
Abstract
The merging of psychological and genetic methodologies has led to an increasing appreciation of environmental moderators of the relationships between genotype and phenotype. Here we used a nonhuman-primate model to study the moderating effect of the mother’s genotype on the association of a dopamine D4 receptor (DRD4) gene polymorphism with juvenile impulsivity, assessed in a standardized social-challenge test. The results showed that juvenile carriers of the rare 5-repeat variant of the exon III 48-base-pair repeat polymorphism scored significantly higher in social impulsivity than juveniles homozygous for the common 6-repeat allele. In addition, juvenile genotype interacted with maternal genotype to influence impulsivity, with the highest rates of impulsivity found in variant offspring with variant mothers. These results highlight the importance of considering the genotype of the parents in studies of early experience and vulnerability genes for impulsivity-related traits.
A core concern in psychology has been to identify the source of stable individual differences in personality and behavior (Penke, Denissen, & Miller, 2007). Impulsivity is a trait that has been shown to be influenced by both environmental and genetic factors (Bezdjian, Baker, & Tuvblad, 2011). One of the more robust findings in behavior genetics research is the association of a 48-base-pair variable number of tandem repeats (VNTR) in exon III of the dopamine D4 receptor (DRD4) gene with attention-deficit/hyperactivity disorder (ADHD; Gizer, Ficks, & Waldman, 2009; Tharpar, Langley, Owen, & O’Donovan, 2007). The DRD4 polymorphism has also been associated with continuous measures of impulsivity-related traits in some studies but not in others (Bidwell et al., 2011; Munafò, Yalcin, Willis-Owen, & Flint, 2008). Part of this inconsistency may be explained by variation in the developmental environment, as several recent studies of child behavior have reported significant interactions of the DRD4 7-repeat VNTR with early family environments in predicting child externalizing behavior (Bakermans-Kranenberg & van IJzendoorn, 2006; Sheese, Voelker, Rothbart, & Posner, 2007).
Many studies of interactions among family influences, genotype, and child outcomes assess maternal behavior in standardized interactions with their children and measure child behavior using parent-report questionnaires (Bakermans-Kranenberg & van IJzendoorn, 2011). An underappreciated feature of the developmental environment for children with psychiatric-risk genotypes is that they are highly likely to have one or both caretaking parents with the risk genotype. This is particularly significant for the DRD4 gene because the 7-repeat VNTR in the mother has been associated with variation in maternal sensitivity under difficult conditions (Kaitz et al., 2010; van IJzendoorn, Bakermans-Kranenburg, & Mesman, 2008). This circumstance makes it difficult to disentangle the influences of parental genotype and parental environment on the assessment of infant and child behavior.
Nonhuman primate models have contributed significantly to our understanding of interactions between early environments and risk genotypes in the development of biobehavioral traits (Suomi, 2006). In the study reported here, we used a nonhuman-primate model to test the effects of maternal and offspring DRD4 genotype on a trait related to ADHD. Like humans, vervet monkeys have a variable-number 48-base-pair repeat in exon III of the DRD4 gene, and this polymorphism has been associated with novelty-seeking behavior. Monkeys with one copy of the rare 5-repeat variant had significantly shorter latencies to approach a novel object compared with individuals homozygous for the common 6-repeat allele (Bailey, Breidenthal, Jorgensen, McCracken, & Fairbanks, 2007).
There are several advantages of the vervet model for assessing influences of parent and offspring genotype on behavioral traits related to impulsivity and ADHD. In the vervet social system, mothers are the primary caregivers, and fathers are not involved. Thus, this system provides the opportunity to differentiate the effects of offspring genotype from possible effects of caregiving parent genotype and Offspring Genotype × Parent Genotype interactions more readily than in two-parent human families. In addition, we developed a standardized procedure, the Intruder Challenge Test, to assess impulsivity in a social situation that does not rely on parental report or parental involvement during the test (Fairbanks, 2001). The Social Impulsivity Index derived from this test has been shown to be a reliable and heritable index of individual differences in impulsivity (Fairbanks & Jorgensen, 2011; Fairbanks et al., 2004).
We measured social impulsivity in juvenile vervets using the Intruder Challenge Test to assess two hypotheses. First, we proposed that juvenile vervets with the 5-repeat variant of the DRD4 VNTR would have higher levels of impulsivity in the Intruder Challenge Test compared with juveniles homozygous for the common 6-repeat allele, and second, we hypothesized that the effect of the variant genotype on juvenile impulsivity would be moderated by the mother’s DRD4 genotype.
Method
Subjects
Subjects were 357 juvenile vervet monkeys (Chlorocebus aethiops sabaeus) born between 1999 and 2005 at the University of California, Los Angeles and Department of Veterans Affairs Vervet Research Colony. The sample consisted of 200 males and 157 females with a mean age of 2.5 years (SD = 0.4) at the time of testing. All subjects were raised by their mothers in matrilineal social groups managed to reflect the natural social composition of vervet groups in the wild. Subjects and their mothers were part of an eight-generation pedigree descended from 57 original founders captured in St. Kitts, West Indies. Juvenile subjects were offspring of 169 mothers (mean subjects per mother = 2.1, SD = 1.3). Mothers ranged in age from 3 to 23 years (M = 9.0, SD = 4.0) at the time of the subjects’ birth.
Social groups were housed in 16 large outdoor enclosures with adjacent indoor shelters. Each enclosure had chain-link siding that provided visual access to the outside, with one or two large sitting platforms and numerous shelves, climbing structures, and enrichments devices.
Procedures
Genotyping
DNA was extracted from whole blood using standard human DNA extraction kits and the manufacturer’s extraction protocol (Qiagen, Valencia, California). The 48-base-pair VNTR in exon III of the DRD4 gene was genotyped using the same oligonucleotide primers used for the human DRD4 exon III 48-base-pair VNTR (Kustanovich et al., 2004). Alleles were determined in comparison with molecular weight standards and human controls with previously determined genotypes in the VNTR. Genotyping of the entire colony revealed two variants of the DRD4 VNTR: a common 6-repeat and a rare 5-repeat variant (Bailey et al., 2007). Distribution of the two DRD4 alleles in the Vervet Research Colony population does not deviate significantly from Hardy-Weinberg equilibrium, χ2(1, N = 1,351) = 0.36.
In this sample of juvenile subjects, 21 were heterozygous with one 5-repeat allele (variant genotype), and 336 were homozygous for the 6-repeat allele (common genotype). Twenty-one juvenile subjects had variant mothers. No subjects or mothers were homozygous for the 5-repeat variant. Breakdown of the sample by subject genotype and mother genotype is as follows: common juvenile–common mother (n = 325), common juvenile–variant mother (n = 11), variant juvenile–common mother (n = 11), and variant juvenile–variant mother (n = 10).
Intruder Challenge Test
All juvenile colony members were tested with the Intruder Challenge protocol when they were between 1.5 and 3.0 years of age, an age range that is developmentally similar to 6- to 12-year-old human children. Before the test, three to five juvenile subjects were retained in the large outdoor portion of their home enclosure while all other group members were confined out of sight in the indoor area. An unfamiliar adult male vervet (the intruder) in an individual cage was placed at the edge of the home enclosure, immediately outside the chain-link fence and within arm’s reach of the subjects. Behavioral responses of each subject—including latency to approach within 1 m, the number of 1-min intervals that subjects were observed standing or sitting within 1 m, touching or nuzzling the intruder or the intruder’s cage, displaying to or threatening the intruder—were coded by trained observers as present or absent in each minute of the 30-min test session (Fairbanks, 2001).
Social Impulsivity Index
The Social Impulsivity Index was derived from factor analysis of behaviors directed toward the intruder. It combines short latency to approach with risky, assertive, and aggressive behaviors to the intruder (Fairbanks, 2001). Information on the index’s internal reliability, test-retest reliability, heritability, and validity is provided in Fairbanks (2001), Fairbanks et al. (2004), and Fairbanks and Jorgensen (2011).
Results
Descriptive statistics
Scores of the juvenile subjects on the Social Impulsivity Index ranged from 0 to 114 (M = 43.1). Juveniles in the most inhibited quartile took 19 min, on average, to approach the intruder for the first time. They averaged only two intervals within 1 m, and most never sat near, touched, nuzzled, displayed to, or threatened the intruder. Juveniles in the most impulsive quartile, in contrast, approached in the first minute, were present within 1 m in 60% of the intervals, and were observed touching, nuzzling, and displaying to the intruder on multiple occasions.
Social-impulsivity scores were influenced by sex and age. Males had significantly higher scores than did females (45.8 vs. 38.3, respectively), t(355) = 3.48, p < .01, and scores increased with age from 1.5 to 3.0 years, r(355) = .30, p < .01. Juveniles with the variant 5-repeat allele did not differ significantly in age or sex from juveniles homozygous for the common 6-repeat allele. Mother’s age, matrilineal dominance rank, and the number of offspring per mother were not significantly related to maternal or juvenile genotype or to juvenile impulsivity scores.
Juvenile behavior by DRD4 genotype
To examine DRD4 effects on behavior, we conducted an analysis of variance with age and sex as covariates and juvenile DRD4 genotype as the grouping variable. Results indicated that juveniles with the 5-repeat variant scored significantly higher on the Social Impulsivity Index than did juveniles homozygous for the common 6-repeat allele (Table 1). Juveniles with the variant genotype were quicker to approach the intruder; they spent more 1-min intervals within 1 m of the intruder; and they had higher rates of touching, nuzzling, and displaying toward the intruder.
Mean Social Impulsivity Index Scores and Component Behaviors by Juvenile Dopamine D4 Receptor Genotype
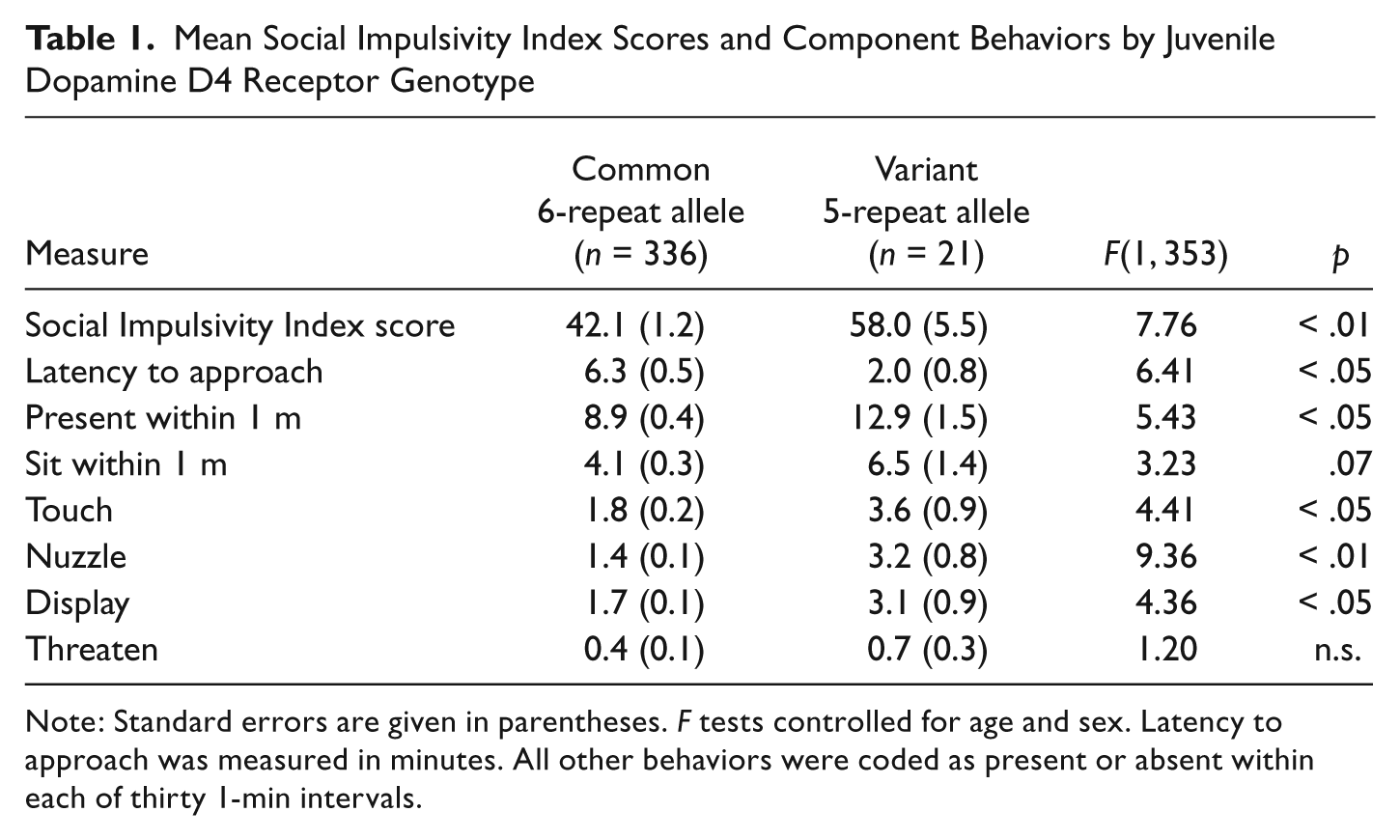
Note: Standard errors are given in parentheses. F tests controlled for age and sex. Latency to approach was measured in minutes. All other behaviors were coded as present or absent within each of thirty 1-min intervals.
Effects of juvenile genotype and maternal genotype on social impulsivity
Analysis of social-impulsivity scores by DRD4 genotype of the juvenile and the mother indicated that the influence of offspring genotype was moderated by maternal genotype (Fig. 1). The analysis of variance results for the full model, with juvenile age and sex as covariates, indicated a main effect of juvenile genotype, F(1, 351) = 12.38, p < .001,

Mean social-impulsivity score as a function of dopamine D4 receptor (DRD4) genotype of the juvenile subjects and their mothers. Subjects and mothers either had the common homozygous 6-repeat allele or the variant 5-repeat allele. Error bars represent standard errors of the mean.
Subjects in this study were all members of an extended pedigree, so to verify that the results were not biased by lack of independence, we repeated the analysis using a pedigree-based variance components approach (Almasy & Blangero, 1998). The variance components model included the kinship matrix with juvenile and maternal DRD4 genotype and their interaction entered as covariates. An analysis that included only main effects confirmed the significant effect of juvenile DRD4 genotype (p = .01) on social-impulsivity scores. Addition of the interaction term to the model revealed a significant interaction of juvenile and maternal genotype (p = .002) on juvenile social impulsivity. The interaction accounted for most of the juvenile genotype effect, which was no longer significant. The heritability of juvenile social impulsivity was h2 = .39 (± .11) after accounting for all of the covariates.
Discussion
The results of this study demonstrate that a polymorphism in the DRD4 gene influences impulsivity in juvenile monkeys. Young monkeys with the rare 5-repeat variant of the DRD4 48-base-pair exon III VNTR were quicker to approach an adult male stranger and more likely to engage in risky and assertive behavior toward it. However, this relationship between impulsivity and the variant genotype was seen only in monkeys whose mothers also carried the variant allele. The relatively small number of variant individuals makes generalization of these results tentative without replication, but they do highlight the importance of considering parental genotype as a component of the developmental environment in studies of genetic and environmental influences on biobehavioral traits.
The current study did not address the mechanisms responsible for the interaction between maternal and offspring genotype. One potential explanation may stem from maternal caregiving style. Nonhuman primates, like humans, have a prolonged period of early dependence on their mothers, and there is considerable evidence that variation in the early mother-infant relationship has effects on behavioral development (Parker & Maestripieri, 2011). Our prior research with vervets demonstrated that highly protective vervet mothers produce juvenile offspring who are less impulsive and more inhibited in their approach toward novel objects and spaces (Fairbanks & McGuire, 1993). Limited observations of early mother-infant interactions among the current subjects indicate that variant mothers with variant offspring had the lowest level of maternal protectiveness and that variant mothers with common offspring had the highest level (Fairbanks, Breidenthal, Bailey, & Jorgensen, 2006). These findings offer a possible explanation for higher impulsivity in the former group and lower impulsivity in the latter group. Further research is needed to determine whether the quality of maternal care is a mediating factor in the development of impulsivity in this study.
In human families, children who are carriers of vulnerability genes for psychiatric disorders will have one or both parents that carry the same vulnerability genes. In the case of the DRD4 VNTR, this means that children with the 7-repeat variant are more likely than children without this variant to have parents with adult ADHD (Lynn et al., 2005). Adult ADHD is known to affect educational attainment, income, and job and marital stability (Brod, Schmitt, Goodwin, Hodgkins, & Niebler, 2011), and ADHD symptoms in fathers add to genetic risk for child hyperactivity/impulsivity (Auerbach et al., 2010). Parents with the 7-repeat VNTR are also more likely than parents without this variant to be impulsive, unfaithful, and promiscuous and have problems with alcohol and substance abuse (Garcia et al., 2010; McGeary, 2009; Reiner & Spangler, 2011). These are all factors that affect the family environment and have the potential of interacting with child characteristics to influence behavioral development.
In addition to potential behavioral mediators of the interaction between parental and offspring genotype, there could be biological mediators. Imprinting is a process in which the allele from one parent is silenced, but evidence indicates that the human DRD4 gene is not subject to imprinting (Cichon, Nöthen, Wolf, & Propping, 1996), which makes it unlikely that imprinting of the vervet DRD4 is an explanation for the interaction reported here. Another potential explanation is genotype-related differences in the prenatal environment, but the nonhuman-primate model controls for many of the factors that could differentially influence gestation. The pregnant mothers in this study ate uniform diets and were not exposed to tobacco, alcohol, or drugs of abuse that can elicit later impulse-control problems in human children (Minnes, Lang, & Singer, 2011).
Clearly, more research is needed to understand the mechanisms by which maternal genotype and offspring genotype interact. There is preliminary evidence that a similar process is occurring in humans (Gadow et al., 2010); therefore, identifying the mechanisms responsible for this effect is likely to clarify the processes by which the DRD4 affects behavior. Increased attention to interactions with parental genotype may help to explain some of the variance in human genetic-association studies. The results presented here demonstrate the value of considering the mother’s genotype in studies of early experience and vulnerability genes for impulsivity-related disorders.
Footnotes
Acknowledgements
We thank Adriana Jacobsen, Karin Blau, Danielle Epstein, Amanda Turner, Clayton Clement, Glenvile Morton, and Daniel Diekmann for assistance with data collection and animal care.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by National Institutes of Health Grants R01-MH61852 and P40-RR019963.