
Abstract
Much social-cognitive processing is believed to occur automatically; however, the relative automaticity of the brain systems underlying social cognition remains largely undetermined. We used functional MRI to test for automaticity in the functioning of two brain systems that research has indicated are important for understanding other people’s behavior: the mirror neuron system and the mentalizing system. Participants remembered either easy phone numbers (low cognitive load) or difficult phone numbers (high cognitive load) while observing actions after adopting one of four comprehension goals. For all four goals, mirror neuron system activation showed relatively little evidence of modulation by load; in contrast, the association of mentalizing system activation with the goal of inferring the actor’s mental state was extinguished by increased cognitive load. These results support a dual-process model of the brain systems underlying action understanding and social cognition; the mirror neuron system supports automatic behavior identification, and the mentalizing system supports controlled social causal attribution.
The psychology of social life is vastly complex, supported by innumerable processes that allow us to, among other things, recognize the people around us, recall memories about them, experience feelings toward them, and interact with them. If all of these processes required conscious control, no individual would be able to participate in the social world. Fortunately, many are automatic, initiated by the presence of social stimuli and completed with little conscious intervention (Bargh & Williams, 2006; Lieberman, Gaunt, Gilbert, & Trope, 2002; Smith & DeCoster, 2000). One particularly important social stimulus is an action performed by another person. Two neurocognitive systems are associated with action understanding in humans: the mirror neuron system and the mentalizing system. The study reported here examined the relative automaticity of these systems’ operation during action observation.
The human mirror neuron system comprises areas of the brain that activate during both the execution and the observation of motor actions. It is typically localized to ventral premotor cortex (vPMC), dorsal premotor cortex (dPMC), and anterior intraparietal sulcus (aIPS; Rizzolatti & Craighero, 2004; Van Overwalle & Baetens, 2009). These regions are thought to enable the automatic comprehension of action on the basis of sensory information alone (Coricelli, 2005; Iacoboni et al., 2005; Keysers & Gazzola, 2007; Rizzolatti & Craighero, 2004). Although there is considerable empirical evidence that the mirror neuron system contributes to action comprehension, there is little evidence that this contribution is automatic. In fact, studies in which observers’ goals and attention have been manipulated have actually suggested that controlled processing occurs in areas of the mirror neuron system during action observation (Engel, Burke, Fiehler, Bien, & Rösler, 2008; Hesse, Sparing, & Fink, 2009; Spunt & Lieberman, 2012).
The mentalizing system is a set of brain regions that reliably activates when individuals are instructed to infer another person’s mental state; it includes areas within medial frontal and parietal cortices, temporo-parietal junction, and anterior temporal cortex (aTC; Frith & Frith, 2006; Van Overwalle & Baetens, 2009). Several studies have demonstrated activation of mentalizing system areas in participants given the goal of explaining actions in terms of the actors’ mental states (de Lange, Spronk, Willems, Toni, & Bekkering, 2008; Grèzes, Frith, & Passingham, 2004; Spunt, Falk, & Lieberman, 2010; Spunt & Lieberman, 2012; Spunt, Satpute, & Lieberman, 2011). Although numerous behavioral studies have suggested that the psychological process associated with mentalizing system activity—mental-state inference—is at least partially dependent on controlled processing (Apperly, Riggs, Simpson, Chiavarino, & Samson, 2006; Gilbert, Pelham, & Krull, 1988; Lin, Keysar, & Epley, 2010), other work has suggested that inferences about other people’s mental states may occur spontaneously, in the absence of control (Uleman, Saribay, & Gonzalez, 2008).
Within social neuroscience, the extent to which the operation of the mentalizing system depends on control remains unclear. The reliable association of mentalizing system activation with the top-down goal of inferring mental states suggests controlled processing; however, results from several studies showing stimulus-driven (bottom-up) mentalizing system activation suggest the opposite (Brass, Schmitt, Spengler, & Gergely, 2007; Wagner, Kelley, & Heatherton, 2011; Wheatley, Milleville, & Martin, 2007).
Critically, automaticity is not a unitary construct but instead comprises a set of relatively independent dimensions (Bargh, 1989). One such dimension is efficiency, or the extent to which a process demands attentional resources. Efficient processes can run to completion even when attention is occupied with an unrelated task. Hence, it is possible to assess the efficiency of the mirror neuron system and the mentalizing system by engaging them in a primary task while simultaneously varying the difficulty of an unrelated secondary task (Satpute & Lieberman, 2006). In the present study, participants observed actions performed by another person while undergoing a validated task known to engage the mirror neuron and mentalizing systems (the why/how task; Spunt et al., 2010). In addition, we simultaneously varied the difficulty of a secondary memory task to determine the relative efficiency of mirror neuron system and mentalizing system activity during action observation.
Method
Participants
Nineteen right-handed participants (9 females, 10 males; mean age = 23.0 years, age range = 18–32 years) were recruited from the University of California, Los Angeles (UCLA), subject pool and from the surrounding community. Participants provided informed consent in a manner consistent with the procedures of the UCLA institutional review board. One participant was excluded from our final sample because of a cavernous malformation in the frontal lobe.
Stimuli
Stimuli were 24 silent 5-s videos used in a previous study of action observation (Spunt et al., 2011). All videos featured the same right-handed male actor performing an ordinary object-directed action with his hands, in a natural scene.
Design and procedure
This study employed a 2 × 4 within-subjects factorial design. We manipulated the first factor, cognitive load, by having participants observe the actions shown in the videos while remembering either easy phone numbers (e.g., 555-5555; low load) or difficult phone numbers (e.g., 813-5467; high load; all phone-number stimuli are shown in Table S1 in the Supplemental Material available online). We manipulated the second factor, goal, by instructing participants to adopt one of four goals while observing each action: to passively observe the action (“Just WATCH”; observe goal), to actively recognize the action (“Try to understand WHAT he is doing”; what goal), to understand the motive behind the action (“Try to understand WHY he is doing it”; why goal), or to understand the implementation of the action (“Try to understand HOW he is doing it”; how goal).
Videos were presented to participants in blocks of six (Fig. 1). Prior to viewing the first video in each block, participants viewed two screens. The first screen displayed instructions for the upcoming block (i.e., it manipulated the participant’s goal for the upcoming block); the second displayed either a high-load or a low-load phone number (i.e., it manipulated the level of load for the block). After the final video in each block was presented, participants saw a memory probe and judged whether the presented phone number matched the original number. At the end of each block, participants rated how difficult the block had been overall, using a scale from 1 (not at all) to 4 (very). Each block was preceded and followed by a 12-s rest period, during which a fixation cross was presented. The order of conditions (low-load/observe-goal condition, high-load/observe-goal condition, low-load/what-goal condition, high-load/what-goal condition, low-load/why-goal condition, high-load/why-goal condition, low-load/how-goal condition, and high-load/how-goal condition) was counterbalanced both within and between subjects. Different videos were used for the two levels of the load factor, and the assignment of videos to level of load was counterbalanced across participants. The same videos were used across all levels of the goal factor.

Schematic displaying the time course of an experimental block. This example is from a high-load/why-goal block. ITI = intertrial interval.
We used MATLAB (The MathWorks, Natick, MA) and the Psychophysics Toolbox (Brainard, 1997) to present stimuli and record responses. Participants viewed the screen through LCD goggles and made responses using a four-button box held in their right hand.
Image acquisition
Images were acquired using a Siemens Trio 3.0-T MRI scanner at the UCLA Ahmanson-Lovelace Brain Mapping Center. For each participant, we acquired 494 functional T2*-weighted echoplanar image volumes (EPIs) with the following parameters: slice thickness = 4 mm, gap = 1 mm, 33 slices, repetition time (TR) = 2,000 ms, echo time (TE) = 30 ms, flip angle = 75°, matrix size = 64 × 64, field of view (FOV) = 220 mm. We also acquired a T2-weighted matched-bandwidth anatomical scan (same parameters as the EPIs, with the following exceptions: TR = 5,000 ms, TE = 34 ms, flip angle = 90°, matrix size = 128 × 128) and a T1-weighted magnetization-prepared rapid-acquisition gradient-echo anatomical scan (same parameters as the EPIs, with the following exceptions: slice thickness = 1 mm, 160 slices, TR = 2,300 ms, TE = 2.47 ms, flip angle = 8°, matrix size = 192 × 192, FOV = 256 mm).
Image analysis
We analyzed images using Statistical Parametric Mapping (SPM) software (SPM8; Wellcome Department of Cognitive Neurology, London, England). Functional image volumes were realigned to correct for head motion, normalized into Montreal Neurological Institute space (resampled at 3 mm × 3 mm × 3 mm) using the SPM segmentation routine, and smoothed with an 8-mm full-width, half-maximum Gaussian kernel.
A general linear model was used to model the effects of load and goal for each participant. We included eight effects of interest corresponding to the eight conditions. Each block was modeled as an epoch spanning the onset of the first video to the offset of the last video and was convolved with a canonical (double-gamma) hemodynamic response function. Each model included covariates of no interest modeling preblock display screens, postblock probes, and the estimates of head motion. The time series was high-pass filtered to 1/160 Hz, and autocorrelation was modeled as a first-order autoregressive, or AR(1), process. Following estimation, we entered participants’ contrast images for the effect of each regressor of interest into random-effects analyses using a flexible factorial repeated measures analysis of variance (ANOVA; within- subjects factors: load, goal; blocking factor: subject).
Given our a priori interest in the mirror neuron and mentalizing systems, we restricted our analysis to an independently defined mask of these systems (details about the mask and whole-brain results are provided in the Supplemental Methods section of the Supplemental Material). We used Analysis of Functional and Neural Images (AFNI) software (National Institute of Mental Health, Bethesda, MD; http://afni.nimh.nih.gov/) to perform a Monte Carlo simulation within this mask (residual smoothness estimated using the AFNI 3dFWHMx program) and found that a voxel-wise p value of .005 combined with an extent threshold of 23 voxels corresponded to a cluster-wise false-discovery rate of .05. Statistical maps were surface rendered using the SPM surfrend toolbox. Percentage signal change was calculated using the MarsBaR SPM toolbox.
Results
Difficulty and accuracy
Paired-samples t tests confirmed the efficacy of the load manipulation. Participants’ ratings of the task’s difficulty increased under high load (M = 2.38, SD = 0.39) compared with low load (M = 1.41, SD = 0.38), t(19) = 11.565, p < .001, and accuracy decreased under high load (M = 83%, SD = 15%) compared with low load (M = 97%, SD = 5%), t(18) = 4.158, p = .001. A repeated measures ANOVA (for full results, see the Supplemental Results section of the Supplemental Material) revealed a main effect of goal on difficulty; this effect was driven by decreased ratings of difficulty for the observe goal relative to the other three goals. Accuracy showed no main effect of goal and no Load × Goal interaction.
Neuroimaging results
If the mirror neuron system operates automatically, it should activate during action observation regardless of the observer’s goal and level of load. Therefore, we used a conjunction analysis to determine which regions were active in all eight conditions. This analysis revealed activation in several regions of the mirror neuron system, including bilateral vPMC, left dPMC, and left aIPS (see Fig. 2 and Table 1). These regions’ response to actions appeared to be relatively unaffected by increased cognitive load. To formally test that this was the case, we used an F contrast to determine if load modulated activity in the mirror neuron system for any of the four goals. This analysis identified no regions within the mirror neuron system, even at a liberal threshold (p < .05, uncorrected). In contrast, two regions of the mentalizing system, medial prefrontal cortex (mPFC) and left aTC, were significant at our a priori threshold (Table 1).

Brain regions showing evidence of efficient operation during action understanding. The surface renderings show areas of significant activation in left (L) and right (R) ventral premotor cortex (vPMC), dorsal premotor cortex (dPMC), and anterior intraparietal sulcus (aIPS). The graphs show percentage signal change from fixation baseline in each of these regions in all conditions. Error bars represent standard errors. Obs = observe.
Peak Coordinates From Significant Clusters Observed in All Analyses

Note: Results are cluster-wise corrected at a false discovery rate of .05 within an a priori mask of the mirror neuron and mentalizing systems. Coordinates are for observed local maxima that were separated by at least 20 mm. Montreal Neurological Institute (MNI) coordinates are reported in the left-right (x), anterior-posterior (y), and inferior-superior (z) dimensions, respectively. k = cluster voxel extent.
We restricted our remaining analyses to the how-goal and why-goal conditions, because our past research showed that these goals produce the strongest goal-related response in the mirror neuron and mentalizing systems, respectively. For these analyses, we examined whether any regions were selectively recruited when participants adopted either the how goal or the why goal, but were also sensitive to the availability of controlled processing resources within a given condition. First, we compared the low-load/how-goal condition against the three other conditions of interest (high load/how goal, low load/why goal, high load/why goal) to identify regions that were recruited by the how goal but were sensitive to load. No regions within either the mirror neuron system or the mentalizing system demonstrated activation in this analysis. Next, we compared the low-load/why-goal condition against the three other conditions of interest (high load/why goal, low load/how goal, high load/how goal) to identify regions that were recruited by the why goal but were sensitive to load. This analysis revealed activation within dorsomedial prefrontal cortex (dmPFC) and left aTC—both of which are regions of the mentalizing system—but no activation within the mirror neuron system (Fig. 3). Both of these regions were also identified by the load-by-goal interaction, as was mPFC (Table 1).
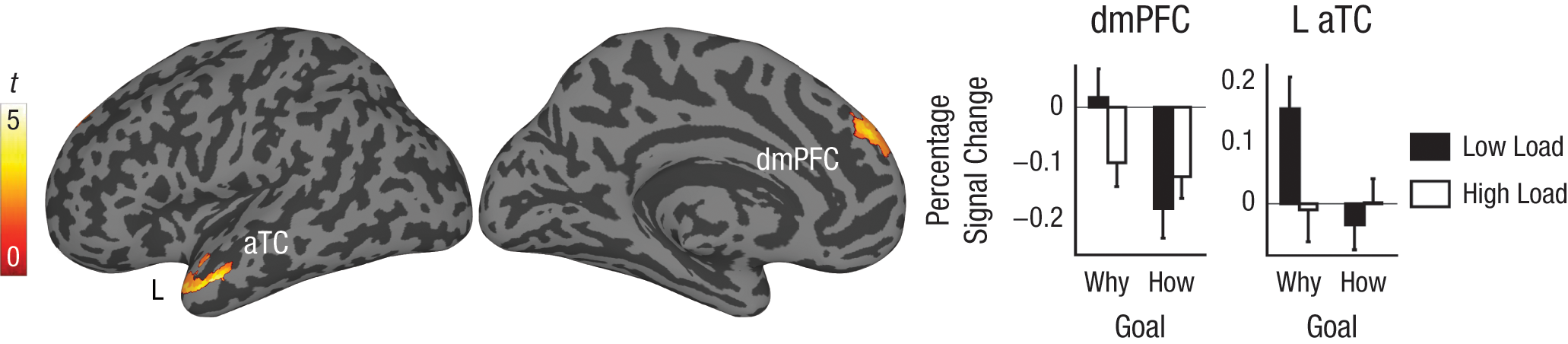
Brain regions showing evidence of nonefficient operation during action understanding. The surface renderings show regions emerging in the contrast comparing the low-load/why-goal condition with the high-load/why-goal, the low-load/how-goal, and the high-load/how-goal conditions combined; these regions were left anterior temporal cortex (L aTC) and dorsomedial prefrontal cortex (dmPFC). The graphs show percentage signal change from fixation baseline for each region in the conditions of interest. Error bars represent standard errors.
Discussion
We report evidence regarding the automaticity of mirror neuron system operation and mentalizing system operation during action observation. The activation of areas within the mirror neuron system—namely, bilateral vPMC, left dPMC, and left aIPS—was unaffected by load regardless of the observer’s goal. In contrast, activation of areas within the mentalizing system—namely, dmPFC and left aTC—was modulated by load only when participants were prompted to attribute observed actions to a motive (i.e., when participants adopted the why goal), not when actions were understood in terms of their implementation (i.e., when participants adopted the how goal). These results support a dual-process model, whereby the mirror neuron system supports relatively automatic behavior identification and the mentalizing system supports relatively controlled components of social causal attribution (Coricelli, 2005; Spunt & Lieberman, 2012).
Although these results provide support for the popular notion that the mirror neuron system operates automatically during action observation (Coricelli, 2005; Keysers & Gazzola, 2007; Rizzolatti & Craighero, 2004), they may appear inconsistent with extant research demonstrating that activation of the mirror neuron system during action observation is modulated by the observer’s goals and attention (Engel et al., 2008; Hesse et al., 2009; Spunt & Lieberman, 2012). However, this inconsistency can be resolved by acknowledging the multidimensional nature of automaticity (Bargh, 1989). As noted earlier, one dimension of automaticity is efficiency, or the extent to which a process can run to completion without consuming attentional resources. A separate dimension of automaticity is controllability; studies manipulating the observer’s goals speak primarily to this dimension. Efficient processes need not be uncontrollable (Bargh, 1989). Hence, it may be the case that the mirror neuron system responds efficiently when spatial attention is directed at an action, but that this response is nevertheless subject to some degree of control.
Indeed, as shown in the plots in Figure 2, our results suggest a profile of activity in the mirror neuron system that is at once efficient (i.e., insensitive to load) and goal dependent. In fact, testing the conjunction of the low-load/how-goal > low-load/observe-goal and high-load/how-goal > high-load/observe-goal contrasts revealed a significant effect in left vPMC (Table 1, Fig. S3 in the Supplemental Material). This result illustrates that efficient activation in response to a stimulus does not preclude the top-down modulation of mirror neuron system activation by changes in the observer’s attention and goals (Lieberman, 2011).
Mentalizing system activation was associated with the goal of understanding an action in terms of the actor’s mental state. This result replicates findings demonstrating that the mentalizing system supports mental-state inference in the context of action observation (de Lange et al., 2008; Grèzes et al., 2004; Spunt et al., 2010; Spunt & Lieberman, 2012; Spunt et al., 2011); moreover, it significantly adds to work on the subject by showing that this effect disappears under conditions of high cognitive load. One alternative explanation for this finding might be that areas of the mentalizing system often deactivate in response to increasing task difficulty (Spreng, Mar, & Kim, 2009); however, this explanation is untenable, given that modulation by load in the mentalizing system was goal dependent, observed only when observers adopted the goal of inferring an actor’s mental state.
Hence, this interference effect is more consistent with inefficient (i.e., effortful) operation of the mentalizing system during mental-state inference, and it converges with results from neuroimaging studies showing that mentalizing system activation parametrically tracks the amount of attention demanded by a primary social-cognitive task (Meyer, Spunt, Berkman, Taylor, & Lieberman, 2012) and is disrupted by the introduction of a secondary task (den Ouden, Frith, Frith, & Blakemore, 2005; Rameson, Morelli, & Lieberman, 2011). Moreover, the observed interference effect converges with theoretical and behavioral results suggesting that components of social causal attribution and belief reasoning require control (Apperly et al., 2006; Coricelli, 2005; Gilbert et al., 1988; Keysers & Gazzola, 2007; Lin et al., 2010).
There is evidence that the mentalizing system is capable of operating in an efficient, stimulus-driven fashion. It is known that inferences about mental states can occur spontaneously, solely on the basis of sensory input about behavior (Gilbert et al., 1988; Uleman et al., 2008), and manipulations intended to induce spontaneous social inferences activate the mentalizing system (Ma, Vandekerckhove, Overwalle, Seurinck, & Fias, 2011). Moreover, individuals who are dispositionally inclined to adopt other people’s perspectives show increased spontaneous activation of the mentalizing system during social perception (Rameson et al., 2011; Wagner et al., 2011). Intriguingly, adults with Asperger’s syndrome can infer other people’s mental states when instructed to do so, but fail to do this spontaneously (Senju, Southgate, White, & Frith, 2009), a pattern suggesting that the relative efficiency of mentalizing system activation may be an important individual differences marker for social expertise. Future research and theorizing on the social brain will profit through the careful consideration of the conditional nature of the operation of its underlying systems, including the mirror neuron and mentalizing systems.
Footnotes
Acknowledgements
We thank several anonymous reviewers for helpful comments and the staff of the Ahmanson-Lovelace Brain Mapping Center for support.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.