, the pulp was minced and cultured in growth medium comprising DMEM/F12, 10% FBS, 10 μM MEM nonessential amino acids (Nacarai Tesque, Inc., Kyoto, Japan), 2 mM GlutaMAX, 50 U/mL penicillin, 50 μg/mL streptomycin (Fujifilm Wako Pure Chemical Corporation, Osaka, Japan), and 0.25 μg/mL fungizone (Hyclone Laboratories, Logan, UT) at 37°C in a 4.7% CO2 atmosphere. Following initial adherence, the medium was gently supplemented. Upon reaching 80% confluency, cells were passaged using 0.1% trypsin–0.02% EDTA/phosphate-buffered saline (PBS) (−) solution. DPSCs were isolated from passages 1 to 3, expanded using the filter paper cloning method, and further cultured for characterization.
Neural cell induction from DPSCs
DPSCs were cultured in neural induction medium containing DMEM/F12, 5% FBS, 15 nM all-trans-retinoic acid, 20 nM each of progesterone, estradiol, NGF-1, 10 ng/mL thyroxine, 10 nM dexamethasone, 50 μM ascorbic acid, and 20 ng/mL IGF-1. After 5–6 weeks, DPiNCs with neural projections were isolated using the filter paper method and cultured in N-GRO maintenance medium (DV Biologics, Yorba Linda, CA) on Matrigel-coated (Becton Dickinson and Company, Franklin Lakes, NJ) dishes.
Cadherin-Fc coating
DPiNCs were cultured on dishes coated with E-cad-Fc or N-cad-Fc to study cell adhesion and differentiation. The coating process involved diluting E-cad-Fc and N-cad-Fc (both from Somar Corporation, Tokyo, Japan) to 10 μg/mL in PBS (−), and the respective solutions were applied to non-treated dishes (Iwaki, Tokyo, Japan), followed by incubation for 2 h. After washing with PBS, DPiNCs were seeded in neural maintenance medium, which was refreshed twice a week depending on the differentiation status. Cells cultured on E-cad-Fc–coated dishes were named E-cad NCs, and those on N-cad-Fc-coated dishes were named N-cad NCs.
Immunofluorescence staining
DPiNCs, cultured under specified conditions, were fixed with 4% paraformaldehyde (PFA) for 15 min, blocked with Blocking One Histo solution (Nacalai Tesque), and incubated with primary antibodies overnight at 4°C (Table 1). Following incubation with secondary antibodies, cells were mounted with 4′,6-diamidino-2-phenylindole (DAPI)-containing mounting medium (Immunoselect Antifading Mounting Medium DAPI, Dianova GmbH, Hamburg, Germany) for examination under a fluorescence microscope (Leica Microsystems, Wetzlar, Germany).
List of Primary Antibodies
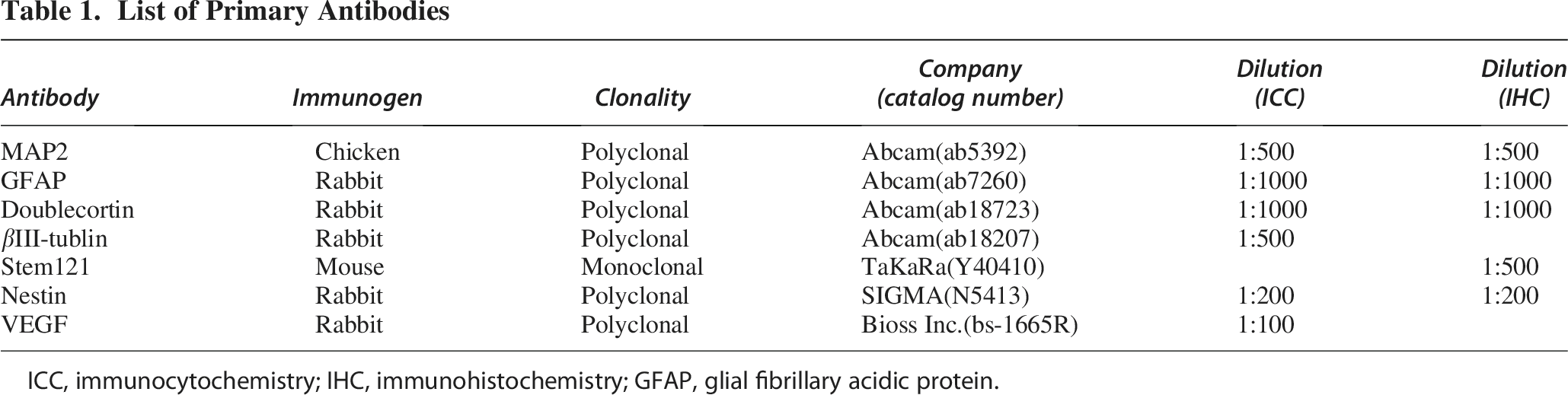
ICC, immunocytochemistry; IHC, immunohistochemistry; GFAP, glial fibrillary acidic protein.
Immunocytochemistry and quantification of marker-positive cells
To evaluate the differentiation characteristics of DPiNC, E-cad NC, and N-cad NC, immunofluorescence images were analyzed using ImageJ software (National Institutes of Health, Bethesda, MD). Moreover, for each marker (glial fibrillary acidic protein [GFAP], β3-tubulin, MAP2, DCX), three random microscopic fields were selected per group, and double-positive cells (marker+/DAPI+) were counted to calculate the percentage of marker-positive cells.
Neurotrophic factor measurement
Neurotrophic and nutritional factors in the culture medium were quantified using ELISA kits (vascular endothelial growth factor [VEGF]: DVE00, R&D Systems, Minneapolis, MN; nerve growth factor β [NGFB]: ab99986, Abcam, Cambridge, UK; brain-derived neurotrophic factor [BDNF]: BEK-2211-1P, Biosensis, Thebarton, Australia; epidermal growth factor [EGF]: ELISA kit PGI KE00138, Proteintech, Rosemont, IL; glial cell line-derived neurotrophic factor [GDNF]: DGD00, R&D Systems) after 48 h of culture.
Gene expression analysis
Microarray analysis was performed using the Clariom S Human Assay (Thermo Fisher Scientific, Waltham, MA) to assess gene expression in DPiNCs, E-cad NCs, and N-cad NCs. RNA purity and integrity were verified using the Eppendorf BioSpectrometer basic (Eppendorf, Hamburg, Germany) and 1% agarose gel, respectively, before analysis by Thermo Fisher Scientific. Data normalization and expression analysis were conducted using Transcriptome Analysis Console version 4.0 (Thermo Fisher Scientific).
Oxygen–glucose deprivation/reoxygenation model
The oxygen
Axonal length measurement
Axonal length was evaluated to determine cell maturity by phase-contrast microscopy at ×200 magnification for image capture.
Generation of a TBI mouse model by cold injury
The Institutional Animal Care and Use Committee of the University of Tsukuba approved all experimental mouse studies (approval number: 22-111), ensuring adherence to the “Guidelines for the Management and Use of Laboratory Animals.” C.B-17/Icr scid/scid Jcl mice (male, 7–8 weeks old, weighing 22–26 g, CLEA, Tokyo, Japan) were maintained with free access to food and water. To create the TBI model, the animals were subjected to cold injury of the primary motor cortex of the right upper limb. Mice were anesthetized via an intraperitoneal injection of ketamine (Ketalar, 70 mg/kg, Daiichi Sankyo Pharmaceutical, Tokyo, Japan) and xylazine (Sedactal, 14 mg/kg, Bayer, Leverkusen, Germany). After securing the mouse using a head holder (Narishige, Tokyo, Japan), a longitudinal skin incision was created to expose the skull using a 3-mm copper rod chilled in liquid nitrogen applied perpendicularly for 30 sec. The incision was made 2.5 mm caudal from bregma and 2.5 mm left of the midline (Fig. 1).
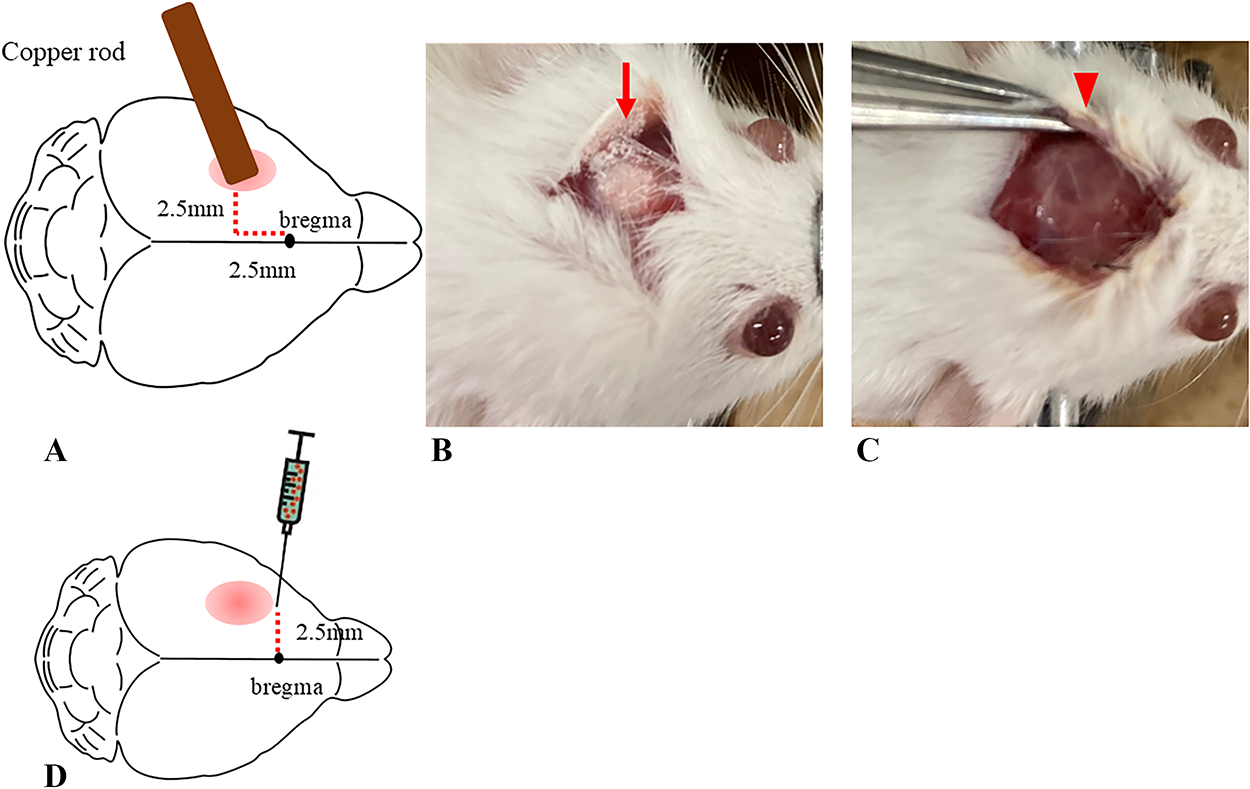
Creation of TBI model mice and transplantation of cells.
Preparation and collection of transplant cells
DPiNCs were cultivated on dishes coated with Matrigel, E-cad-Fc, and N-cad-Fc until reaching 80–100% confluence. This typically required approximately 5 days of culture. To align with the transplantation schedule, TBI induction timing was carefully coordinated to ensure that the cells would reach the appropriate confluence level exactly 2 days later. Cells on Matrigel-coated dishes were detached using trypsin, whereas 0.5 mM EDTA (TaKaRa, Shiga, Japan) was employed for E-cad-Fc– and N-cad-Fc–coated dishes. Following centrifugation at ≈ 42,800 × g for 5 min, cells were resuspended in Hanks’ solution at a concentration of 1 × 105 cells/2 μL for transplantation. In a preliminary experiment, E-cad NCs were transplanted to TBI model mice at varying densities (1 × 104, 5 × 104, and 1 × 105 cells) 2 days after TBI, and a control sham group was included for comparison. By day 28 post-transplantation, substantial improvements in motor and sensory functions were observed across all transplantation groups compared with the findings in the sham group, with the most pronounced improvement observed for 1 × 105 cells/2 μL Hanks’ solution.
Direct intracerebral transplantation of neural cells into TBI model mice
After anesthetizing the mice via an intraperitoneal injection of a ketamine–xylazine mixture, their heads were fixed using the head holder. The skin incision made at the time of TBI creation was reopened, and a 23-gauge needle was used to manually perforate the skull at a position 2.5 mm to the left of bregma around the TBI site (Fig. 1). A syringe (Hamilton micro-syringe 701RN, Hamilton Company, Reno, NV) was then inserted to a depth of 2.0 mm from the brain surface through the perforation and left in place for 1 min. Afterward, the cells were slowly injected into the brain over a period of 8 min at a rate of 0.25 μL/min. The needle was left in place for 5 min after the injection and then slowly withdrawn. The incision site was sutured to complete the procedure.
The following four groups were included: (1) sham group (only craniotomy without cell injection; n = 6); (2) DPiNC group (transplanted with DPiNCs; n = 6); (3) E-cad NC group (transplanted with E-cad NCs; n = 6); and (4) N-cad NC group (transplanted with N-cad NCs; n = 6). In the E-cad NC group, one mouse died the day after TBI. Cell transplantation was completed, and the brain was harvested, but the cause of death could not be confirmed. Consequently, behavioral evaluations were conducted with n = 5 in this group.
Cell transplantation was performed 2 days after TBI induction. The timing was selected to reflect the acute phase of injury and match the completion of cell culture at 80–100% confluence.
Assessment of transplanted cells
At 28 days post-transplantation, mouse brains were extracted to evaluate engraftment and differentiation. Following deep anesthesia, the brains were perfused with Hanks’ solution followed by 4% PFA, and subsequently cryopreserved in sucrose solutions (10%, 20%, and 30% in sequence) before embedding in optimal cutting temperature compound (Sakura Finetek, Tokyo, Japan). Coronal sections (10 μm thick) were analyzed via immunostaining and fluorescence microscopy.
Symptom assessment in mice
Functional recovery was assessed using the wire hang test, cylinder test, Revised Neurobehavioral Severity Scale (NSS-R), and marble-burying test. The tests were conducted before TBI, 2 days post-TBI (pretransplantation), and weekly thereafter, with the results compared among the sham (n = 6), DPiNC (n = 6), E-cad NC (n = 5), and N-cad NC groups (n = 6).
Wire hang test
Mice were placed on a wire mesh (10 × 10 cm2) and observed for 30 sec. Their performance was scored as follows: falling, 0 points; hanging by both forelimbs, 1 point; hanging and attempting to climb, 2 points; using both forelimbs and one hindlimb, 3 points; using all limbs and the tail, 4 points; and escaping the mesh, 5 points. The average score from four attempts was recorded.
Cylinder test
Mice were placed in a transparent cylinder (10 cm in diameter) to observe forelimb use, which is particularly relevant for mice with left cerebrum contusions affecting right forelimb function. The test measured the stability of the right forelimb against the cylinder wall. Stable contact was considered normal, and partial contact or slipping was recorded as paw drags. The test calculated the percentage of right forelimb use in 20 consecutive bilateral uses and the percentage of paw drags in 20 consecutive right forelimb uses.
Revised Neurobehavioral Severity Scale
This scoring method assessed balance, motor coordination, and sensorimotor reflexes across 10 items, each scored from 0 (optimal) to 2 (impaired), with a total possible score of 20. Lower scores indicate better function. The test components included balance on a beam, landing reflex, limb extension reflex upon tail lift, resistance to backward drag, righting reflex from supine, ear and eye reflexes to touch, reaction to sound, tail pinch response, and paw flexion reflex upon pinch.
Marble-burying test
This test evaluated anxiety-like behavior by counting how many of 20 glass marbles, placed atop a 5-cm layer of wood shavings in a standard cage, were buried by the mouse over 30 min. A successfully burying attempt was indicated by burying more than two-thirds of the marble’s volume, with a higher count suggesting increased anxiety behavior.
Statistical analysis
Data were presented as the mean ± standard error and analyzed using IBM SPSS Statistics 28.0 (IBM, Armonk, NY). Student’s t-test and repeated-measures analysis of variance were utilized for statistical comparisons, with significance set at p < 0.05.
Results
Induction of DPiNCs from DPSCs
DPiNCs were evaluated for morphological and immunocytochemical properties, as reported previously.34,36 Whereas DPSCs were spindle-shaped, DPiNCs exhibited axon-like projections. Immunostaining identified positive expression for GFAP, weak positivity for DCX and βIII-tubulin, and positivity for MAP2. Notably, VEGF expression was robustly positive in DPiNCs, indicating a mixture of mature and immature neuronal traits along with astrocytic characteristics (Fig. 2).
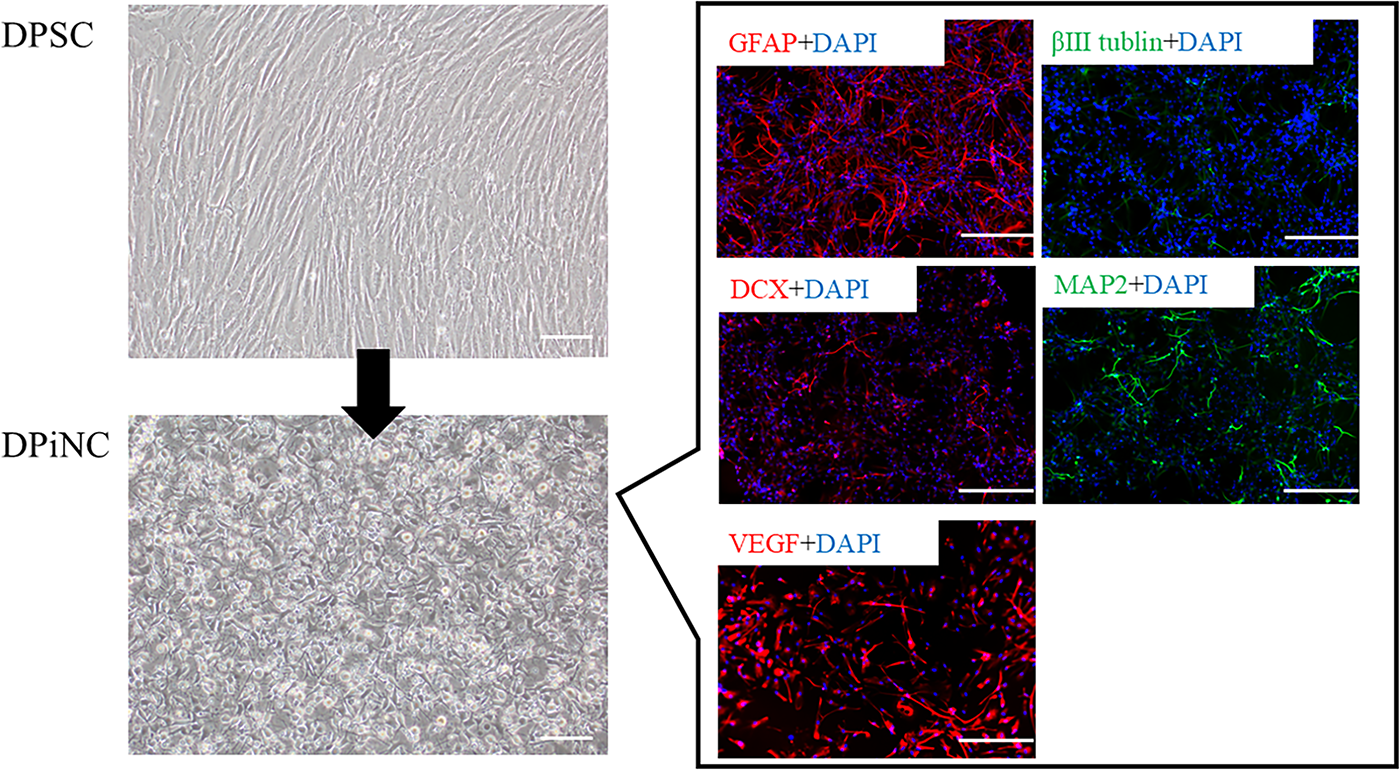
Generation of DPiNCs from DPSCs. Induction from DPSCs (top left) to DPiNCs (bottom left). To the right, the immunostaining of DPiNCs is displayed. Immunostaining was performed on DPiNCs at 80% confluence. The scale bar represents 100 µm. DPiNC, dental pulp-derived induced neural cell; DPSC, dental pulp stem cell; GFAP, glial fibrillary acidic protein; VEGF, vascular endothelial growth factor.
Differential properties of DPiNCs, E-cad NCs, and N-cad NCs
E-cad NCs displayed short axon-like structures and rounded cytoplasm, contrasting the features of DPiNCs. N-cad NCs exhibited thicker cytoplasm and extended axon-like structures. Immunostaining revealed enhanced staining for DCX and βIII-tubulin but reduced staining for MAP2 in E-cad NCs, indicating a more immature phenotype compared with DPiNCs. Conversely, N-cad NCs exhibited lesser staining for DCX and βIII-tubulin but increased staining for MAP2, suggesting a more mature state (Fig. 3).
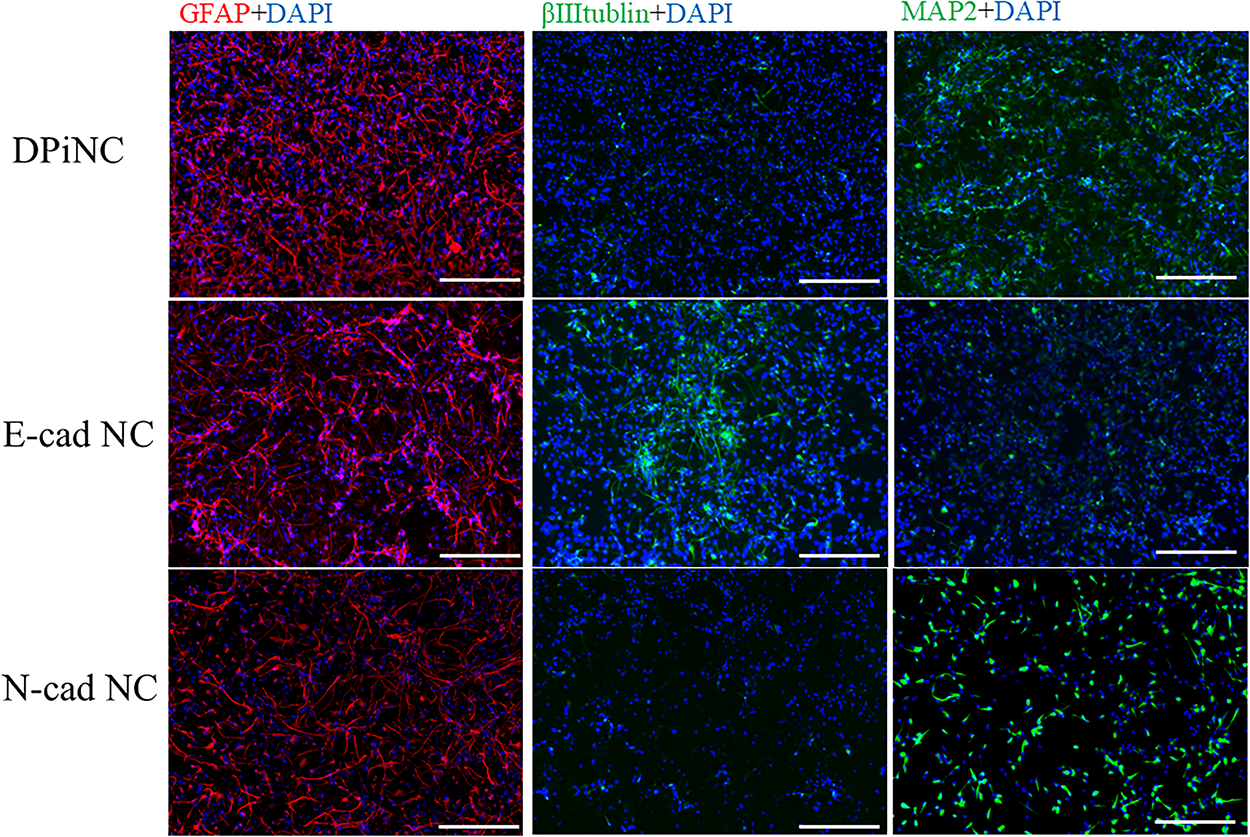
Immunostaining of each cell type. Costaining with various antibodies and DAPI was performed at 80% confluence. The scale bar represents 100 µm. DPiNC, dental pulp-derived induced neural cell group; E-cad NC, dental pulp-derived induced neural cell cultured with E-cadherin group; GFAP, glial fibrillary acidic protein; N-cad NC, dental pulp-derived induced neural cell cultured with N-cadherin group.
Neurotrophic and nutritional factor analysis
Comparative analysis demonstrated that VEGF, NGFB, and EGF levels were significantly increased in E-cad NCs, supporting angiogenesis, survival, and regeneration. N-cad NCs exhibited elevated levels of BDNF and GDNF, which are crucial for neuronal survival and maturation, highlighting distinct regenerative profiles between E-cad NCs and N-cad NCs (Fig. 4).

Comparison of neurotrophic and nutrient factor levels among the different cell types. The secretion of neurotrophic and nutrient factors was compared among the three cell types. *p < 0.01. BDNF, brain-derived neurotrophic factor; DPiNC, dental pulp-derived induced neural cell group; E-cad NC, dental pulp-derived induced neural cell cultured with E-cadherin group; EGF, epidermal growth factor; GDNF, glial cell line-derived neurotrophic factor; N-cad NC, dental pulp-derived induced neural cell cultured with N-cadherin group; NGFB, nerve growth factor β; VEGF, vascular endothelial growth factor.
Gene expression analysis
Microarray analysis highlighted distinct gene expression profiles for E-cad NCs and N-cad NCs compared with DPiNCs. Notable upregulation of genes related to cell differentiation, neuronal migration, and neuroprotection was observed in both E-cad NCs and N-cad NCs (Fig. 5). Both cell types strongly expressed proteins associated with neural protection and neural/axonal regeneration, especially chitinase-3-like protein 1 (CHI3L1) and αB-crystallin (CRYAB), although the types of genes varied between the two cell types.
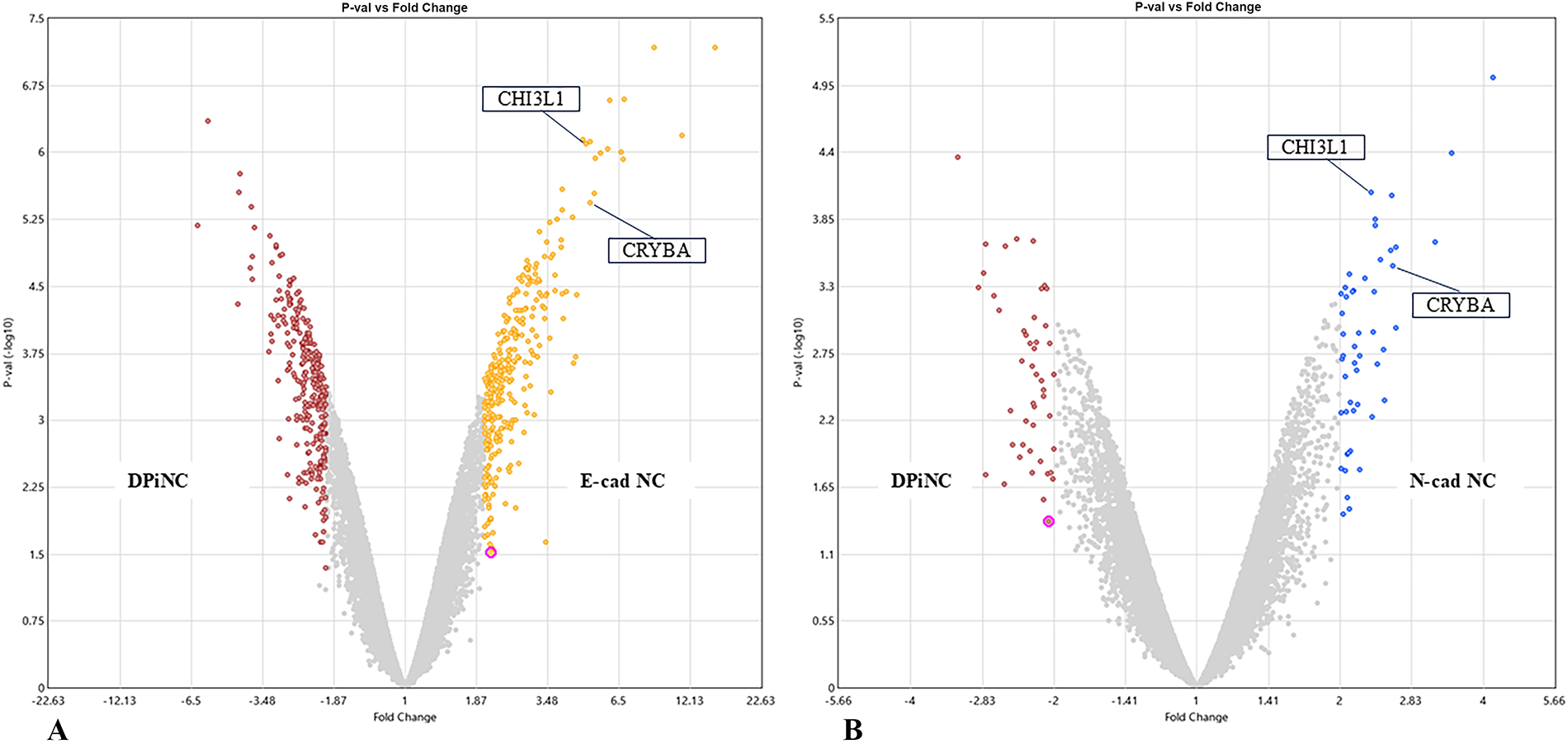
Comprehensive gene analysis of each cell type.
Response to OGD/R
E-cad NCs displayed higher βIII-tubulin and DCX levels than DPiNCs, indicating an immature but proliferative state post-OGD/R. By contrast, MAP2 staining was weaker (Fig. 6). E-cad NCs displayed extended axons post-OGD/R, unlike DPiNCs, which exhibited reduced axon length, demonstrating the superior resistance of E-cad NCs to hypoxic stress (Fig. 7).
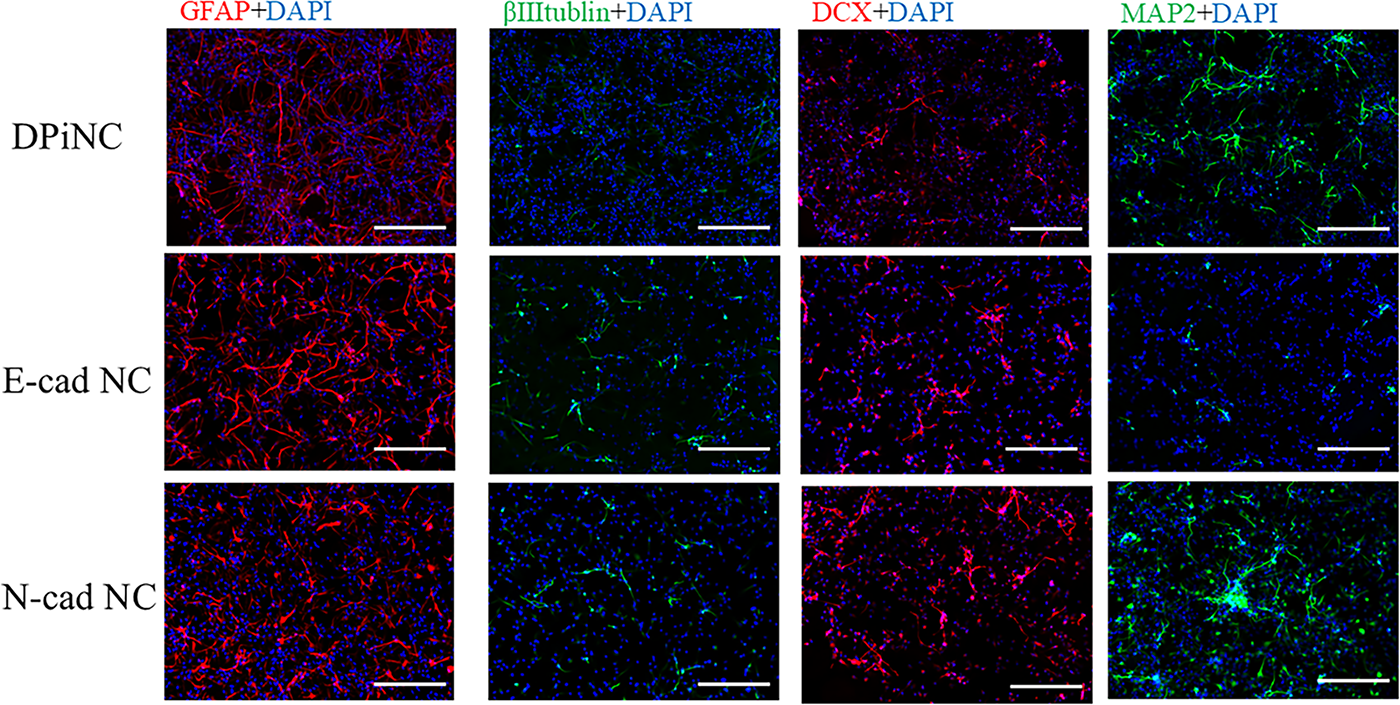
Immunostaining after OGD/R. Immunostaining was performed on each cell type following OGD/R. The scale bar represents 100 µm. DPiNC, dental pulp-derived induced neural cell group; E-cad NC, dental pulp-derived induced neural cell cultured with E-cadherin group; N-cad NC, dental pulp-derived induced neural cell cultured with N-cadherin group; OGD/R, oxygen–glucose deprivation/reoxygenation; sham, sham-operated group.
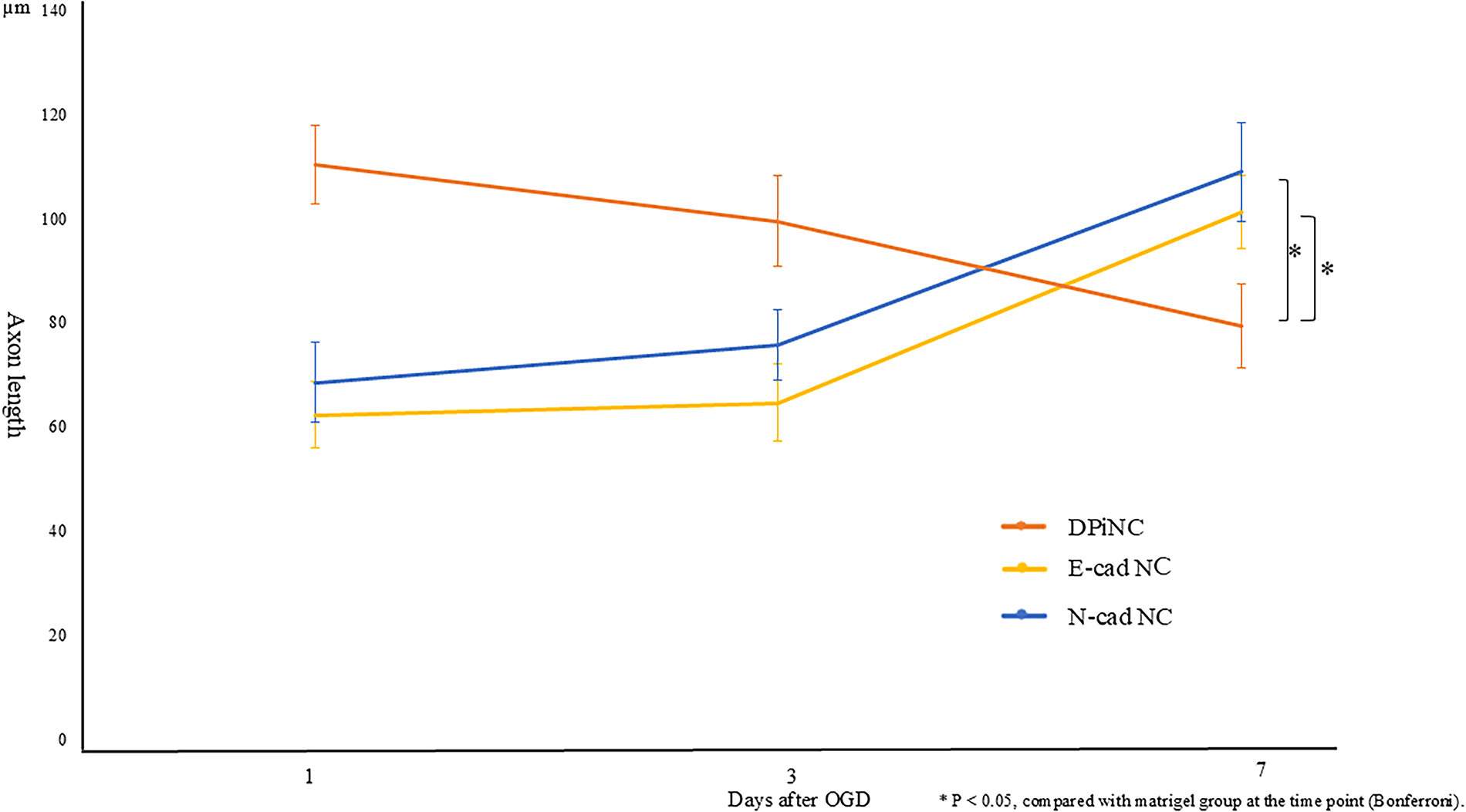
Progression of axon length after OGD/R. Axon length was measured to assess cellular damage in each cell type after OGD/R. Twenty cells were randomly selected from those photographed with a phase-contrast microscope, and the average axon length was calculated. *p < 0.05 compared with the Matrigel group at the same time point (Bonferroni). DPiNC, dental pulp-derived induced neural cell group; E-cad NC, dental pulp-derived induced neural cell cultured with E-cadherin group; N-cad NC, dental pulp-derived induced neural cell cultured with N-cadherin group; OGD/R, oxygen–glucose deprivation/reoxygenation; sham, sham-operated group.
N-cad NCs featured stronger MAP2 staining than DPiNCs under both normal and OGD/R conditions, indicating a mature and proliferative state (Fig. 6). Axon length analysis revealed that axons grew in N-cad NCs post-hypoxia, indicating resilience and growth potential under stress (Fig. 7).
Quantitative evaluation of marker expression in cultured cells
In normal culture, N-cad NCs exhibited significantly higher GFAP positivity compared with DPiNCs (p = 0.048), indicating enhanced glial characteristics. E-cad NCs showed significantly higher βIII-tubulin expression than DPiNCs (p = 0.014) and N-cad NCs (p = 0.030), suggesting a more pronounced immature neuronal phenotype. Although MAP2 positivity showed no statistically significant differences among groups, N-cad NCs tended to express higher levels (Supplementary Fig. S1).
Moreover, under OGD/R conditions, no significant differences were observed in GFAP expression. However, βIII-tubulin expression remained significantly higher in E-cad NCs compared with DPiNCs (p = 0.017), suggesting resilience in maintaining immature neuronal characteristics under hypoxia. Furthermore, DCX expression was significantly higher in E-cad NCs (p = 0.013) and N-cad NCs (p = 0.003) compared with DPiNCs. MAP2 expression was markedly elevated in N-cad NCs, with significant differences observed against DPiNCs (p = 0.001) and E-cad NCs (p = 0.014), indicating a shift toward neuronal maturation under stress (Supplementary Fig. S2).
Determination of nervous system cells for transplantation into TBI model mice
In the TBI model mice, E-cad NCs were transplanted around the contusion site at varying densities (1 × 104, 5 × 104, and 1 × 105 cells) 2 days post-TBI, and a control sham group was included for comparison. By day 28 post-transplantation, significant improvements in both motor and sensory functions were observed across all transplantation groups compared with the findings in the sham group, with the most pronounced improvement observed for 1 × 105 cells. As such, this cell count was used in subsequent experiments.
Improved motor function and sensory responses in the E-Cad NC transplantation group
Wire hang test
Whereas no improvement was observed in the sham group after TBI, all transplantation groups displayed a tendency toward improvement. On days 14 and 21 post-transplantation, significant improvements compared with the sham group were observed for both E-cad NCs and N-cad NCs. However, by the final assessment on day 28 post-transplantation, a significant improvement versus the sham group was only maintained in the E-cad NC group. Compared with the findings on day 0, the wire hang test score had increased by 1.93 ± 0.44 points on day 28 in the E-cad NC group. Meanwhile, increases of 1.15 ± 0.46 and 1.25 ± 0.41 points were recorded in the DPiNC and N-cad NC groups, respectively. Conversely, the score decreased by 0.12 ± 0.55 points in the sham group (Fig. 8A).
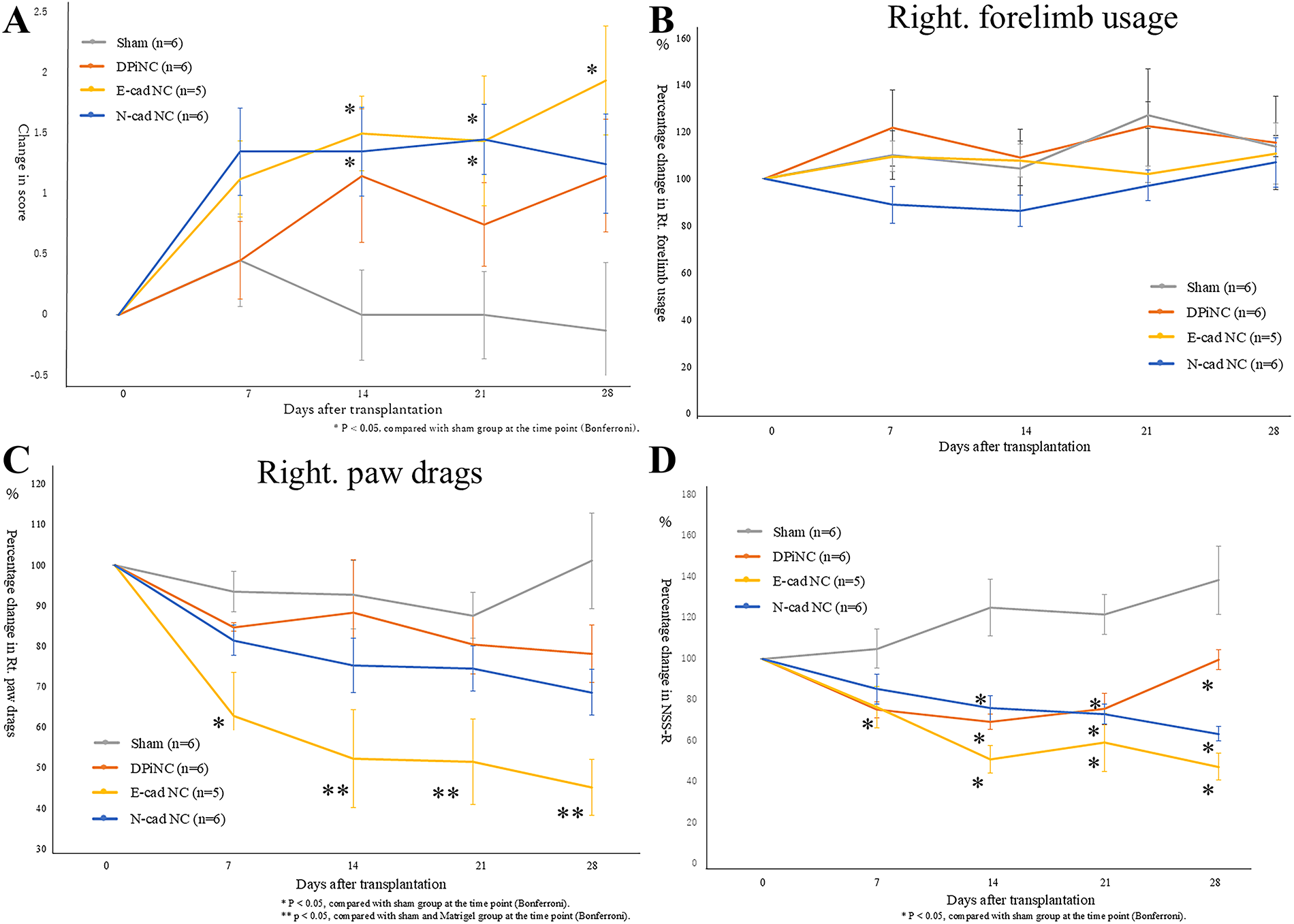
Symptom assessments.
Cylinder test
In the assessment of right forelimb usage, no significant changes were observed in any of the transplantation groups compared with the sham group after transplantation. However, concerning right paw drags, which are considered to more strongly reflect the degree of paralysis, mice in both the E-cad NC and N-cad NC transplantation groups displayed improvements in symptoms over time. Furthermore, starting from day 7 post-transplantation, a significant improvement was observed in the E-cad NC group compared with both the sham and DPiNC groups. Compared with the findings on day 0, the score decreased by 45.15 ± 5.69% in the E-cad NC group, 78.16 ± 7.06% in the DPiNC group, and 68.60 ± 6.96% in the N-cad NC group. In contrast, the score in the sham group increased by 1.06 ± 11.76 points (Fig. 8B,C).
Revised Neurobehavioral Severity Scale
Compared with the results in the sham group, the DPiNC group displayed significant improvement from day 7 post-transplantation, whereas significant improvement was noted from day 14 in the E-cad NC and N-cad NC groups. Among the transplantation groups, no significant differences were observed, but the E-cad NC group exhibited the highest rate of improvement on day 28 (47.94%, Fig. 8D).
Marble-burying test
After transplantation, all groups displayed a trend of worsening symptoms. However, no significant differences were observed among the groups up to day 28 post-transplantation.
Assessment of transplanted cells
Dual immunostaining on day 28 post-transplantation confirmed the survival and differentiation of transplanted cells within the brain, with the cells extending from the transplantation site to the contusion area. The transplanted cells were positive for STEM121, indicative of their human origin, and they differentiated into mature neurons, immature neurons, and astrocytes, as evidenced by positivity for nestin, DCX, MAP2, and GFAP (Fig. 9).
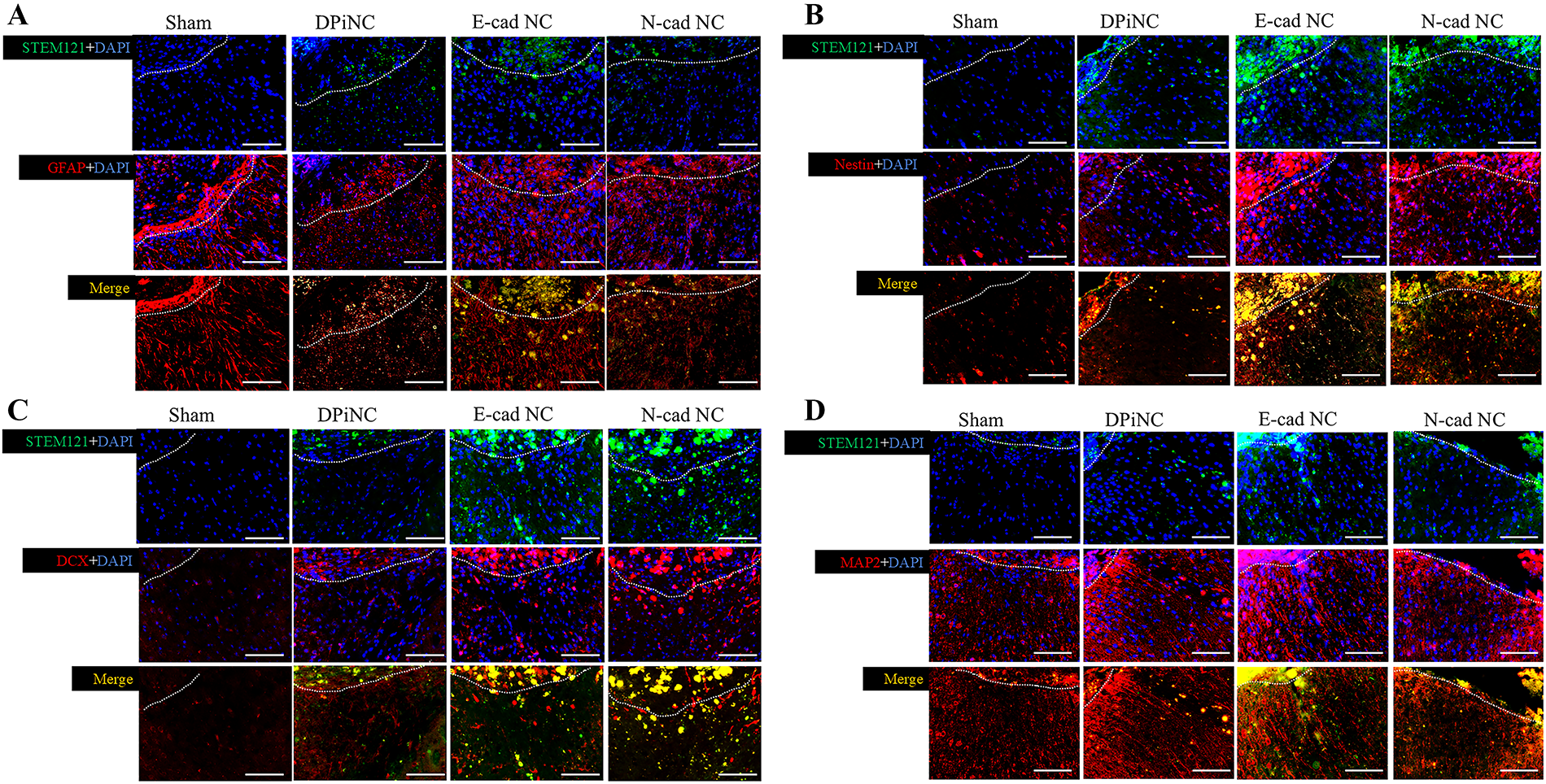
Immunostaining on day 28 post-transplantation.
Counts of transplanted cells
STEM121-positive cell counts did not differ among the groups (p = 0.209). However, the cell survival rate was higher in the N-cad NC group than in the DPiNC group, suggesting superior post-transplantation resilience (Fig. 10).
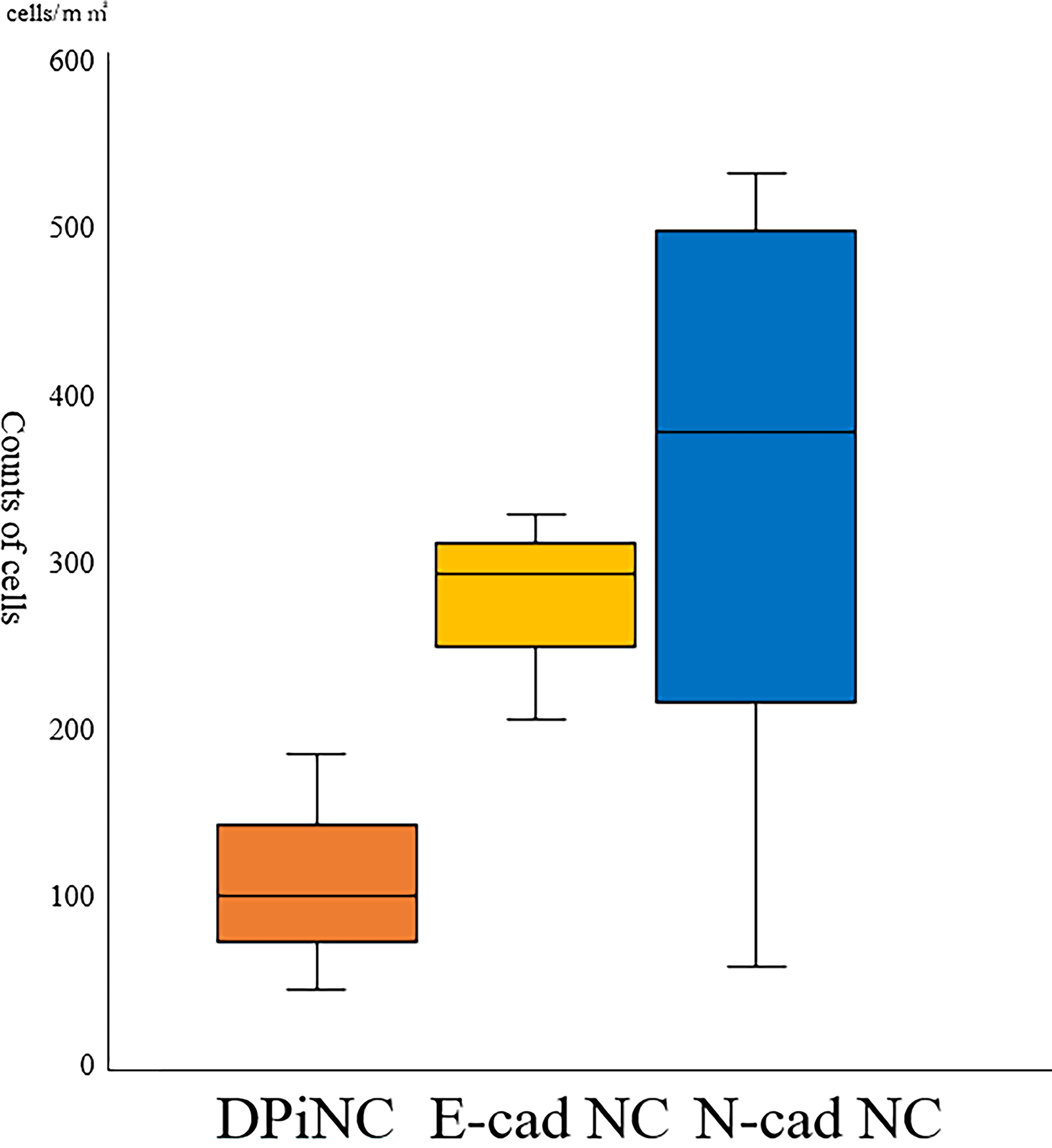
Survival of transplanted cells on day 28 post-transplantation. n = 3 per group. DPiNC, dental pulp-derived induced neural cell group; E-cad NC, dental pulp-derived induced neural cell cultured with E-cadherin group; N-cad NC, dental pulp-derived induced neural cell cultured with N-cadherin group.
Discussion
Our study, which focused on controlling cell maturity, demonstrated that the intracerebral transplantation of E-cad NCs enhances motor and sensory reflex functions and improves the survival of transplanted cells in immunodeficient mice after TBI. E-cad NC populations contain a higher proportion of immature cells than DPiNC populations, and they secrete more neuroprotective and growth factors, and they exhibit reduced cellular damage even in hypoxic conditions. Similarly, the intracerebral transplantation of N-cad NCs improved functional recovery in immunodeficient mice after TBI. Compared with E-cad NCs, N-cad NCs include mature neural cells, and despite the lack of a significant difference, many surviving cells were observed 28 days post-transplantation. The acute phase of TBI presents an environment with numerous unfavorable factors for cell transplantation, making effective regenerative medicine challenging. However, both E-cad NCs and N-cad NCs displayed have shown potential for functional improvement even when transplanted into such environments.
Regenerative medicine for TBI remains difficult, especially in the acute phase. Clinical trials of SB623, an allogeneic modified bone marrow-derived mesenchymal stromal/stem cell line, have displayed promising results, but these trials were conducted in patients with chronic-phase TBI. 1 The challenging environment for transplanted cells in the acute phase remains a significant obstacle to achieving regenerative medicine.
E-cad is expressed in immature neural cells and neural precursors. 37 Immunostaining revealed that E-cad NCs strongly expressed markers of immature neural cells, such as βIII-tubulin and DCX, whereas the expression of MAP2, a marker of mature neural cells, was weak. N-cad is also expressed in neural precursors and immature neural cells. 27 In our study, N-cad NCs exhibited different properties compared with DPiNC. Immunostaining revealed weaker expression of βIII-tubulin and DCX and stronger expression of MAP2.
The results of immunostaining under standard and OGD/R culture suggested that E-cad NCs maintained an immature phenotype, whereas N-cad NCs promoted neural maturation. These phenotypic characteristics, consistent across normoxic and hypoxic environments, suggest that E-cad NCs and N-cad NCs preserved their intended properties under stress. This stability under hypoxia emphasizes the potential utility of these cells in acute-phase TBI. We also assessed the neurotrophic and growth factors secreted by DPiNCs, E-cad NCs, and N-cad NCs, revealing differences in their secretory profiles. It is well known that DPSCs and other mesenchymal stem cells (MSCs) participate in tissue regeneration and functional recovery through mechanisms mediated by neurotrophic and growth factors.38–41 Previous reports revealed that administering neurotrophic and growth factors derived from MSCs to TBI model mice inhibited neural cell apoptosis caused by secondary brain damage and promoted functional recovery.42,43 The expression of neurotrophic and growth factors is significantly higher in DPSCs than in bone marrow-derived stem cells and adipose tissue-derived MSCs.44,45 In cases of spinal cord injury, these factors have proven effective in inhibiting the apoptosis and degeneration of neural cells, protecting motor and sensory neurons, and promoting axon regeneration.46–49
The effects of MSCs on tissue regeneration and functional recovery involving neurotrophic and growth factors are well known.38–41 Previous reports found that administering neurotrophic and growth factors derived from MSCs to TBI model mice inhibited neural cell apoptosis caused by secondary brain damage and improved functional recovery.42,43 We also assessed the neurotrophic and growth factors secreted by DPiNCs, E-cad NCs, and N-cad NCs, revealing differences in their secretory profiles.
Furthermore, gene expression was analyzed in E-cad NCs and N-cad NCs using microarray. Compared with the findings in DPiNCs, both E-cad NCs and N-cad NCs strongly expressed genes related to neural protection and neural/axonal regeneration. CHI3L1 is known to suppress inflammatory responses in neural diseases, particularly in conditions associated with neurological inflammation such as TBI, and contribute to the survival and regeneration of neural cells. 50 CRYAB is known for its anti-inflammatory effects and the ability to inhibit apoptosis.51,52 Although the effects on TBI of some proteins are not clear, both E-cad NCs and N-cad NCs have potential for neural protection and neural/axonal regeneration.
Additionally, to model secondary brain injury, the cells were subjected to hypoxia followed by reoxygenation, and their changes were assessed. The extent of cell damage was assessed by the time-dependent progression of axon length. After OGD/R, DPiNC exhibited a decrease in axon length over time, indicating cellular damage associated with the hypoxic environment. By contrast, axonal length was increased after OGD/R in both E-cad NCs and N-cad NCs, suggesting they are less affected by hypoxic conditions than DPiNCs. Immunostaining of each cell type indicated that E-cad NCs strongly expressed βIII-tubulin and DCX, whereas N-cad NCs strongly expressed MAP2, suggesting that they maintain similar properties even after OGD/R.
Although the in vivo TBI model and in vitro OGD/R model represent different aspects of brain injury, we used the OGD/R model as a simplified system to selectively evaluate cellular responses to hypoxic stress, which is a critical component of secondary injury after TBI. This approach complements the in vivo findings and provides mechanistic insights into the resilience of the transplanted cells under ischemic conditions.
Furthermore, to induce focal TBI, we selected the cold injury model due to well-defined lesion borders. Unlike widely used models such as the controlled cortical impact model, cold injury produces localized cortical damage with minimal hemorrhage, facilitating consistent cell transplantation and evaluation. Thus, although it does not replicate mechanical trauma, it effectively mimics secondary injury processes making it suitable for assessing cell-based therapies in the acute phase of TBI.
For the purpose of regenerative medicine in acute-phase TBI, cell transplantation was performed 2 days post-TBI in mice. Mice implanted with E-cad NCs exhibited significant improvements in motor function and sensory-motor reflex symptoms compared with sham mice. Specifically, in terms of motor function, the E-cad NC group displayed significantly better symptom improvement than the DPiNC group. In addition, the improvements in motor function and sensory-motor reflexes were better in the E-cad NC group than in the N-cad NC group. This supports the hypothesis that immature neural cells are advantageous in acute-phase TBI.
However, no difference was observed in anxiety symptoms between the transplantation groups and the sham group. It is possible that the improvement in paralysis in the transplantation groups led to increased activity, resulting in more marbles being buried, which was interpreted as increased anxiety. 53 To strictly compare improvements in anxiety symptoms, it is necessary to use a trauma model mouse that does not exhibit paralysis symptoms.
The transplanted cells were present in the mouse brain for 28 days post-transplantation, spreading from the transplantation site to the bottom of the contusion. It became clear that the cells differentiated into both immature and mature neural cells, in addition to extensive differentiation into astrocytes. Because the brain requires many astrocytes to maintain a single neuron, 54 the differentiation of the transplanted cells into both neuronal and astrocytic cells was considered an important finding. 54 However, no significant difference was observed for the number of surviving transplanted cells at the contusion base among the groups. Numerically, the number of surviving cells was lowest in the DPiNC group, followed by the E-cad NC group and N-cad NC group. Previous studies reported that the transplantation of neural progenitor cells with N-cad supplementation into animals with neonatal cortical brain injuries efficiently promoted neuronal regeneration and functional recovery, with cells migrating to the injury site while binding to N-cad. 55 Furthermore, the effect of N-cad on cell transplantation for TBI has been reported to involve the migration of transplanted cells from the transplantation site to the cortex and the promotion of maturation. 56 Thus, N-cad NCs are likely to migrate from the transplantation site to the contusion and participate in neuronal regeneration following TBI.
As mentioned previously, differences were observed in the neurotrophic and growth factors secreted by E-cad NCs and N-cad NCs in vitro. N-cad NCs secreted high levels of BDNF and GDNF. BDNF is critical for the survival of neurons, synaptic plasticity, and the growth of neural projections and axons, exerting strong effects on neuronal survival in TBI.57,58 GDNF is known to promote the survival of neurons and enhance the effects of neuronal migration.58,59 Therefore, N-cad NCs, which secreted high levels of BDNF and GDNF, might be more advantageous regarding cell survival in TBI than E-cad NCs.
Our study suggested that E-cad NCs promote functional recovery by maintaining immature neural characteristics and enhancing neuroprotective factor secretion in the acute TBI environment. Conversely, N-cad NCs, exhibiting a more mature phenotype, support regeneration via neurotrophic factor secretion and greater post-transplant survival. These results imply that E-cad and N-cad support neural repair through complementary mechanisms. Although tested separately, combining both—through co-culture, sequential conditioning, or cadherin-functionalized biomaterials—could enhance therapeutic efficacy. This perspective highlights the translational potential of cadherin-mediated modulation in regenerative strategies. Our study had several limitations. First, the dental pulp used in this study was derived from a single subject. Therefore, the generalizability of the findings to all DPSCs remains unclear. Additionally, the DPSCs and DPiNCs used in this study were obtained through a specific differentiation induction method, and it is unknown whether similar results can be achieved using other induction methods. Although DPSCs represent a type of MSCs derived from the neural crest, the results might not be extrapolatable to MSCs derived from other origins.
For comparison with the cell transplantation groups, a sham group was also created. We acknowledge that injecting 2 μL of Hanks’ solution in the sham group could have served as a more appropriate vehicle control.
Moreover, the long-term persistence of symptom improvement beyond 28 days post-transplantation is uncertain, necessitating further follow-up. There was also a gap in comparing the long-term effectiveness and survival of E-cad NCs and N-cad NCs beyond the acute phase. Additionally, this specific TBI model might not encompass the variety of TBI manifestations, such as diffuse injuries common in accidents, limiting the applicability of our findings. Finally, we did not investigate the integration of the transplanted cells with the host’s neural network, requiring additional studies to understand their connectivity and interactions within the neural system.
Conclusion
In conclusion, our study highlighted the potential utility of both E-cad NCs and N-cad NCs in regenerative medicine approaches for acute TBI. E-cad NCs were typified by their ability to proliferate while maintaining immaturity, offering advantages in hypoxic environments and exerting neural protection and regenerative effects. Similarly, the N-cad NCs promoted maturity, also offering advantages in hypoxic environments, and they exerted neural protection and regenerative effects.
Transparency, Rigor, and Reproducibility Summary
Data collection methods, data processing steps, and analysis plans were prespecified and are available upon request or in the supplementary materials.
Rigor: To ensure methodological rigor, sample size calculations were conducted to achieve adequate statistical power. Standardized procedures were used for data collection, and all assessments were conducted by trained personnel following validated protocols.
Reproducibility: The data and statistical code for all analyses in this study are available upon request. We encourage replication efforts by providing detailed descriptions of all procedures and analysis approaches in the methods section. Additionally, any deviations from the prespecified analysis plan are documented and justified.
Authors’ Contributions
J.N.: Conceptualization, data curation, formal analysis, investigation, methodology, visualization, and writing—original draft. Aiki M.: Conceptualization, data curation, funding acquisition, methodology, project administration, resources, and writing—review and editing. H.I.: Conceptualization, methodology, resources, validation, and writing—review and editing. T.T.: Methodology and writing—review and editing. J.T.: Resources, visualization, and writing—review and editing. A.O.: Resources, visualization, and writing—review and editing. Arnela M.: Resources and writing—review and editing. H.M.: Resources and writing—review and editing. S.T.: Resources and writing—review and editing. H.B.: Resources and writing—review and editing. T.S.-S.: Resources and writing—review and editing. T.A.: Resources and writing—review and editing. Y.M.: Writing—review and editing. E.I.: Supervision and writing—review and editing.
Footnotes
Acknowledgments
The authors thank Ms. Tsukada and Ms. Miyakawa from Graduate School of Comprehensive Human Sciences, University of Tsukuba, for their technical support.
Author Disclosure Statement
The authors declare that they have no competing interest.
Funding Information
This article was supported by Japan Society for the Promotion of Science (JSPS) KAKENHI Grant Number 20H03787, JST FOREST Program, Grant Number JPMJFR2112 to A.M., and Grant-in-Aid for Early-Career Scientists for T.T. (No. 21K16601) from the JSPS.
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.