
Abstract
In the longitudinal study reported here, we examined genetic and caregiving-based contributions to individual differences in infant attachment classifications. For 154 mother-infant pairs, we rated mothers’ responsiveness to their 6-month-old infants during naturalistic interactions and classified infants’ attachment organization at 12 and 18 months using the Strange Situation procedure. These infants were later genotyped with respect to the serotonin-transporter-linked polymorphic region (5-HTTLPR). Maternal responsiveness uniquely predicted infants’ attachment security. Infants’ 5-HTTLPR variation uniquely predicted their subtype of attachment security at 12 months and their subtype of attachment insecurity at 12 and 18 months. The short allele for 5-HTTLPR was associated with attachment classifications characterized by higher emotional distress. These findings suggest that 5-HTTLPR variation contributes to infants’ emotional reactivity and that the degree to which caregivers are responsive influences how effectively infants use their caregivers for emotion regulation. Theoretical implications for the study of genetic and caregiving influences are discussed.
Individual differences in infant attachment are one of the foundations for variations in social and emotional adjustment across the life course. Infant attachment classifications, as assessed with the Strange Situation procedure (Ainsworth, Blehar, Waters, & Wall, 1978), predict behavior problems during childhood (Fearon, Bakermans-Kranenburg, van IJzendoorn, Lapsley, & Roisman, 2010), symptoms of psychopathology in adolescence (Carlson, 1998), and the quality of interpersonal relationships across childhood and extending into adulthood (Berlin, Cassidy, & Appleyard, 2008). Although these associations are sometimes contingent on subsequent developmental experiences (e.g., Salvatore, Kuo, Steele, Simpson, & Collins, 2011), attachment classifications in infancy nonetheless mark the initiation of developmental trajectories that probabilistically lead to differences in socioemotional functioning. For this reason, it is important to understand the factors that contribute to variations in infant attachment.
Two contrasting explanations have been proposed to account for individual differences in infant attachment. Attachment theory conceptualizes variations in infant attachment as reflecting differences in the quality—or security—of the infant-caregiver relationship. Thus, attachment theory emphasizes the importance of infant-caregiver interactions—namely, the caregiver’s responsiveness to the infant’s signals—for promoting secure attachment (Ainsworth et al., 1978; Bowlby, 1982; Sroufe, 1985). Other accounts have conceptualized infant attachment classifications as a reflection of infants’ temperament, or biologically based emotional predispositions (Goldsmith & Alansky, 1987; Kagan, 1982). Debates involving the relative contributions of caregiver responsiveness and infant temperament to infant attachment have continued for decades, and questions and controversies still remain (Kagan, 2011; Sroufe & Siegel, 2011).
One fundamental—and often overlooked—difference between these two perspectives is their respective emphasis on emotional reactivity versus emotion regulation. Whereas proponents of the temperament-based perspective focus on individual differences in infants’ emotional reactions to contextual stimuli, the attachment perspective emphasizes how infants use their caregiver to regulate their emotional arousal (Sroufe, 1985). During the Strange Situation procedure, the standard measure of infant attachment, infants’ responses to a series of separations and reunions with a caregiver are observed. Attachment security is demonstrated by an infant’s capacity to use his or her caregiver to effectively regulate emotional arousal during this moderately stressful task. Although infants’ levels of distress in response to this procedure are highly variable, distress is not a criterion for attachment insecurity. In fact, the two most common patterns of insecure attachment show markedly different distress profiles: During separations from their caregivers, insecure-avoidant infants display minimal distress, whereas insecure-resistant infants are highly upset.
Variability in distress reactivity is also observed among the subtypes of securely attached infants. A portion of securely attached infants display minimal distress (denoted as subcategory B1 or B2), at levels comparable to those of insecure-avoidant infants. 1 Other securely attached infants (denoted as subcategory B3 or B4) are as distressed as insecure-resistant infants (Frodi & Thompson, 1985). 2 The distinguishing feature of securely attached infants is their ability to effectively use their caregiver to regulate their distress, however intense it may be. Thus, securely attached B1 and B2 infants show delight and actively reengage with their caregiver during reunions, whereas insecure-avoidant infants actively avoid interacting with their caregiver. Similarly, securely attached B3 and B4 infants intentionally seek out and are soothed by their caregiver, whereas insecure-resistant infants are inconsolable and show a marked ambivalence toward their caregiver (Ainsworth et al., 1978).
One proposed reconciliation between the temperament- and attachment-based perspectives is that infants’ distress reactivity, but not their attachment security, during the Strange Situation procedure is shaped by biologically based predispositions for negative emotionality (Belsky & Rovine, 1987; Sroufe, 1985). In an initial test of this hypothesis, Belsky and Rovine (1987) grouped infants according to their distress reactivity to the Strange Situation procedure. The high-distress category comprised securely attached B3 and B4 infants and insecure-resistant infants; the low-distress category included securely attached B1 and B2 infants and insecure-avoidant infants. Using this alternative classification system, Belsky and Rovine found that infants’ distress-reactivity grouping was predicted by measures of their early temperament. Compared with low-distress infants, infants in the high-distress category showed less autonomic stability as newborns and were rated by their mothers as being more difficult. However, these measures of early temperament failed to distinguish between securely and insecurely attached infants. This unique association between infant temperament and distress-reactivity grouping underscored the distinction between emotional reactivity and emotion regulation during the Strange Situation procedure and implied that there are unique pathways to each of these behavioral responses.
Subsequent attempts to empirically evaluate this hypothesis have yielded mixed results. Whereas a number of investigations have found unique, positive associations between early measures of infants’ temperament and infants’ distress reactivity during the Strange Situation procedure, these associations have not been uniformly found (Vaughn, Bost, & van IJzendoorn, 2008). One likely explanation for this lack of consistency is the complexity of measuring infants’ biologically based emotional tendencies using traditional approaches (Kochanska & Coy, 2002; Marshall & Fox, 2005; Rothbart & Bates, 2006). Moreover, the nature and timing of infant temperament assessments have varied across studies (Vaughn et al., 2008). Because of these methodological issues, it is challenging to determine on the basis of the extant empirical evidence whether infants’ distress reactivity, but not their attachment security, during the Strange Situation procedure is shaped by their temperamental predispositions.
Molecular genetic analyses afford a more direct and objective means of examining potential biological contributors to infants’ distress reactivity. In particular, a common variation (serotonin-transporter-linked polymorphic region, or 5-HTTLPR) in the serotonin-transporter gene (SLC6A4) has been linked to individual differences in negative emotionality (Caspi, Hariri, Holmes, Uher, & Moffitt, 2010). The 5-HTTLPR polymorphism is characterized by a 44-base-pair insertion/deletion in the promoter region of the gene (Lesch et al., 1996). The short (S) allele is associated with diminished gene transcription, reduced transporter levels, reduced serotonin uptake, and increased activation of the amygdala during the processing of emotional stimuli (Caspi et al., 2010; Hariri et al., 2002; Lesch et al., 1996). At the behavioral level, the S allele is also associated with a heightened risk for negative affect and emotional disorders in adults (Clarke, Flint, Attwood, & Munafo, 2010; Sen, Burmeister, & Ghosh, 2004) and with a heightened risk for negative emotionality in infants (Auerbach et al., 1999). Recent investigations have identified an additional single-nucleotide polymorphism, rs25531, in the long (L) allele. This additional variant (denoted as LG) results in gene-expression levels that are more similar to those of the S allele than to those of noncarriers (denoted as LA; Hu et al., 2005). Examination of the triallelic (S, LG, LA) genotype may provide the most accurate picture of the consequences of 5-HTTLPR variation.
The purpose of the longitudinal study reported here was to examine genetic and caregiving-based contributions to individual differences in infants’ attachment classifications. We predicted that caregiving quality and infants’ genetic variation would have unique associations with attachment classifications. Specifically, we hypothesized that infants’ attachment security would be predicted by their history of maternal responsiveness. In addition, we expected that infants’ 5-HTTLPR variation would predict their subtype of security or insecurity, such that the S allele (or LG equivalent) would be associated with attachment classifications marked by relatively high distress reactivity during the Strange Situation procedure. Potential gene-by-environment interactions were also examined in light of recent interest in the interplay between caregiving quality and infants’ 5-HTTLPR variation (Barry, Kochanska, & Philibert, 2008; Spangler, Johann, Ronai, & Zimmermann, 2009). Because the available evidence for infant attachment and gene-by-environment interactions is mixed (e.g., Luijk et al., 2011), these analyses were considered exploratory.
Method
Participants
Participants were a subsample of 155 infants (74 males, 81 females) and their primiparous mothers, drawn from a larger longitudinal study (N = 267; Sroufe, Egeland, Carlson, & Collins, 2005). This subsample comprised individuals for whom genetic information was available and did not systematically differ from the original sample with respect to sociodemographic risk or focal variables (for details about and results from attrition analyses, see the Supplemental Material available online). At the time of recruitment (between 1975 and 1977), all mothers were below the poverty line and receiving public assistance through the Minneapolis Heath Department. At the time of delivery, the mothers ranged in age from 15 to 34 years (M = 20.8 years, SD = 3.8), and 63% were single. Sixty-seven percent of the infants were Caucasian, 20% were multiracial, 9% were African American, 3% were Native American, and less than 1% were Hispanic or Asian American.
Measures
Maternal responsiveness
When the infants were 6 months old, interactions between mothers and infants were observed in their homes on two different days. Interactions consisted of two 30-min feeding situations and one 20-min play situation. During each of these interactions, maternal behaviors were rated using Ainsworth’s global scales of maternal sensitivity and cooperation (Ainsworth et al., 1978). The Sensitivity vs. Insensitivity Scale assessed the mothers’ ability to perceive and accurately interpret their infants’ signals, as well as to respond to these signals appropriately and promptly. A high score indicates keen attunement to the infant’s cues and timely, contingent responsiveness; a low score indicates a lack of awareness of or disregard for the infant’s signals. The Cooperation vs. Interference Scale assessed the extent to which mothers synchronized their behaviors with their infants’ current mood and interests. Mothers with high scores respected their infant’s autonomy by coordinating their own desires and responsibilities with the infant’s activities. Mothers with low scores behaved in ways that severely or frequently interfered with the infant’s activities or interests.
For both scales, agreement among independent coders was calculated using the Lawlis-Lu index of agreement (Tinsley & Weiss, 1975), with agreement defined as a discrepancy of 2 points or less for each 9-point scale. There was significant agreement for all ratings (p < .05), with t values ranging from 0.66 to 0.87, which indicated that interrater agreement for both scales was moderate to high. The measures of sensitivity and cooperation during the feeding and play interactions were averaged to form a composite measure of maternal responsiveness (α = .93, M = 5.67, SD = 1.47). Data for maternal responsiveness were not available for 6 mother-infant pairs.
Infants’ attachment classifications
Infants’ attachment behaviors were assessed twice, when infants were 12 and 18 months old, using Ainsworth’s Strange Situation procedure (Ainsworth et al., 1978). During this laboratory procedure, infants’ responses to a series of mildly stressful separations and reunions with their mothers were observed. The traditional system was used to classify infants as securely attached (B), insecure-avoidant (A), or insecure-resistant (C). Although the attachment disorganization/disorientation classification has more recently been added to Ainsworth’s original classification system, attachment security and disorganization/disorientation represent theoretically and empirically distinct infant attachment outcomes (see Luijk et al., 2011). Because our hypotheses concerned types of attachment security or insecurity, we did not include the disorganization/disorientation classification in this study.
The attachment assessments at 12 and 18 months were coded by independent teams, and interrater agreement was high for both assessments (89% and 93%, respectively). Any discrepancies in coding were resolved though conferencing. At the 12-month assessment, there were 88 securely attached infants, 36 insecure-avoidant infants, and 23 insecure-resistant infants. At the 18-month assessment, there were 88 securely attached infants, 32 insecure-avoidant infants, and 20 insecure-resistant infants. Attachment data were unavailable for 8 infants at 12 months and for 15 infants at 18 months. There was significant stability in attachment security from 12 to 18 months (κ = .34 p < .001). Continuity in attachment security was not significantly related to distress-reactivity grouping at either 12 or 18 months.
Infants were also categorized according to their level of emotional distress in response to the Strange Situation procedure, using a previously established procedure (e.g., Belsky & Rovine, 1987). Securely attached B1 and B2 infants (n = 50 at 12 months, n = 47 at 18 months) were grouped with insecure-avoidant infants to form a low-distress category. Securely attached B3 and B4 infants (n = 38 at 12 months, n = 41 at 18 months) were grouped with insecure-resistant infants to form a high-distress category. There was significant stability in distress-reactivity groupings from 12 to 18 months (κ = .18, p = .04). Continuity in distress reactivity was not significantly related to attachment security at either 12 or 18 months.
5-HTTLPR variation
In a follow-up assessment when the infant participants were 32 years old, 156 participants provided buccal cells for DNA analysis. Using the conventional method, we extracted DNA with the Epicentre (www.epibio .com) BuccalAmp DNA Extraction Kit to prepare DNA for polymerase-chain-reaction amplification. Genotyping was conducted following previously published protocols (Cicchetti, Rogosch, & Toth, 2011). All samples were genotyped in duplicate for quality control. We were unable to genotype genetic information from 1 participant. There were 43 participants with an LA/LA genotype, 12 participants with an LA/LG genotype, 1 participant with an LG/LG genotype, 61 participants with an S/LA genotype, 7 participants with an S/LG genotype, and 31 participants with an S/S genotype. The 5-HTTLPR triallelic genotype was in the Hardy-Weinberg equilibrium, χ2(3, N = 155) = 1.47, p = .39. Because the LG and S alleles are functionally equivalent in genetic expression (Hu et al., 2005), they were grouped together as S′ alleles. The LA allele was designated as L′. Categorized in this way, 43 participants (27.7%) were in the L′/L′ group, 73 participants (47.1%) were in the S′/L′ group, and 39 participants (25.2%) were in the S′/S′ group.
Missing data
Observations of maternal responsiveness and infants’ attachment classifications were not available for all individuals who provided DNA information. These participants with missing data did not significantly differ from other participants with regard to demographic variables, so their data were assumed to be missing at random (Schafer & Graham, 2002). Following best practices, we carried out multiple imputation for missing data using Bayesian estimation (Schafer & Graham, 2002). Data were imputed 10 times, with results of analyses of the 10 data sets combined according to Rubin’s rules (Rubin, 1987). One participant’s maternal-responsiveness score was an extreme outlier (more than 3 SD below the mean) and was removed prior to analyses (final N = 154).
Analytic approach
The primary analytic approach involved estimating a series of logistic regression models in which measures of infants’ attachment security and distress reactivity during the Strange Situation procedure were separately regressed on ratings of maternal responsiveness from the interactions when the infants were 6 months old, infants’ 5-HTTLPR genotypes (number of S′ alleles), and their statistical interaction. Separate logistic regression analyses were conducted for each time period (i.e., 12 months and 18 months). Sex and ethnicity were included as controls in all models to correct for potential confounds due to population stratification and sex differences. With significance levels of .05 and power of .80, we were able to detect caregiving effect sizes of approximately 1.25, genetic effect sizes of approximately 2.0, and Caregiving × Genetic interaction effect sizes of approximately 1.75, using QUANTO software (Version 1.2.4; hydra.usc.edu/GxE).
Results
Before examining genetic and environmental contributions to infants’ attachment classifications, we evaluated the presence of a correlation between maternal responsiveness and infants’ 5-HTTLPR genotype. Maternal responsiveness did not differ across infants’ genotypes (β = −0.08, p > .10).
Predicting infants’ attachment security
We first conducted analyses predicting infants’ attachment security at 12 months. Greater maternal responsiveness when the infants were 6 months old predicted a greater likelihood of secure attachment at 12 months (OR = 1.42, p < .01; see Table 1). Neither infants’ 5-HTTLPR variation nor the interaction between 5-HTTLPR and maternal responsiveness were significant predictors of infants’ attachment security at 12 months. Further tests evaluated whether maternal responsiveness predicted attachment security for both the high-distress and low-distress reactivity groups. When only high-distress (insecure-resistant and securely attached B3 and B4) infants were considered, maternal responsiveness did not significantly differentiate between securely attached and insecure-resistant infants (OR = 1.27, p > .10). However, when only low-distress (insecure-avoidant and securely attached B1 and B2) infants were considered, greater maternal responsiveness significantly predicted a greater likelihood of secure attachment (OR = 1.54, p = .01).
Results From Regression Analyses Predicting Infants’ Attachment Security During the Strange Situation Procedure at 12 and 18 Months
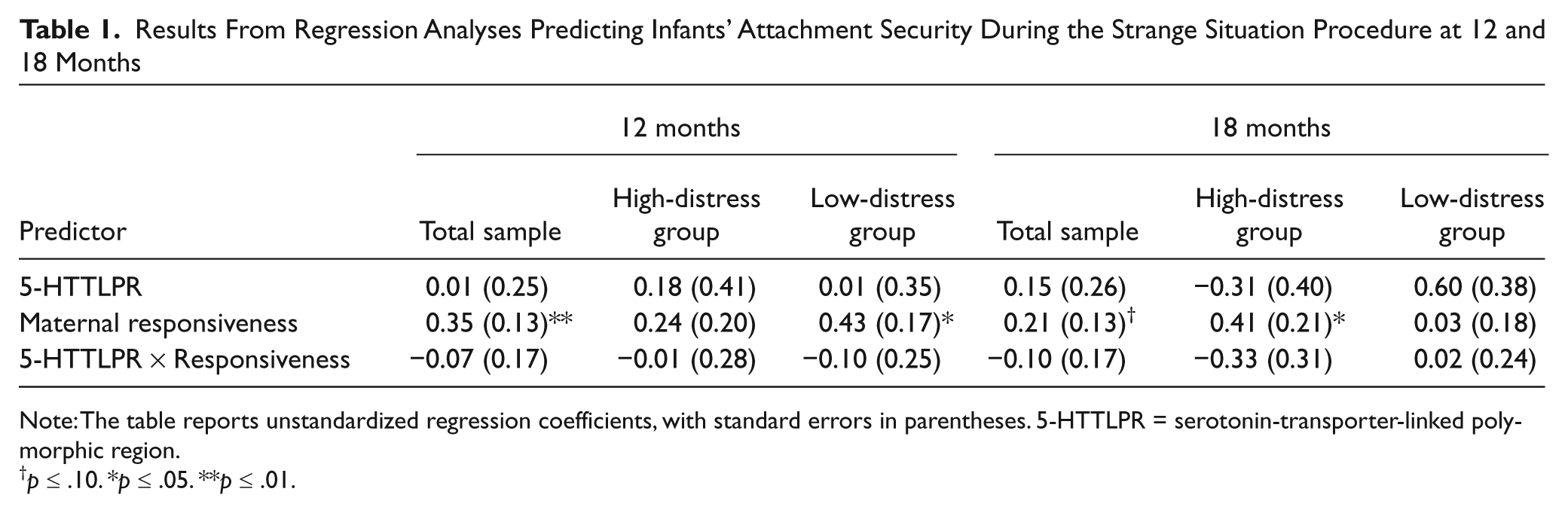
Note: The table reports unstandardized regression coefficients, with standard errors in parentheses. 5-HTTLPR = serotonin-transporter-linked polymorphic region.
p ≤ .10. *p ≤ .05. **p ≤ .01.
Maternal responsiveness was a marginally significant predictor of attachment security at 18 months (OR = 1.23, p = .09; see Table 1). Neither infants’ 5-HTTLPR variation nor the interaction between 5-HTTLPR and maternal responsiveness were significant predictors of infants’ attachment security. When subtypes were considered separately, maternal responsiveness significantly predicted attachment security for high-distress (insecure-resistant and securely attached B3, and B4) infants (OR = 1.50, p = .05) but not for low-distress (insecure-avoidant and securely attached B1, and B2) infants (OR = 1.03, p > .10).
Predicting infants’ distress reactivity
The second set of analyses predicted infants’ distress reactivity during the Strange Situation procedure (see Table 2). Analyses of the 12-month reactivity groupings revealed that infants’ 5-HTTLPR variation was the only significant predictor of infants’ distress reactivity (OR = 2.04, p < .01). Each S′ allele was associated with a twofold increase in the probability of high distress reactivity during the Strange Situation at 12 months. Neither maternal responsiveness nor the interaction between 5-HTTLPR and maternal responsiveness were significant predictors of infants’ distress reactivity. Further tests evaluated whether this association between 5-HTTLPR and distress reactivity applied to both securely and insecurely attached infants. Among securely attached infants, 5-HTTLPR variation continued to predict high-distress classifications of B3 and B4 (OR = 2.13, p = .02). Each S′ allele was associated with approximately a twofold increase in the likelihood of high distress reactivity. Analyses for insecurely attached infants at 12 months were completed next. Because of the low number of insecurely attached infants with two S′ alleles at 12 months, insecurely attached infants with at least one S′ allele (S′/S′ and S′/L′ infants) were considered together for these analyses. Among insecurely attached infants, an S′ allele predicted the insecure-resistant classification (OR = 4.01, p = .05). The odds ratio indicated that an S′ allele predicted a fourfold increase in the probability of being classified as insecure-resistant, rather than insecure-avoidant, at 12 months.
Results From Regression Analyses Predicting Infants’ Distress Reactivity During the Strange Situation Procedure at 12 and 18 Months

Note: The table reports unstandardized regression coefficients, with standard errors in parentheses. 5-HTTLPR = serotonin-transporter-linked polymorphic region.
p ≤ .10. *p ≤ .05. **p ≤ .01.
When attachment classifications at 18 months were examined, infants’ distress reactivity was not significantly associated with 5-HTTLPR variation (OR = 1.21, p > .10). Neither maternal responsiveness nor the interaction between 5-HTTLPR variation and maternal responsiveness were significant predictors of infants’ distress reactivity at 18 months. However, follow-up tests indicated that 5-HTTLPR variation marginally predicted reactivity among insecurely attached infants (OR = 1.99, p = .08). This association was considered significant because it was the result of a two-tailed test of our directional hypothesis. Moreover, the odds ratio indicated that each S′ allele was associated with approximately a twofold increase in the likelihood of being classified as insecure-resistant, rather than insecure-avoidant, during the Strange Situation procedure. Infants’ 5-HTTLPR genotype was not associated with distress reactivity for securely attached infants (OR = 0.80).
Discussion
The results of this longitudinal study are consistent with our hypotheses about genetic and caregiving-based contributions to infants’ attachment classifications. When infants were 12 months old, their 5-HTTLPR variation predicted how distressed they became during the moderately stressful Strange Situation procedure, and infants’ history of interactions with their caregiver predicted how effectively they used their caregiver to regulate their distress. These associations were generally replicated when infants were 18 months old, although genetic influences on distress reactivity were observed only for insecurely attached infants. These findings underscore the distinction between emotional reactivity and emotion regulation during the Strange Situation procedure and provide evidence for unique developmental pathways to each of these behavioral responses in early infancy.
These findings support attachment theory’s central hypothesis that infants’ attachment security reflects the history of their interactions with their caregiver (Ainsworth et al., 1978; Bowlby, 1982; Sroufe, 1985). Infants who experience sensitive and cooperative care are hypothesized to form perceptions of their caregiver as available and responsive to their needs, which enables them to use their caregiver for effective emotion regulation. Our findings provide additional support for this idea, given that infants’ attachment security at both 12 and 18 months was predicted by their history of maternal responsiveness. Although the associations for specific subgroups were not always statistically significant, these results may have reflected Type II errors resulting from small sample sizes. Furthermore, the observed effects were consistently in a positive direction. Numerous observational and experimental investigations have provided additional support for this hypothesis (Cicchetti et al., 2011; de Wolff & van IJzendoorn, 1997; NICHD Early Child Care Research Network, 1997). Moreover, several behavioral genetics investigations have found large shared-environmental contributions and trivial genetic contributions to infants’ attachment security (Bakermans-Kranenburg, van IJzendoorn, Bokhorst, & Schuengel, 2004; Fearon et al., 2006; Roisman & Fraley, 2008).
Although 5-HTTLPR variation was unrelated to infant attachment security, it did predict infants’ subtypes of security or insecurity. More specifically, the less efficient 5-HTTLPR S′ allele was associated with attachment classifications characterized by high distress during the Strange Situation procedure. Securely attached 12-month-old infants who became highly distressed but effectively used their caregiver for comfort (subtypes B3 and B4) carried the 5-HTTLPR S′ allele more often than did securely attached infants who showed minimal distress and active enjoyment of their caregiver (subtypes B1 and B2). Infants’ 5-HTTLPR variation also predicted their subtypes of insecure attachment at both 12 and 18 months. Infants who became inconsolably distressed during the Strange Situation (i.e., insecure-resistant infants) carried the 5-HTTLPR S′ allele more often than did infants who displayed minimal distress and avoided interactions with their caregiver (i.e., insecure-avoidant infants). Although these findings appear to contradict the hypothesis that the insecure-avoidant and insecure-resistant classifications reflect specific histories of interactions between infants and their caregivers, more fine-grained measures of specific types of low maternal responsiveness (e.g., rejection and inconsistency) are needed to thoroughly evaluate this hypothesis. These results do indicate that infants’ 5-HTTLPR variation biases insecure attachment in the direction of either avoidance or resistance by contributing to differences in emotional reactivity inherent to these two patterns of attachment insecurity (Belsky & Rovine, 1987).
There was limited evidence for interactions between genetic and environmental influences in our study. First, infants’ 5-HTTLPR variation was not associated with the quality of their mother’s responsiveness (Fearon et al., 2006; Roisman & Fraley, 2008). Second, maternal responsiveness and infants’ 5-HTTLPR variation did not have an interaction effect in predicting either infants’ attachment security or their distress reactivity. Although genetic variation and caregiving quality both played a role in shaping infants’ attachment behavior, they did so independently of one another. The lack of an interaction between genetic and environmental effects on attachment security further underscores the centrality of the parent-child relationship for infant attachment security (e.g., Luijk et al., 2011). However, studies with nonhuman primates and with older children have demonstrated that an unsupportive caregiving context may exacerbate 5-HTTLPR’s influence on negative affect (Fox et al., 2005; Suomi, 2004). In our sample, the finding that genotype influenced emotional reactivity among securely attached infants at 12 months, but not 18 months, may suggest that genetic influences on distress reactivity are dependent on the quality of the infant-caregiver relationship as well as the age of the child (Sroufe, 1985).
Although a handful of previous studies have investigated molecular genetic contributions to infant attachment (Barry et al., 2008; Cicchetti et al., 2011; Luijk et al., 2011; Spangler et al., 2009), this study represents an innovative approach to the issue by emphasizing individual differences in distress reactivity that are inherent in subtypes of attachment security and insecurity. The use of attachment subtypes may assist in clarifying the mixed evidence regarding the association between infants’ 5-HTTLPR variation and attachment security (Luijk et al., 2011). One important task for future research will be to replicate these findings using more direct measures of infants’ distress during the Strange Situation. The current findings extend those of prior investigations of infant attachment and temperament (Marshall & Fox, 2005; Vaughn et al., 2008), and also extend previous behavioral genetics findings of differential heritability estimates for infant-caregiver relationship quality and infant temperament (Bakermans-Kranenburg et al., 2004; Roisman & Fraley, 2006). Finally, our study also contributes to the growing evidence for the association between 5-HTTLPR and negative emotionality in early infancy (Auerbach et al., 1999; Caspi et al., 2010).
These findings indicate that caregiving context and infants’ genetic variation each make unique contributions to individual differences in infants’ attachment behavior. A history of responsive caregiving influenced whether or not the infants’ attachment relationships became secure or insecure, and the infants’ 5-HTTLPR variation shaped how this security or insecurity was manifested. A fruitful avenue for future investigations of infant attachment would be to combine measures of caregiving quality with genetic variables, not because either caregiving or genetics has a privileged status or might independently provide a superior explanation, but because combining measures would yield a more comprehensive understanding of the nature and origins of individual differences in infant attachment.
Footnotes
Acknowledgements
We thank Alan Sroufe for his comments on an earlier version of this manuscript.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by National Institute of Mental Health Grant R01MH40864-09 to Byron Egeland and by National Institute of Mental Health Predoctoral Training Grant T32MH015755-33 to K. Lee Raby.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.