
Abstract
Background
Esophageal cancer (EC) is a common malignant tumor of the digestive tract and an important health-related problem in many developing countries. Esophageal squamous cell carcinoma (ESCC) is the most common subtype of EC. The cancer-associated fibroblasts (CAFs) are the major stromal cells in ESCC microenvironment. They play important role in ESCC proliferation, metastasis, angiogenesis and chemotherapy resistance through paracrine processes. However, the roles of circRNAs enriched in CAF-derived exosmes have not been reported.
Objective
To explore the mechanisms of how CAF affects ESCC proliferation and metastasis through paracrine processes and to investigate the role of circRNAs enriched in CAF-derived exosomes.
Methods
Exosomes were isolated from the conditional medium of CAF using differential ultracentrifugation, and then validated by Nanosight analysis. Exosome secretion inhibitor-GW4869 validates the pro-carcinogenic role of exosomes. The qRT-PCR showed the highest expression of circ_0076535 in the exosomal CircRNA, and knockdown of it confirmed its function. Online bioinformatics tool was utilized to predict the potential target gene of circ_0076535, and captured miR-145-5p as the target gene with high predictive value. The targeting association between miR-145-5p and circ_0076535 is further confirmed by the dual luciferase reporter experiment. The stimulation of tumour development and EMT by the CAF-derived exosome circ_0076535 is further validated in vivo.
Results
In our research, we found that CAF-derived exosomes increased proliferation, migration, invasion and EMT in ESCC cells. Circ_0076535 was highly enriched in CAF-exosomes and transferred into ESCC cells directly depend on internalization of exosomes. CAF-exosomal circ_0076535 increased the level of circ_0076535 in ESCC cells and induced EMT. Mechanistic experiments revealed circ_0076535 acted as a sponge to absorb miR-145-5p and activated NF-κB signaling pathway.
Introduction
Esophageal cancer (EC) is one of the most common malignant tumors in the world. It is estimated that there were at least 600,000 new cases and 500,000 deaths of esophageal cancer worldwide in 2018. Approximately 604,000 new cases of oesophageal cancer (EC) are expected to be diagnosed in 2020, resulting in 537,000 cancer deaths worldwide. 1 Currently, the incidence of esophageal cancer ranks seventh and the mortality rate ranks sixth among all kinds of malignant tumors, creating a serious disease burden for humans. 2 Esophageal squamous cell carcinoma (ESCC) is the most common histological type of EC, accounting for approximately 80% of all cases worldwide. 3 ESCC is the result of multiple factors such as heredity, behavior, diet, mainly including family history of EC, smoking, drinking, liking to eat hot food and drink and so on. 4 Due to lack of obvious clinical symptoms in the early stage of ESCC, many patients were diagnosed as being at an advanced stage, which lead to a poor prognosis and high mortality. The 5-year overall survival rate of ESCC is less than 30%, which is even lower in some developing countries. 5 Since early detection is essential to improve the prognosis and mortality of ESCC patients, it is very urgent to identify new effective therapeutic targets in ESCC.
In recent years, targeting tumor microenvironment (TME) has become a focus in cancer therapy. TME is a complex tissue environment, including extracellular matrix and various stromal cells, such as cancer-associated fibroblasts (CAFs), macrophages, inflammatory cells, endothelial cells, adipocytes and mesenchymal stem cells.6–8 CAFs are one of the most important cellular components in TME, and it has been reported that CAFs play a vital role in the progression of ESCC. 9 Zhang et al. suggested that CAFs induced lncRNA DNM3OS expression through a PDGFβ/PDGFRβ/FOXO1 signaling pathway to regulate radio response in ESCC. 10 And Kashima et al. found that CAFs extremely facilitated the lymph nodes of ESCC. 11 It is a well-accepted conclusion that CAFs effectively regulate the malignant phenotypes of cells to affect the progression of ESCC. However, the mechanisms haven’t been revealed clearly.
Exosomes are membranous vesicles with a diameter of 30–100 nm secreted by diverse cells, which can transmit information between different cells. 12 It is reported that exosomes contain lots of mRNAs, proteins, lipids and non-coding RNAs. 13 There have been many researches about the regulatory of exosomes derived from CAFs in various tumor progression. For instance, CAFs promoted oral cancer cells proliferation and metastasis via transferring exosomal miR-34a-5p. 14 In colorectal cancer, exosomal lncRNA H19 derived from CAFs induced the stemness and chemoresistance of CC cells. 15
Circular RNA (circRNA) is one type of non-coding RNAs with a covalent closed loop structure and lack of 5′-caps or 3′-poly(A) tail. 16 CircRNA is featured by stable structure, highly conserved sequence and high abundance, and these characteristics made circRNA focused in recent years. 16 It has been found that there is a large amount of circRNA contained in exosomes stably, and could be delivered into target cells, especially cancer cells, to affect the biological behaviors. Gastric cancer cells exosomes contained with ciRS-133 promoted the differentiation of preadipocytes into brown-like cells. 17 Exosomes derived from hepatocellular carcinoma cells transferred circRNA Cdr1as to surrounding normal cells to induced proliferation and migration and accelerate the progression of HCC in a miR-1270/AFP pathway-dependent manner. 18 However, there are no research on the functions and mechanisms of exosomal circRNA in the progression of ESCC.
In this study, we aimed to explore the effects of CAF-derived exosomal circ_0076535 functionally and mechanically impacting on ESCC. Firstly, we verified that CAF-derived exosomes induce EMT in ESCC cells. Subsequently, we screened the highly expressed circRNA in CAF-exosomes to pick out the target gene circ_0076535. Functionally, CAF-derived exosomal circ_0076535 promoted ESCC proliferation, migration, invasion and EMT in vivo and in vitro. And mechanistic investigations revealed that CAF-derived exosomal circ_0076535 acted as a sponge of miR-145-5p, which reversed circ_0076535-induced biological functions by targeting NF-B pathway in ESCC.
Methods and materials
Cell culture
Human esophageal squamous cell carcinoma cells, including TE-1 and KYSE-150 cells, are purchased from the Chinese Academy of Sciences, Shanghai, China. TE-1 and kyse-150 cells are cultured with DMEM medium supplemented with 10% FBS (fetal bovine serum, Gibco) and incubated at a humid, 5% CO2 and 37°C atmosphere.
qRT-PCR
Total RNAs are isolated with the TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. cDNAs are produced by the reverse transcript kit from Takara (Dalian, China). qRT-PCR is conducted with SYBR Green real-time PCR Kit (Qiagen, Hilden, Germany). Data were analyzed by the
CCK8 assay
ESCC cells treated with CAF-exosomes were seed into a 96-well cell culture plate in a density of 8 × 103 cells per well. Until the cells adhered, 10 μl of CCK8 reagent was add to each well at different time points, incubate at 37°C for 1 h, measure the absorbance at 450 nm in a microplate reader, and plot the cell proliferation curve.
Colony formation assay
ESCC cell were plated into six-well dishes in a density at 0.4 × 103 cells each well. The medium was updated every 2 days until the number of cells in each colony exceeds 50, colonies were fixed with 4% PFA and stained with 0.1% crystal violet at room temperature for 30 min. The colonies are randomly selected for observation and count at a fluorescence microscope.
Transwell assay
ESCC cells treated with CAF-exosomes (5 × 105 cells/ml) are suspended in serum free DMEM medium. 100 µl of the cell suspension was seeded on the upper part of a 24-well culture plate with an 8 µm Transwell chamber (Corning, USA) for 48 h, and 500 µl of DMEM medium supplemented with 10% FBS was added to the lower chamber. The chambers uniformly covered with Matrigel (BD Biosciences) are used to invasion assay. The cells on the upper surface of the insert are then removed. The insert was fixed in 4% paraformaldehyde for 15 min and then stained with 0.1% crystal violet for 5 min. The stained insert was washed with PBS and observed under a microscope. Cell migration and invasion capacities were measured using average stained cells in 5 areas. Each experiment was repeated three times.
Western blot
Total proteins were extracted from ESCC cells with radioimmunoprecipitation assay (RIPA) lysis buffer, and then centrifuged for 15 min at 12,000 rpm. The concentration of total proteins was quantified by a BCA protein assay kit (Beyotime). 30 μg of total proteins was separated by the 6%, 8% or 10% SDS–PAGE gel and then transferred to PVDF membrane (Millipore). After blocked with 5% non-fat milk for 2 h at room temperature the membrane is then incubated with specific primary antibodies. Next, the membrane is incubated with goat anti-rabbit/anti-mouse secondary antibody. The signals at protein level are visualized with the ECL Substrates (Millipore). The WB results are quantified by Image J software.
Luciferase reporter assays
Dual luciferease reporter activities are conducted in HEK-293 T cells co-transfected with the circ_0076535 (WT or Mut) and miR-145-5p mimic, inhibitor or negative control. After transfection for 48 h, luciferase activity was measured according to the manufacturer's instruction (Promega, USA).
Animal model
BALB/c-nu/nu mice purchased form Model Animal Research Center Of Nanjing University at 4-6 weeks of age were used for this study. ESCC cells treated with different CAF-exosomes (1 × 107 cells suspended in 200 μl PBS) were injected subcutaneously into mice. These mice were randomly divided into 3 groups: mock, CAFsh-NC-exosomes, CAFsh-circ_0076535-exosomes, there were 6 in each group. The mice were sacrificed after 4 weeks, and the tumor specimens were quickly removed and divided into two parts: one part for qRT-PCR and western blot analysis of circ_0076535 and EMT markers, and another part for H&E stain and immunohistochemistry.
Statistical analysis
All statistical analyses were performed using SPSS 20.0 software (SPSS, Chicago, IL, USA) and Prism version 5.0 software (Graph Pad Software, La Jolla, California, USA). Data are expressed as the mean ± SEM of at least 3 independent experiments. An unpaired t-test was performed to compare the mean of the two groups, and then a one-way ANOVA and Bonferroni multiple comparison test was used to detect differences between the two or more groups. Significant differences were defined as P < 0.05.
Results
CAF-derived exosomes induce EMT in ESCC cells
To clarify the roles of exosomes secreted by CAF in ESCC progression, the functional experiments were performed in vitro. Firstly, exosomes were isolated from the conditional medium of CAF using differential ultracentrifugation. Nanosight analysis revealed that the diameter of these nanoparticles was range in 50–100 nm, which is typical of exosomes. And exosome hallmarks CD63, CD81 and TSG101 were highly expressed in these nanoparticles, whereas weakly expressed in CAF (Figure 1A). Subsequently, PKH67 labeled-exosomes were respectively incubated with ESCC cells TE-1 and KYSE-150 to examine whether CAF-exosomes could be absorbed by ESCC cells, and the green fluorescent signal was detected both in TE-1 and KYSE-150 (Figure 1B), suggesting that ESCC cells internalized CAF-derived exosomes. Functionally, CCK8 and colony formation assay were used to assess the effect of CAF and CAF-exosomes on ESCC cells proliferation. As shown in Figure 1C and 1D, treatment with CAF or CAF-exosomes promoted TE-1 and KYSE-150 proliferation, CAF and CAF-exosomes also increased the migration and invasion of TE-1 and KYSE-150 (Figure 1E). And western blot analysis of EMT markers showed that ESCC cells treated with CAF or CAF-exosomes expressed lower E-cadherin and higher N-cadherin and Vimentin (Figure 1F). However, those promotion effects of CAF on ESCC cells proliferation, migration, invasion and EMT were significantly suppressed by GW4869, a compound that blocks exosome secretion, suggesting the induction of aggressive behaviors in ESCC cells by CAF-derived exosomes.
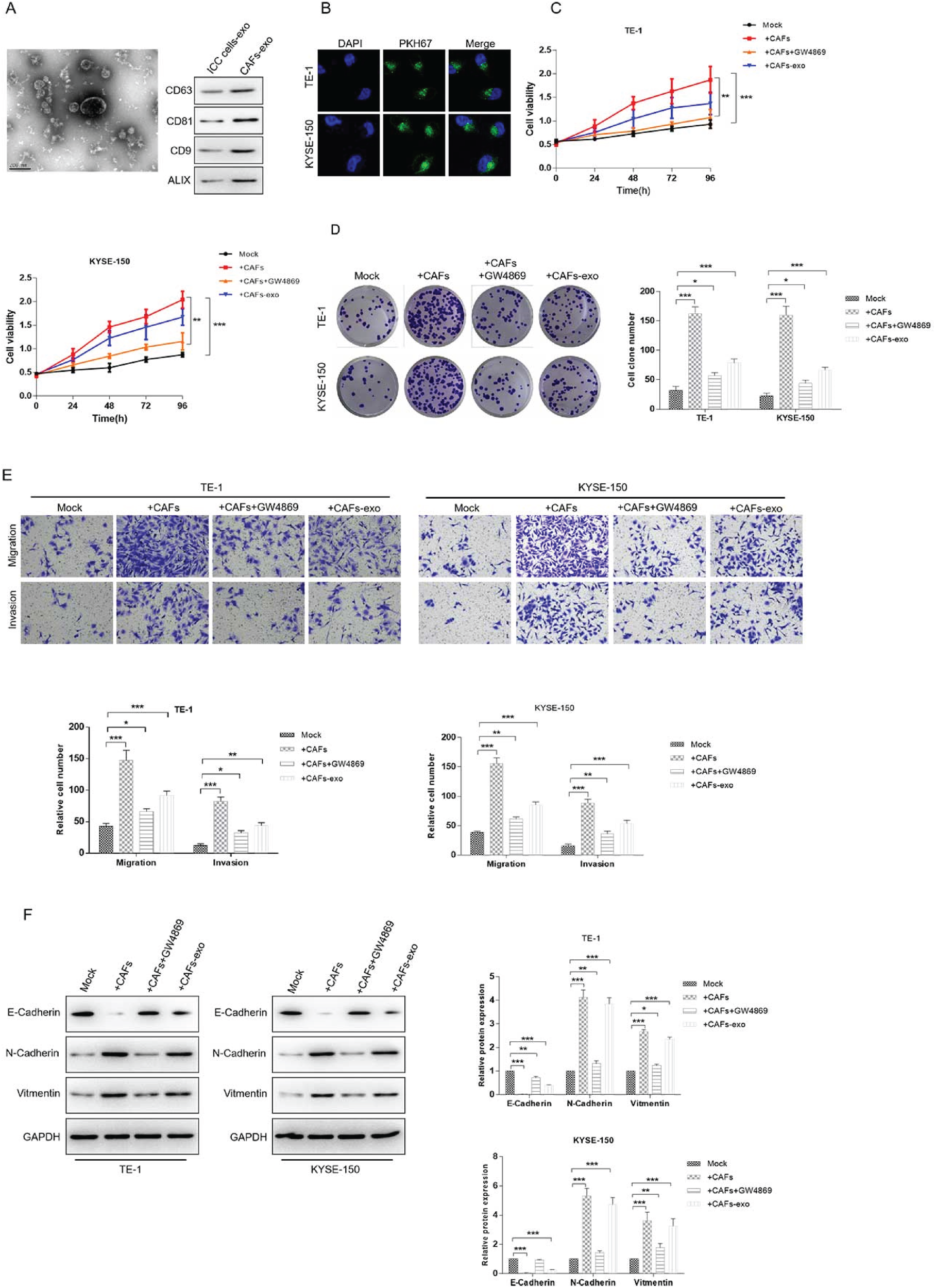
CAF-derived exosomes ninduces EMT in ESCC cells. (A) Isolation and characterization of exosomes derived from CAF. Left: particle size analysis; Right: western blot assay of exosome hallmarks. (B) PKH67-labeled exosomes are respectively co-cultured with ESCC cell lines, TE-1 and KYSE-150 for 4 h. Represent images of PKH67-labeled CAF-exosomes uptaken by ESCC cells were captured by an inverted fluorescence microscope (400×). (C-F) ESCC cells were treated with CAF, CAF-exosomes, CAF plus GW4869 or mock. CCK8 (C) and colony formation assay (D) were used to determinate the proliferation of ESCC cells. Transwell assay was applied to examine the migration and invasion abilities (E). The protein levels of EMT markers E-Cadherin, N-Cadherin and Vimentin were detected by western blot analysis (F). GAPDH was used as the internal control. *p<0.05, **p<0.01, ***p<0.001.
We analyzed the differential expressed circular RNA profile performed by Fan, 19 and detected the expression levels of six circRNAs (circ_0062459, circ_0076535, circ_0072215, circ_0042261, circ_0001946, and circ_0043603) in CAF-derived exosomes by qRT-PCR. As shown in Figure 2A, circ_0076535 was the highest expressed among 6 circRNAs. But the roles of circ_0076535 were unknown in ESCC progression. To explore whether internalization of CAF-derived exosomes by ESCC cells increases circ_0076535 expression, the level of circ_0076535 was validated in ESCC cells incubated with CAF-exosomes or not. Circ_0076535 was significantly upregulated in TE-1 and KYSE-150 cells treated with CAF-exosomes compared with that in untreated group (Figure 2B). Moreover, to verify whether the upregulation of circ_0076535 in ESCC cells was induced by direct diversion of CAF-exosomal circ_0076535, CAF were transfected with circ_0076535 shRNA or negative control (named as “CAFsh−circ_0076535”and “CAFsh−NC”), which knockdown the expression of circ_0076535 in CAF and corresponding secreted exosomes (Figure 2C). As excepted, circ_0076535 expression was obviously decreased in ESCC cells co-cultured with CAFsh−circ_0076535-exosomes (Figure 2D). The above results indicated that CAF-derived exosomal circ_0076535 was transferred in ESCC cells directly and increased circ_0076535 expression.
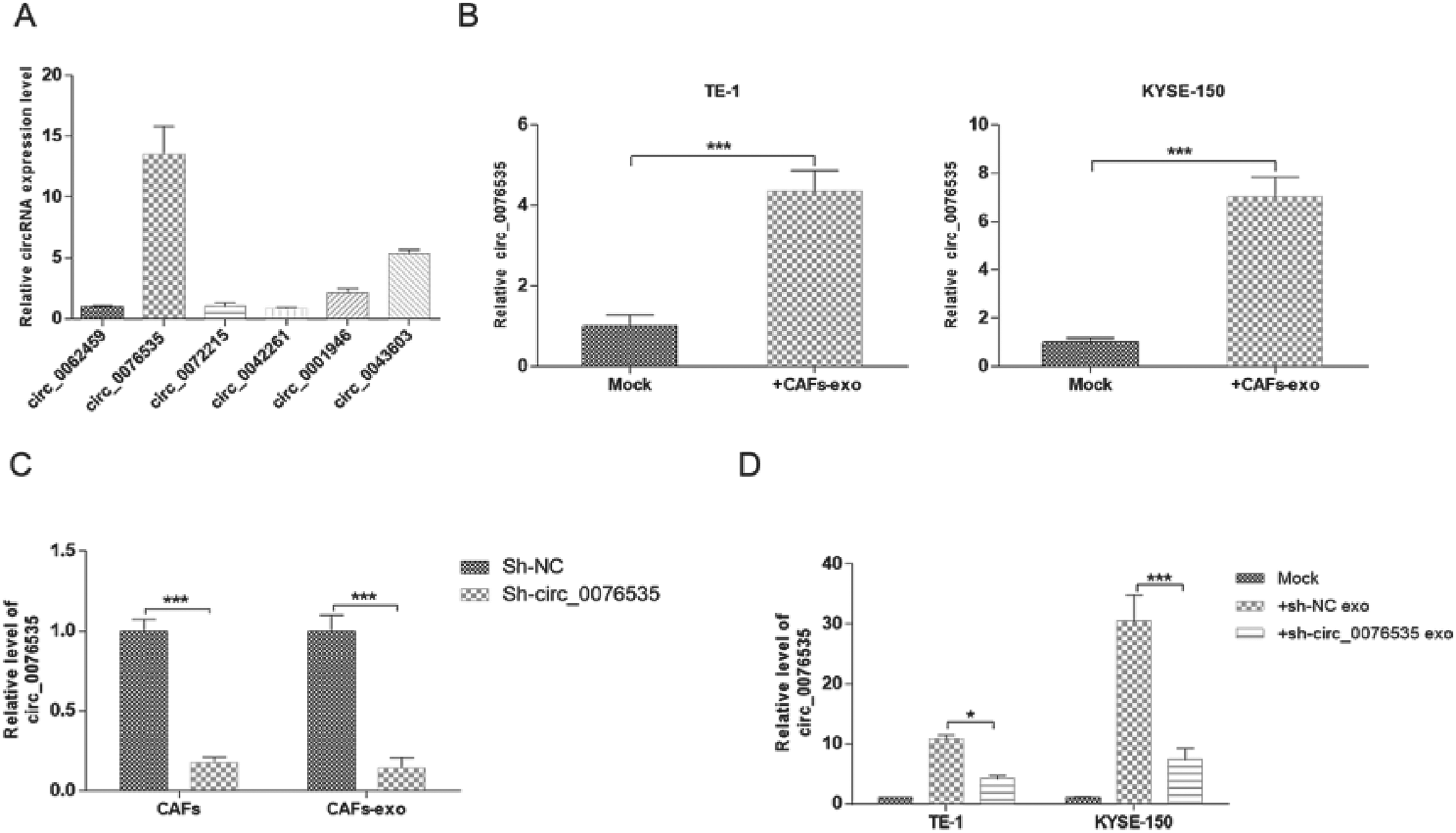
CAF-derived exosomal circ_0076535 is transferred into ESCC cells directly. (A) Enrichment of differentlly expressed circRNAs in CAF-derived exosomes by qRT-PCR. (B) Relative expression of circRNAs in ESCC cells co-cultured with CAF-exosomes. (C) Relative expression of circ_0076535 in CAF transfected with circ_0076535 shRNA or negative control, and their respective exosomes. (D) Effect of low-circ_0076535 expressed CAF-derived exosomes on circ_0076535 expression of ESCC cells. GAPDH was used as the internal control. *p<0.05, **p<0.01, ***p<0.001.
Then, the role of CAF-exosomal circ_0076535 were explored in ESCC progression. The results of CCK8 and colony formation assay indicated that CAFsh−circ_0076535-exosomes treatment inhibited ESCC cells proliferation (Figure 3A and 3B). CAFsh−NC-exosomes induced ESCC cells migration and invasion, while circ_0076535 knockdown in CAF-exosomes effectively inhibited migration and invasion (Figure 3C). Similarly, EMT markers N-cadherin and Vimentin were significantly downregulated, while E-cadherin was upregulated in ESCC cells incubated with CAFsh−circ_0076535-exosomes, compared to these in ESCC cells incubated with CAFsh−NC-exosomes (Figure 3D). These results demonstrated that CAF-exosomes induce EMT in ESCC cells via transferring circ_0076535.
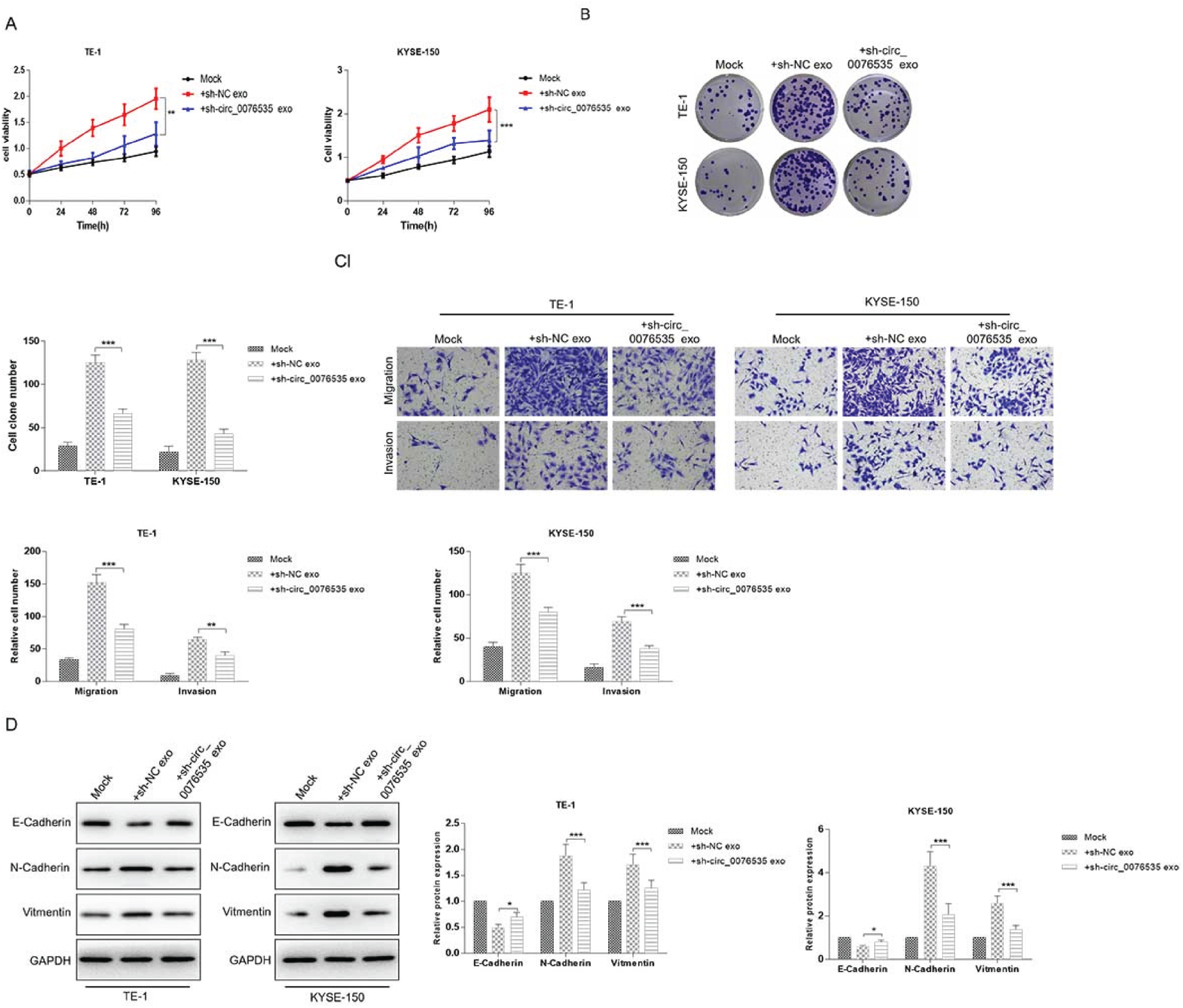
CAF-derived exosomal circ_0076535 induces EMT in ESCC cells. ECSS cells (TE-1 and KYSE-150) were co-cultured with exosomes derived from circ_0076535 knockdown CAF or NC CAF, ECSS cells with no-treatment were used as the blank control group. (A-B) The proliferation of ESCC cells were determined by CCK8 (A) and colony formation assay (B). (C) ESCC cells migration and invasion were measured by transwell assay. (D) Western blot analysis of EMT markers. GAPDH was used as the internal control. *p<0.05, **p<0.01, ***p<0.001.
Online bioinformatics tool was utilized to predict the potential target gene of circ_0076535, and captured miR-145-5p as the target gene with high predictive value. The complementary sequences of circ_0076535 and miR-145-5p were represented in Figure 4A. To determine whether circ_0076535 serve as a sponge to absorb miR-145-5p, the cire_0076535 luciferase reporter containing the predicted binding sequences or mutated sequences (named as “circ_0076535 WT” and “circ_0076535 MUT”) was conducted and dual-luciferase reporter assay was performed. The luciferase activity of circ_0076535 WT was inhibited by miR-145-5p mimic and active by miR-145-5p inhibitor, however, the luciferase activity of circ_0076535 MUT had no change regardless of co-transfection with miR-145-5p mimic or inhibitor (Figure 4B). In addition, miR-145-5p expression was significantly downregulated in ESCC cells incubated with CAFsh−NC-exosomes, but CAFsh−circ_0076535-exosomes could reduce the effect (Figure 4C). These data indicated that CAF-exosomal circ_0076535 act as a sponge to absorb miR-145-5p in ECSS cells.
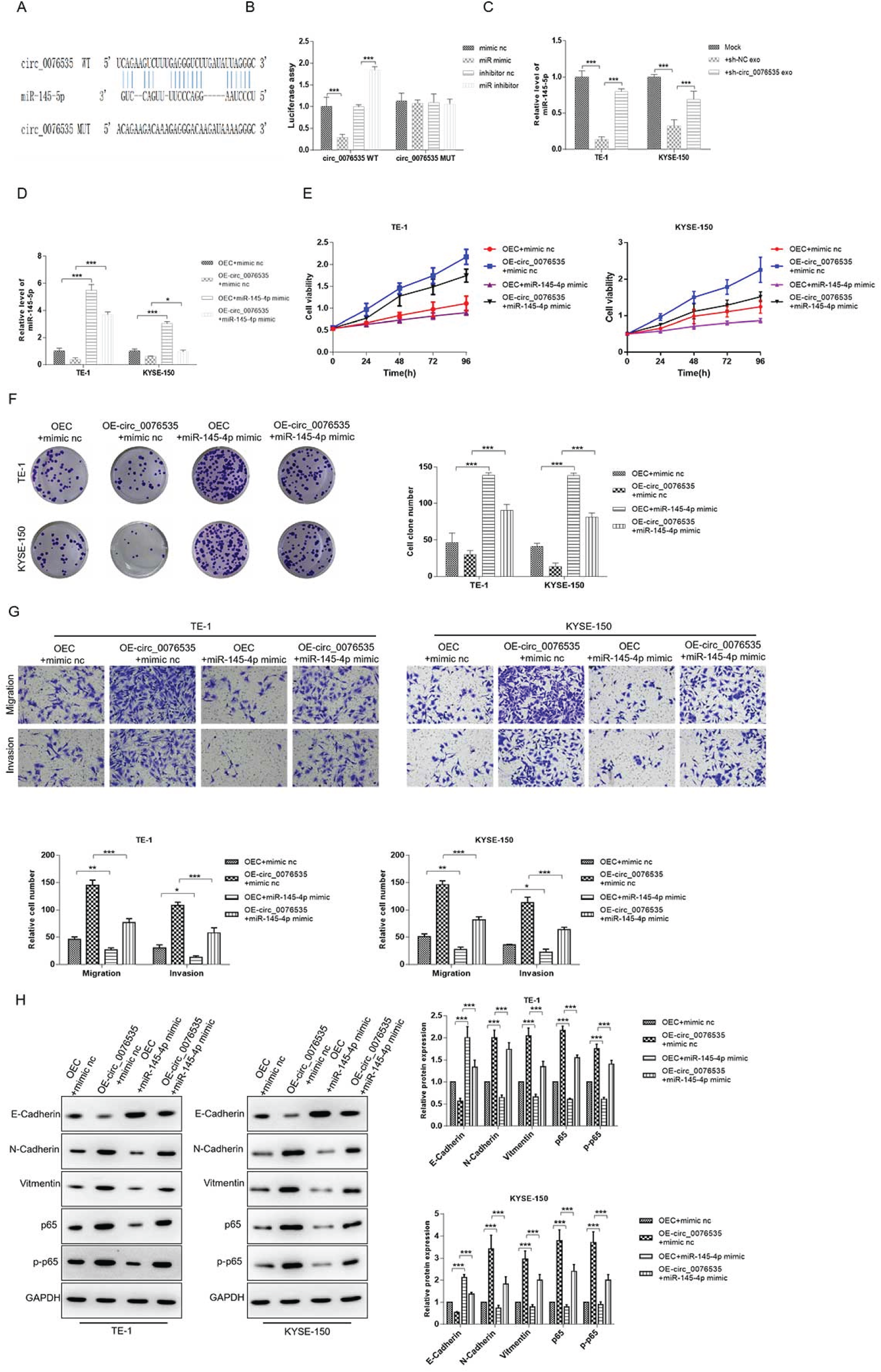
miR-145-5p attenuates circ_0076535-induced EMT via inactivating NF-κB pathway in ESCC. (A) Schematic diagram of the binding sites of circ_0076535 and miR-145-5p. (B) The direct binding of circ_0076535 and miR-145-5p was verified by dual-luciferase reporter assay. (C) Effect of low-circ_0076535 expressed CAF-derived exosomes on miR-145-5p expression of ESCC cells. (D-I) ESCC cells (TE-1 and KYSE-150) were co-transfected with circ_0076535 overexpression plasmid or vector and miR-145-5p mimic or mimic nc for 48 h. CCK8 (E) and colony formation assay (F) were performed to assess the proliferation of cells. Transwell assay (G) was used to measure the migration and invasion of ESCC cells. Western blot (H) was used to detected the protein levels of E-Cadherin, N-Cadherin, Vimentin, p65 and p-p65. GAPDH was used as the internal control. *p<0.05, **p<0.01, ***p<0.001.
Further, the regulation of circ_0076535/miR-145-5p axis in ESCC cells was explored. ESCC cells TE-1 and KYSE-150 were separately co-transfected with circ_0076535 overexpression plasmid or negative control and miR-145-5p mimic or mimic nc. The promotion effects of ESCC cells proliferation, migration, invasion and EMT induced by circ_0076535 overexpression was effectively suppressed by miR-145-5p mimic (Figure 4E-H). Moreover, it has been comfirmed that miR-145-5p was a tumor suppressor that inhibited ESCC cells proliferation, migration, invasion and EMT through activating NF-κB pathway. 20 So, NF-κB pathway was detected by western blot, and the result revealed that p65 and p-p65 were obviously upregulated in circ_0076535 overexpressing ESCC cells, and the upregulation was reversed by miR-145-5p mimic (Figure 4H). These results suggested that miR-145-5p attenuates circ_0076535-induced EMT via inactivating NF-κB pathway in ESCC.
The roles of CAF-derived exosomal circ-007635 in vivo were explored in mice. ESCC cells TE-1 and KYSE-150 cells treated with CAFsh−NC-exosomes, CAFsh−circ_0076535-exosomes were injected into the subcutaneous of nude mice, and the xenografts were collected after 4 weeks of injection. The results revealed that the CAFsh−NC-exosomes treatment group expressed the highest circ_0076535 expression and CAFsh−circ_0076535-exosomes group expressed lowest (Figure 5A). The volume and weight of tumor nodules in CAFsh−NC-exosomes treatment group were greater than that of mock (Figure 5B and 5C). Result of H&E staining showed a weak malignant morphology in CAFsh-NC-exosomes treatment group (Figure 5D). Further, the expression levels of EMT markers were detected by IHC and western blot, N-cadherin and Vimentin were significantly downregulated, and C-cadherin was upregulated in CAFsh−NC-exosomes group, which were reversed in CAF sh−circ_0076535-exosomes group (Figure 5E and 5F). Results of in vivo experiments showed that CAF-derived exosomal circ_0076535 induces tumor formation and EMT.
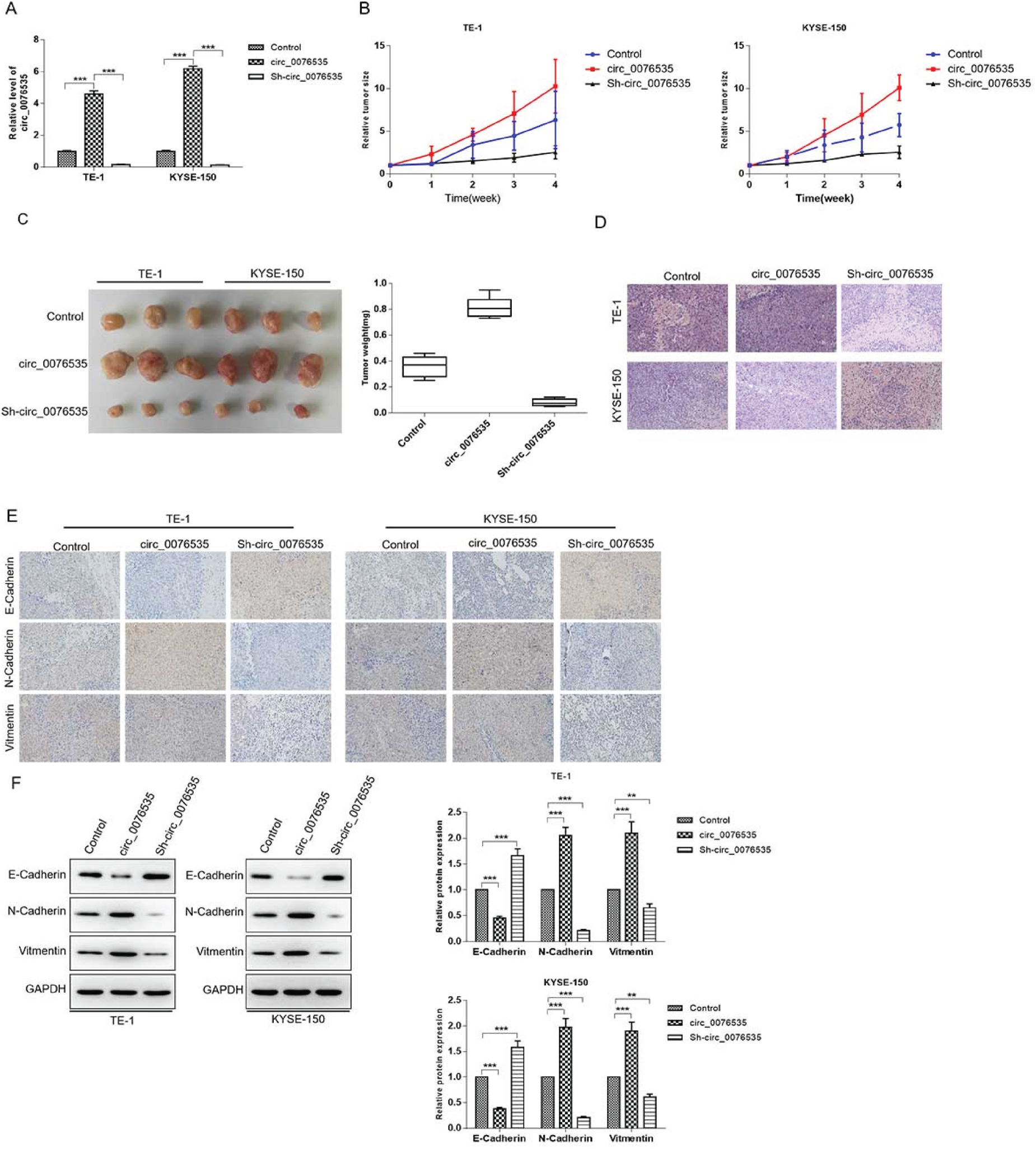
Knockdown of circ_0076535 suppresses tumor formation and EMT in vivo. ECSS cells co-cultured with CAFsh−NC-exosomes, CAFsh−circ_0076535-exosomes or no treatment were injected subcutaneously in nude mice, n = 3 each group. (A) Representative images of tumor morphology (left) and tumor volumes (right). (B) Tumor volume growth curve. (C) Representative images of H&E staining. (D-E) Expression levels of E-cadherin, N-cadherin and Vimentin in xenografts were detected by immunohistochemical (E) and western blot (F). GAPDH was used as the internal control. Scar bar = 50 μm. *p<0.05, **p<0.01, ***p<0.001.
As the most important non-lymphocyte in TME, cancer fibroblast associated cells (CAFs) are involved in multiple processes of tumor development, including cancer cells proliferation, metastasis, drug resistance, and epithelial to mesenchymal transition (EMT). 8 Current research suggested that CAF plays the regulatory roles by releasing CAFs-specific proteins, cytokines or macrovesicles. 21 In breast cancer, CAFs-secreted IL32 specifically interacted with integrin β3 to activate p38/MAPK signaling pathway, thus promoting migration and EMT. 22 In colorectal cancer METTL3 in cancer-associated fibroblasts-derived exosomes promotes the proliferation and metastasis and suppresses ferroptosis by eliciting ACSL3 m6A modification. 32 Meanwhile, the research showed that CAF-derived exosomal LINC01833 promotes the occurrence of non-small cell lung cancer through miR-335-5p -VAPA axis. 33 Exosomes derived from senescent CAFs promoted the proliferation of breast cancer cells by delivering EPHA2. 23 In addition, the effects of CAFs on the progression of esophageal squamous cell carcinoma has been reported. Du et al. indicated that hydrogen peroxide-inducible clone 5 (HIC-5) released by CAFs increased ESCC cells migration and invasion via regulating cytokines and modifying ECM. 24 Kashima et al. suggested that CAFs extremely facilitated the lymph nodes of ESCC, 11 but the underlying mechanisms have not been elucidated. Our study verified that the promotion effects of CAFs on ESCC cells proliferation, migration, invasion and EMT, and GW4869 could effectively suppress the promotion, which means that CAF-derived exosomes induced EMT in ESCC.
In addition to proteins, CAFs-derived exosomes also transport non-coding RNA into the receptor cells to regulate the biological behaviors. 25 CAFs-derived exosomal miR–382–5p enhanced oral squamous cell carcinoma cells migration and invasion. 26 Domiammma E found that CAFs-derived exosomes which contained miR-21, miR-278e and miR-143 significantly promoted breast cancer cells EMT. 27 Compared with linear RNA (miRNA or lncRNA), circRNA forms a special covalently closed loop structure without a 5′cap end and 3′poly(A), which makes circRNA more resistant to exonuclease and relatively highly abundant in the cytoplasm. 28 These excellent characteristics makes circRNAs gradually become new diagnostic markers and therapeutic targets for various cancers.
CircRNAs play critical roles in gene regulation by adsorbing miRNAs, regulating splicing or transcription, and other mechanisms. 29 It has been reported that circRNAs are contributed to the development of ESCC. CircGSK3β was upregulated in ESCC patients and was positively related with advanced clinical stage and poor prognosis. Further investigation suggested that circGSK3β enhanced ESCC metastasis through bounding to GSK3β and inhibiting GSK3β activity directly. 30 Moreover, the most renowned circRNA, ciRS-7 (alternate named CDR1as) promoted ESCC cells proliferation, migration and invasion via sponging miR-876-5p to promote MAGE-A family members expression. 31 However, there are few researches about the enrichment of circRNA in exosomes, and there is no report about circRNA in CAFs-derived exosomes.
Conclusion
In this study, we demonstrated that circ_0076535 was enriched in CAF-derived exosomes, and was transferred from CAFs to ESCC cells, thus regulating the level of circ_0076535 in ESCC cells. Functionally, elevated circ_0076535 promoted the proliferation, migration, invasion and EMT in ESCC cells both in vivo and in vitro. Mechanistically, CAF-exosomal circ_0076535 acted as a sponge to absorb miR-145-5p, and thus activated NF-κB signaling pathway. miR-145-5p overexpression attenuated circ_0076535-induced EMT via inactivating NF-κB pathway in ESCC. In conclusion, CAF-exosomal circ_0076535 was an oncogene in ESCC progression via targeting miR-145-5p/NF-κB pathway. CAF-exosomal circ_0076535 might be an effective biomarker for early diagnosis and treatment of ESCC. Although in this study, we demonstrated that circ_0076535 in the CAF-derived exosome could induce ESCC cell proliferation, migration. However, it is unknown whether it also plays a role in other cancer cells. In the later study, we plan to extract it and culture it with other cancer cells to test whether it also has an inducing effect on other cancer cells.
Footnotes
Acknowledgments
The authors have no acknowledgments.
Funding
This work was supported by Anhui Medical University School-level fund (2022xkj157).
Conflict of interest
The authors declare that they have no conflict of interest.