
Abstract
People often conform to others with whom they associate. Surprisingly, however, little is known about the possible hormonal mechanisms that may underlie in-group conformity. Here, we examined whether conformity toward one’s in-group is altered by oxytocin, a neuropeptide often implicated in social behavior. After administration of either oxytocin or a placebo, participants were asked to provide attractiveness ratings of unfamiliar visual stimuli. While viewing each stimulus, participants were shown ratings of that stimulus provided by both in-group and out-group members. Results demonstrated that on trials in which the ratings of the in-group and out-group were incongruent, the ratings of participants given oxytocin conformed to the ratings of their in-group but not of their out-group. Participants given a placebo did not show this in-group bias. These findings indicate that administration of oxytocin can influence subjective preferences, and they support the view that oxytocin’s effects on social behavior are context dependent.
Humans and nonhuman primates alike adjust their behavior and beliefs toward others in their social circles (Cialdini & Goldstein, 2004; Whiten, Horner, & de Waal, 2005). In humans, conformity can affect judgments ranging from low-level perceptual line-length estimates (Asch, 1951) to more complex behaviors, such as energy conservation (Schultz, Nolan, Cialdini, Goldstein, & Griskevicius, 2007) and jury verdicts (Davis, 1973). To explain these well-documented tendencies, cultural-evolutionary models suggest that conformity has adaptive value under a wide range of conditions (Henrich & Boyd, 1998). By conforming to common behaviors and shared opinions of one’s own group or community (i.e., one’s in-group), members benefit from the wisdom of the group as a whole and thus increase survival likelihood at both the personal and the group level.
If in-group conformity indeed has adaptive value, it seems likely that there are evolved biological mechanisms that may facilitate and sustain in-group conformity. In the study reported here, we hypothesized that in-group conformity is mediated by oxytocin, a neuropeptide produced in the hypothalamus that functions as both hormone and neurotransmitter (Bartz, Zaki, Bolger, & Ochsner, 2011; Donaldson & Young, 2008). Numerous studies have demonstrated that oxytocin, well known for its role in reproduction and pair-bond formation (Gainer & Wray, 1994), also plays a pivotal role in motivating affiliation and prosocial behavior. For instance, experiments with animals have found that the manipulation of oxytocin binding in the rat brain modulates caring behavior toward pups (Olazábal & Young, 2006) and that the neural distribution of oxytocin receptors correlates with the intensity of pair bonding in voles (Insel & Shapiro, 1992). In humans, intranasal administration of oxytocin reduces distress, enables the behavioral expression of trust, and appears to motivate cooperation and generosity (Baumgartner, Heinrichs, Vonlanthen, Fischbacher, & Fehr, 2008; Declerck, Boone, & Kiyonari, 2010; Kosfeld, Heinrichs, Zak, Fischbacher, & Fehr, 2005; Zak, Stanton, & Ahmadi, 2007).
Although there is a robust relationship between oxytocin and prosociality, it is becoming evident from the literature that the effect of oxytocin on social behavior is not indiscriminately positive but that situational and individual factors critically moderate the effects of oxytocin on prosocial behavior (Bartz et al., 2011; Kemp & Guastella, 2011). For instance, oxytocin appears to increase trust toward partners only when cues about untrustworthiness are absent (Mikolajczak et al., 2010); it enhances cooperation only when one’s partner is known (Declerck et al., 2010); and it motivates negative feelings, such as envy and schadenfreude, following losses in interpersonal competition (Shamay-Tsoory et al., 2009). Similarly, oxytocin’s effects on trust and cooperation depend on group membership—people given oxytocin as compared with people given a placebo are more cooperative toward in-group members than toward out-group members (De Dreu et al., 2010) and are less willing to sacrifice in-group than out-group members when presented with hypothetical moral-choice dilemmas (De Dreu, Greer, Van Kleef, Shalvi, & Handgraaf, 2011).
This emerging insight that oxytocin plays an important role in group affiliation—and may therefore encourage parochial cooperation and in-group favoritism—raises the possibility that oxytocin may stimulate in-group conformity. However, whether oxytocin’s effects on in-group favoritism are sufficiently powerful to actually influence individuals’ subjective preferences remains an important open question. In the present study, we examined this possibility by assessing whether oxytocin (a) stimulates conformity and (b) induces in-group conformity in particular. We employed a minimal-group-setting paradigm, which utilized a neutral decision domain and allowed each participant’s opinion to be voiced anonymously. By using this basic setting, we were able to test the most fundamental conditions for an in-group conformity effect, as the design eliminated reputation concerns for the participant as well as the possibility of gaining positive public recognition by conforming to the opinions of other group members.
After administration of either oxytocin or a placebo, participants were assigned to arbitrary groups on the basis of trivial criteria. We hypothesized that administering oxytocin would lead to greater in-group conformity than administering the placebo would. Demonstrating that oxytocin influences in-group conformity in particular, even under these most basic conditions, would illustrate that the effects of oxytocin are sufficiently strong to influence actual preference and would provide valuable support for earlier findings suggesting that oxytocin tunes the individual specifically toward his or her in-group.
Method
Design and participants
To examine effects of oxytocin on in-group conformity, we recruited 74 males for a double-blind, randomized, placebo-controlled between-subjects design. We administered oxytocin to 37 participants and a placebo to the other 37 participants. To avoid pharmacological effects other than those caused by oxytocin, we used a placebo that contained all active ingredients except for the neuropeptide. Participants received €10 ($13) and provided informed consent before the experiment. Exclusion criteria were significant medical or psychiatric illness, medication, smoking more than five cigarettes per day, and drug or alcohol abuse. Participants were instructed to refrain from smoking or drinking (except for water) for 2 hr before the experiment and were tested between noon and 4 p.m. The experiment was approved by the University of Amsterdam ethics committee and complied with American Psychological Association guidelines.
Five participants (3 in the oxytocin condition and 2 in the placebo condition) were excluded because they failed to follow instructions correctly (> 10 trials with response times < 1 s); therefore, analyses were conducted on 69 participants (mean age = 21.1 years, SD = 2.9 years). Exclusion of the 5 participants did not affect the results.
Experimental procedure
Each experimental session tested 6 participants at a time, with participants seated in individual cubicles to prevent any form of communication. They self-administered the medication (Syntocinon spray, Novartis, Basel, Switzerland; three puffs per nostril, each with 4 International Units of oxytocin or the placebo) under experimenter supervision. The experimenter then left the cubicle, and participants completed a series of unrelated tests. The test instructions guaranteed complete anonymity for the participants.
Effects of oxytocin typically emerge after 30 to 40 min (Baumgartner et al., 2008; De Dreu et al., 2010; De Dreu et al., 2011; Kosfeld et al., 2005); therefore, after 38 to 42 min (depending on how quickly participants completed the unrelated tasks), the computer switched to the main experiment. First, participants learned that they would perform a task involving their own group (denoted as “Team Y6”) and another three-person group (denoted as “Team X4”; labeling was counterbalanced, had no effects, and was therefore not considered further). Group assignment was based on the order in which participants signed up for the experiment, and participants were told that most, though not necessarily all, group members were currently present in the laboratory (De Dreu et al., 2010). Finally, they were told that they would not know who was in either group and that responses would remain anonymous.
Then, participants were told that they would be asked to rate a series of symbols, using a scale ranging from 1 (not attractive at all) to 11 (very attractive). 1 The task was self-paced. While viewing the symbols, participants also saw the ratings for each symbol from the members of their own team as well as from the members of the other team, if available at that time. On some trials, no ratings were provided; on other trials, either one or two ratings were given so we could examine whether the number of ratings affected the strength of the conformity bias and to enhance the impression that ratings were in real time (see Fig. 1 for a sample trial). Trials of interest were those in which in-group or out-group members either liked (high ratings of 8–11) or disliked (low ratings of 1–4) the symbol. Trials with intermediate ratings (from 5–7) were used as filler trials to prevent participants from guessing the study aim, and trials in which participants received no rating information (blank trials) were used to assess whether oxytocin affected general attractiveness judgments. Following this task, participants completed the Positive and Negative Affect Schedule (Watson, Clark, & Tellegen, 1988) to assess potential mood effects of oxytocin.
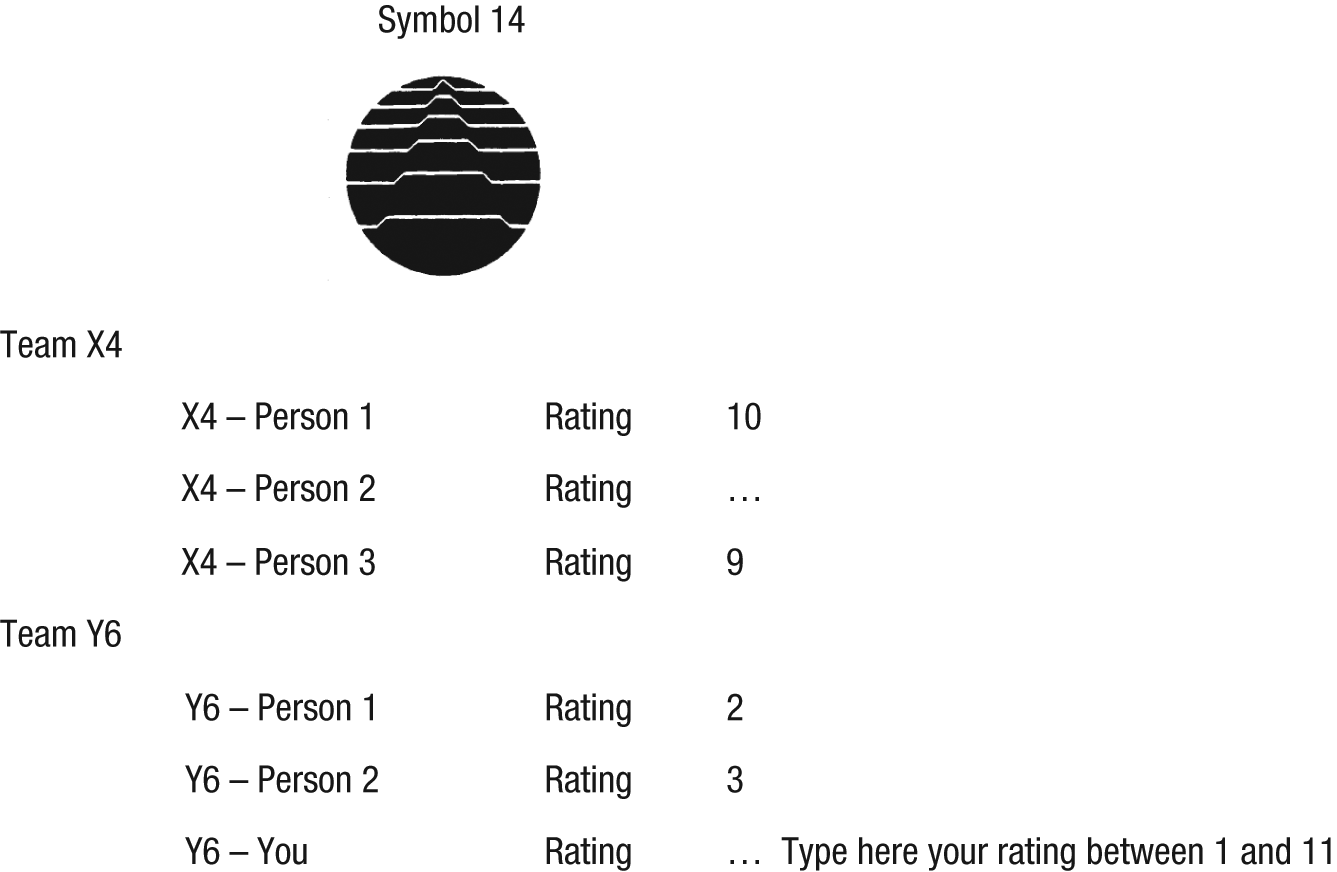
Sample trial. On each trial, participants saw a symbol and ratings of that symbol’s attractiveness by out-group members and in-group members. Participants were then asked to provide their own rating. Ratings were made on a scale ranging from 1, not attractive at all, to 11, very attractive, and participants were shown one, two, or no ratings from each team, depending on the trial. In this example, two out-group members (Team X4) rated the symbol as being attractive, and two in-group members (Team Y6) rated the symbol as being unattractive.
Results
To determine whether participants showed a general conformity effect, we first analyzed the congruent trials in which two in-group and two out-group members both liked or both disliked the symbol. This 2 (treatment: placebo vs. oxytocin) × 2 (rating: in-group high, out-group high vs. in-group low, out-group low) mixed-model analysis of variance, with treatment as a between-subjects factor and rating as a within-subjects factor, showed a main effect of rating, F(1, 67) = 16.44, p < .001, but no interaction between treatment and rating, F(1, 67) = 1.79, p = .185. This finding demonstrated that participants gave significantly higher ratings when both in-group and out-group members liked the symbol (M = 6.51, SD = 1.36) and lower ratings when both in-group and out-group members disliked the symbol (M = 5.51, SD = 1.65), thus confirming the presence of a general tendency to conform to other peoples’ judgments (Cialdini & Goldstein, 2004).
To test our hypothesis that oxytocin affected in-group conformity in particular, we analyzed the incongruent trials—that is, where the in-group and the out-group gave opposite ratings (i.e., when the in-group liked the symbols but the out-group did not and vice versa). These trials included those in which two members of each group gave ratings, as well as trials in which ratings were given by just one in-group member and one out-group member. A 2 (treatment: placebo vs. oxytocin) × 2 (rating: in-group high, out-group low vs. in-group low, out-group high) × 2 (number of members: 1 in-group, 1 out-group vs. 2 in-group, 2 out-group) mixed-model analysis of variance, with treatment as a between-subjects factor and rating and number of members as within-subjects factors, revealed a significant interaction only between rating and treatment, F(1, 67) = 5.76, p = .019. As Figure 2 shows, oxytocin stimulated conformity when in-group and out-group members gave contrasting ratings, independent of the number of ratings provided. Specifically, participants given oxytocin expressed preferences that were closer to those of the in-group, t(33) = 2.50, p = .018 (in-group low, out-group high: M = 5.44, SD = 1.27; in-group high, out-group low: M = 6.25, SD = 1.4), but participants given placebo did not, t(34) = 0.93, p = .360 (in-group low, out-group high: M = 6.00, SD = 1.47; in-group high, out-group low: M = 5.69, SD = 1.42). This supports the hypothesis that oxytocin stimulates in-group conformity.
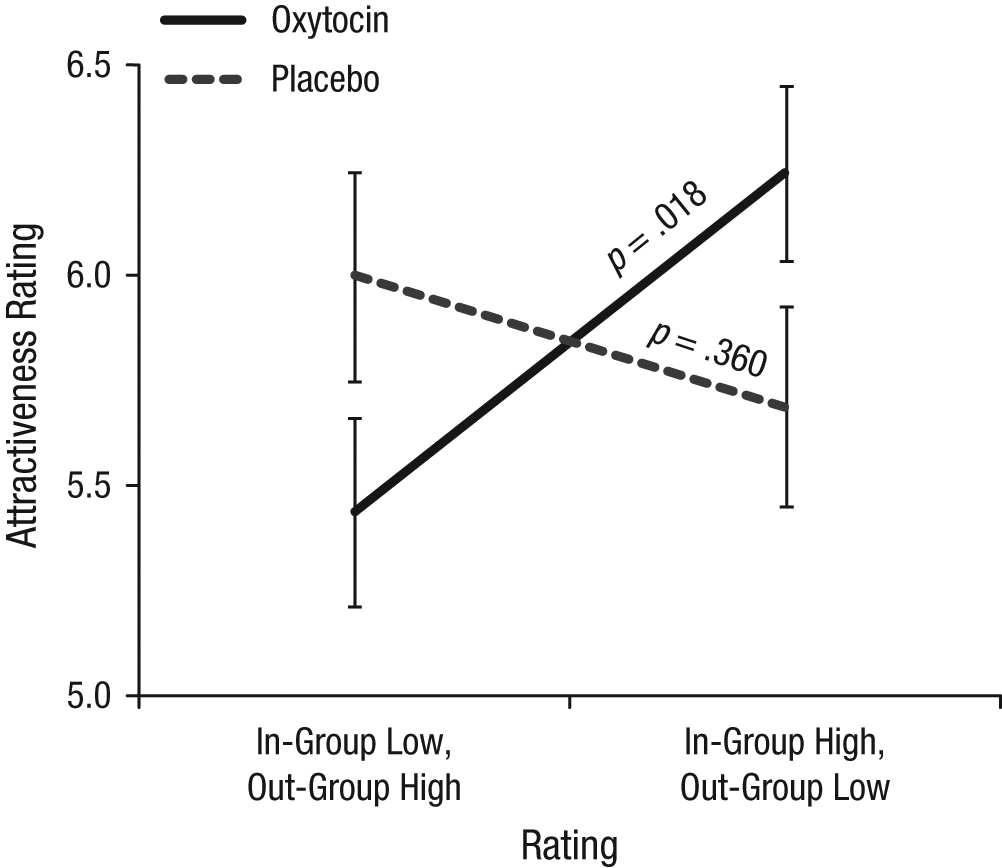
Mean rating of the attractiveness of symbols as a function of groups’ ratings and participants’ condition. Results are shown for incongruent trials, that is, when the in-group and the out-group gave opposite ratings. Error bars indicate standard errors of the mean.
Analysis of ratings on trials in which no ratings were provided showed no effect of oxytocin, t(67) = 0.78, p = .437 (oxytocin: M = 5.37, SD = 1.91; placebo: M = 5.75, SD = 1.83), which indicates that oxytocin did not result in a particular response bias. Furthermore, participants’ responses on the Positive and Negative Affect Schedule showed no effect of oxytocin on either positive affect, F(1, 67) = 0.25, p = .83 (oxytocin: M = 3.3, SD = 1.27; placebo: M = 3.22, SD = 1.22), or negative affect, F(1, 67) = 0.33, p = .80 (oxytocin: M = 2.41, SD = 1.18; placebo: M = 2.47, SD = 1.08). Finally, oxytocin did not influence response latencies, F(1, 67) = 0.26, p = .614 (oxytocin: M = 5.08 s, SD = 1.97 s; placebo: M = 4.85 s, SD = 1.96 s).
Discussion
The results reported here demonstrate that oxytocin stimulates in-group conformity. When asked to rate novel visual stimuli on attractiveness and when in-group and out-group members exhibited opposing preferences, individuals given oxytocin expressed preferences that were closer to those of the in-group than the out-group. This selective in-group bias was absent in the placebo condition and independent of the number of ratings that participants were shown. This finding provides novel evidence that oxytocin is involved in influencing people’s preferences about actual stimuli, complementing earlier work demonstrating that oxytocin alters perceptions of more abstract concepts, such as generosity, trust, and fairness. The results also support the notion that the effects of oxytocin on social behavior are context dependent (Bartz et al., 2011; Kemp & Guastella, 2011). In line with past work demonstrating that oxytocin’s effect on trust is reduced when the other person is described as untrustworthy (Mikolajczak et al., 2010), is unknown (Declerck et al., 2010), or is a member of an out-group (De Dreu et al., 2010), our findings showed that the effects of oxytocin on conformity are limited to the in-group and do not include preferences of the out-group.
Our results raise some interesting questions regarding what processes underlie oxytocin’s influence on in-group conformity. One explanation of the selective in-group conformity bias under oxytocin may be that this neuropeptide stimulates conformity to in-group members by enhancing in-group identification processes (De Dreu et al., 2011); previous work has shown that the perception of shared group membership is both essential and sufficient to establish in-group favoritism (Deutsch & Gerard, 1955; Tajfel, Flament, Billig, & Bundy, 1971). Thus, participants in our study may have identified more with their in-group after receiving oxytocin rather than after receiving a placebo. An additional question is to what extent oxytocin’s effects on in-group conformity depend on social-comparison processes with out-group members. We found that oxytocin induced conformity toward the in-group only when the two groups provided opposing ratings but not when both had similar views. Therefore, oxytocin may play a critical role in mediating in-group identification processes, particularly when the situation involves intergroup comparisons and when group conformity functions to distinguish oneself from members of other groups. This possibility aligns well with previous work suggesting that a primary function of in-group biases is to signal group membership and establish intergroup differentiation (Jetten, Spears, & Postmes, 2004). Future studies could examine whether oxytocin’s effects on conformity are driven by increased in-group affiliation or, rather, whether oxytocin also leads people to move away from the out-group—this is an important question, but these processes cannot be disentangled with the present data.
The minimal group setting employed in the present study was designed to be as modest a manipulation as possible. It is thus not surprising that we did not observe in-group conformity in the placebo condition—previous studies have shown that in-group conformity is stronger when individuals engage in face-to-face interactions and when responses are made public and not in minimal group settings with anonymous private reporting (such as we employed here; Bond, 2005; Deutsch & Gerard, 1955). Accordingly, future research focusing on intergroup settings with reduced anonymity, increased reputation concerns, and potentially stronger in-group identification appears likely to uncover increased conformity under oxytocin than under a placebo.
Future work combining brain-imaging methods with oxytocin administration could reveal whether effects on in-group conformity are mediated via in-group identification processes or whether other mechanisms may also play a role. For example, neuroimaging studies have linked in-group identification to the medial prefrontal cortex, with subregions in this area differentially involved when thinking about self and others (Mitchell, Macrae, & Banaji, 2006). In addition, oxytocin’s effects on in-group conformity may be mediated via brain areas involved in reward processing, such as the striatum (Sanfey, 2007), an area dense with oxytocin receptors (Skuse & Gallagher, 2009) and heavily involved in social-influence processes (Klucharev, Hytonen, Rijpkema, Smidts, & Fernandez, 2009).
The current results extend previous findings regarding oxytocin’s effects on in-group favoritism (De Dreu et al., 2010; De Dreu et al., 2011) into the domain of social influence. Although group membership here was artificially manipulated, neutral stimuli were presented, and participants’ evaluations were kept anonymous, the effect of oxytocin was sufficiently powerful to influence actual preferences and induce in-group conformity. Thus, oxytocin not only promotes in-group favoritism but may even stimulate individuals to conform to the behavior and beliefs of others in their group. The finding that oxytocin stimulates conformity toward in-group members, but not out-group members, provides useful insights into the nature of this hormone and will help in developing a more refined theory of the effects of oxytocin on human social judgment and behavior.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.