
Abstract
Neurological changes in older adults suggest that their mental representation of peripersonal space, the space around their bodies, might be degraded compared with that of young adults. These changes may lead to differences in how attention is allocated within peripersonal space, affecting how older adults plan and guide their actions. In the present study, we show that there are indeed profound differences between the spatial representations of older and young adults: In a task involving simple hand movements, young adults adopted an attentional reference frame centered on the hand, and older adults adopted a reference frame centered on the body. Such differences may help to explain age-related changes observed in the performance of many common movement tasks.
As the U.S. population has aged, problems common in older adults, such as decreased mobility, have become important issues. For example, an estimated 60% to 70% of the elderly population reports difficulty with activities of daily living, such as eating and bathing (Desai, Lentzner, & Weeks, 2001), and many show deficiencies in performing goal-directed hand movements (Carmeli, Patish, & Coleman, 2003). Because an accurate mental representation of the environment is vital to an individual’s ability to act and move appropriately, it is important to understand how older adults attend to nearby space. Of particular interest is how older adults construct and use attentional reference frames for the space within which they act (i.e., peripersonal space), as this can be a determinant of their success in reaching desired objects (e.g., a cup of tea) while avoiding hazardous obstacles (e.g., a hot stove) and unwanted collisions. The little work that has been done to date suggests that older adults represent space differently than young adults do (Fujii, Fukatsu, Yamadori, & Kimura, 1995; Ghafouri & Lestienne, 2000), but it is unknown whether this is because the elderly use different attentional reference frames and whether such a difference affects how older adults plan and guide actions in peripersonal space. We examined those issues using a selective-reaching task.
Aging is associated with both behavioral changes and corollary neurological changes. Many of the brain areas that form the neural circuitry for representing peripersonal space show changes during senescence. The space surrounding the hands (perihand space), in particular, is primarily coded by the premotor cortex, intraparietal sulcus, and precuneus (e.g., Cavanna & Trimble, 2006). These three areas are highly involved in visually guided hand actions like reaching and grasping (Astafiev et al., 2003; Filimon, Nelson, Huang, & Sereno, 2009) and in creating attentional reference frames that are used to guide such actions (Cohen & Anderson, 2002). Older adults exhibit volumetric declines in the parietal cortex (Lehmbeck, Brassen, Weber-Fahr, & Braus, 2006; Xu et al., 2000) and intraparietal sulcus (Kochunov et al., 2005), as well as white-matter loss in the parietal lobe and precuneus (Lehmbeck et al., 2006). These neurological changes in older adults suggest that their representations of peripersonal space may be compromised relative to those of young adults and that, consequently, young and older adults might encode and attend to peripersonal space in fundamentally different ways.
In the present study, we examined the attentional reference frames adopted by older adults for executing a reach-and-point action, such as the movement needed to dial a phone. Young adults have been shown to adopt an action-centered reference frame for such movements, such that selective attention is preferentially allocated along the path of the movement (e.g., Pratt & Abrams, 1994; Tipper, Lortie, & Baylis, 1992). An action-centered reference frame might lead to more successful movements in part because representations of obstacles along the movement path are enhanced. The use of an action- centered reference frame, however, is presumed to rely on current and spatially accurate knowledge of hand position and the objects present in peripersonal space. As noted earlier, such information might not be as accessible to older adults as it is to young adults. In this study, we compared the performance of young and older adults on a selective-reaching task, with the expectation that young adults would have slowed responses when a distractor appeared along the movement path, whereas older adults’ responses would not be slowed by movement-path distractors. We found that young adults did adopt an action-centered reference frame, but older adults appeared to use a body-centered reference frame. As we discuss later, this difference may explain, in part, the age-related deficits observed in many movement tasks.
Method
Participants
Thirty healthy, young adult participants (M = 19.78 years, range = 18–23; 8 males, 22 females; 24 Caucasians, 6 people of other races) were recruited from the undergraduate community at Washington University in St. Louis. Thirty-two healthy, community-dwelling older adults (M = 73.72 years, range = 65–86; 7 males, 25 females; 27 Caucasians, 5 people of other races) were recruited from the Washington University Department of Psychology’s Older Adult Volunteer Pool. As compensation, young adults received course credit, and older adults received $10 an hour. Each session lasted approximately 2 hr. All participants had normal or corrected-to- normal vision. The groups had similar levels of education (young adults: M = 13.75 years; older adults: M = 15.75 years).
Apparatus and procedure
We used a variation of the selective-reaching task employed by Tipper and his colleagues (Meegan & Tipper, 1998; Tipper et al., 1992). Participants were seated at a 15-in. LCD computer monitor, tilted 44° in relation to the table upon which it sat. As shown in Figure 1, the monitor displayed nine boxes arranged in a 3 × 3 grid. Each box measured 20 × 20 pixels, and adjacent boxes were separated by 18 pixels. Viewing distance was approximately 45 cm, with head position unconstrained to permit natural reaching movements.
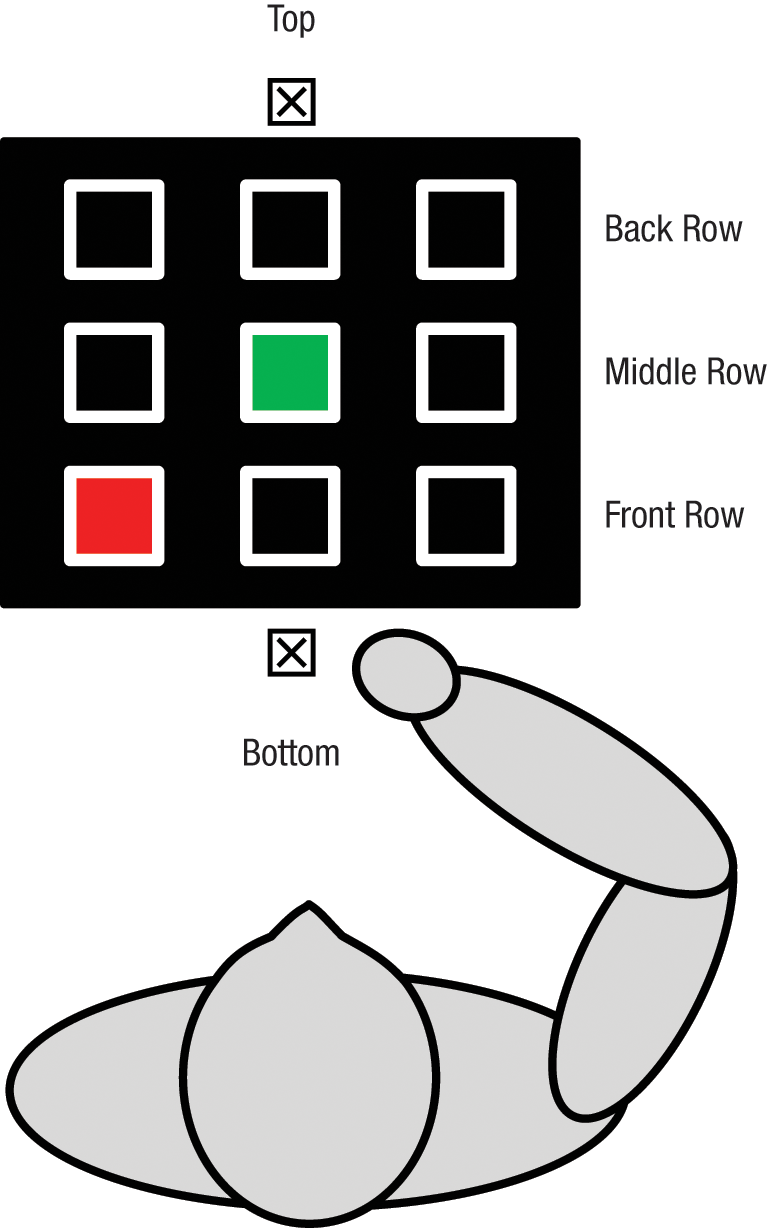
Experimental setup. On each trial, participants started with their right hand at one of the two home locations (designated by the squares containing an X) and reached to touch the green target among nine boxes arranged in a 3 × 3 grid. On some trials, the grid included a red distractor.
At the beginning of each trial, each of the nine boxes appeared as a white outline on a black background. Participants first moved their index finger to a “home location,” a piece of fabric (2.54 cm per side) mounted at either the bottom or the top of the display. One of the black boxes turned green 2,000 ms after the participant’s finger reached the home location; this box was the target. On trials without a distractor, all other boxes remained black with a white outline; on trials with a distractor, one of these other boxes turned red at the same time the target box turned green. Participants were instructed to reach with their right hand and touch the target with their index finger as quickly and accurately as possible (while ignoring the distractor, if one was present). After participants completed their reach, they maintained their hand position at the target location for an additional 2,000 ms, after which the boxes disappeared from the monitor. This signaled the end of the trial and cued participants to return their hand to the home location. We gave feedback to participants if they began a reaching movement too early (< 100 ms after the target appeared) or if no movement was detected.
We monitored participants’ hand movements throughout the experimental session using an Ascension Technology (Burlington, VT) Flock of Birds sensor (25.4 × 25.4 × 20.3 mm). The sensor was attached to the top of the index finger of a glove that participants wore on their right hands. The system sampled the sensor’s position in three dimensions at a rate of 100 Hz.
Participants completed two blocks of 416 trials, one block for each home location. The order of the blocks was counterbalanced across participants. If present, the distractor could appear in the front, middle, or back row. Likewise, the target could appear in the front, middle, or back row. When a distractor was present, it was immediately above, below, next to, or diagonally adjacent to the target (i.e., either in the same row or in an adjacent row). The only trials of interest were those in which the target appeared in the middle row. In each block, there were 288 such trials; 48 contained no distractor, 48 had a distractor in the same row as the target, 96 had a distractor in the front row, and 96 had a distractor in the back row. Of the remaining 128 trials (in which the target appeared in either the front or the back row), 28 had no distractor, 36 had a distractor in the same row as the target, and 64 had a distractor in the middle row. At the beginning of each block, participants were given 16 practice trials containing various combinations of target and distractor locations. The experimenter was present during the practice trials to provide verbal feedback and correct performance if necessary.
The dependent variable was the total response time (TRT) for each trial, measured from the onset of the target (and distractor, if present) until the end of the participant’s hand movement. The end of the movement was determined by a velocity criterion. Thus, TRT was a combination of reaction time (latency of movement onset following display onset) and movement duration. Only TRTs from trials in which the target appeared in the middle row and a distractor was present were considered. Trials in which the target appeared in other locations were included in the design to ensure that participants were attending to all locations on the screen, as well as to prevent participants from planning a movement prior to target onset.
Results
Seven older adults were excluded from analysis because they had neurological disorders (n = 2), restricted arm or shoulder mobility (n = 2), or a high error rate (> 10%; n = 3). The remaining older-adult sample had a mean age of 73.00 years. Four young adults were excluded because a technical error resulted in the loss of their data, and 2 more were excluded because they had high error rates (> 10%). The remaining young-adult sample had a mean age of 19.35 years.
As noted, all results are based on trials in which the target appeared in the middle row of the display and a distractor was present. Mean TRTs and error rates are shown in Figure 2. A mixed-factors 2 (home location: bottom, top) × 3 (distractor location: front row, middle row, back row) × 2 (group: young adult, older adult) analysis of variance was conducted on TRTs. The most important results involve the Group × Home Location × Distractor Location interaction, F(2, 46) = 6.51, p = .003, η p 2 = .22. Young adults exhibited a performance pattern in which the TRT associated with a given distractor location depended on the starting position of the hand: When the home location was at the bottom of the display, TRTs were longest when a distractor appeared in the front row. However, when the home location was at the top of the display, TRTs were longest when a distractor appeared in the back row.

Mean total response time (top row) and mean error rate (bottom row) for young and older adults as a function of distractor location and starting hand position (home location). Only data for trials in which the target appeared in the middle row and a distractor was present are included. Error bars represent within-subjects 95% confidence intervals (Morey, 2008).
Young adults’ slowed reaching movements in the presence of a near-hand distractor indicate that they planned and produced their movements using an action-centered reference frame. Older adults, however, did not show this pattern. For older adults, the effect of distractors on TRTs did not depend on the starting hand position: TRTs were always longest when the distractors were in the front row. Thus, the older adults planned and produced their movements using a body-centered reference frame. Post hoc analyses supported our conclusions: Young adults showed a significant Home Location × Distractor Location interaction, F(2, 22) = 11.52, p < .001, η p 2 = .51, but did not show a main effect of distractor location, F(2, 22) = 3.25, p > .05, η p 2 = .25. In contrast, older adults did not show a Home Location × Distractor Location interaction, F(2, 23) = 1.18, p > .10, η p 2 = .09, but did show a main effect of distractor location, F(2, 23) = 15.01, p < .001, η p 2 = .57.
Results of post hoc t tests comparing TRTs across home locations at each distractor location are in the Supplemental Material available online. Additionally, in the Supplemental Material, we report the patterns of reaction times and movement times separately for each age group, report overall main effects, and provide evidence ruling out age-related strategies and the side on which the distractor appeared as contributors to the observed age-related differences in performance. Finally, the Supplemental Material presents additional results for accuracy (error rates). Results of all analyses were consistent with our conclusions.
One concern, especially in a heterogeneous sample such as our older-adult sample, is that a small number of participants may show a pattern of performance counter to the pattern exhibited by the majority, which could produce the appearance of an age-related change that does not actually exist. To verify that this did not occur in our study, we computed a summary statistic indexing the relative degree to which participants used an action-centered reference frame rather than a body-centered reference frame. We calculated this index of interference by considering only trials in which the home location was at the top of the screen. Mean TRTs for trials with a distractor in the front row were subtracted from mean TRTs for trials with a distractor in the back row. If a participant had used an action-centered reference frame, this index would be a positive number; if a participant had used a body-centered reference frame, this index would be a negative number. As Figure 3 shows, the young-adult sample had a positive index of interference on average (M = 27.51, SE = 4.53), and the older-adult sample had a negative mean index (M = −12.75, SE = 3.40), t(47) = 6.75, p < .001. This indicates that our results were not driven by outliers in either age group.
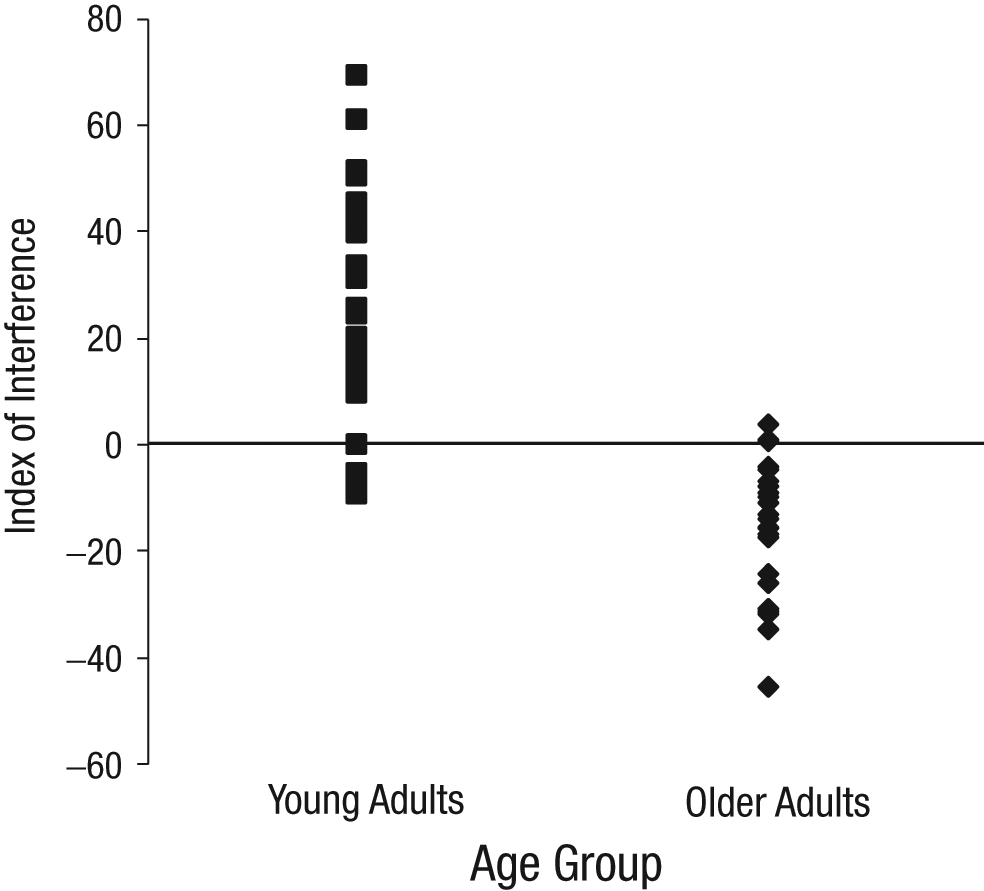
Index of interference as a function of age group. Each plotted point represents the value for a single participant. Positive values indicate the use of an action-centered reference frame; negative values indicate the use of a body-centered reference frame.
Discussion
Previous research has shown that young adults use an action-centered reference frame when reaching with their hands toward a target, indicating that young adults’ attention is preferentially allocated to the path of the hand movement (Tipper et al., 1992). The present study leads to the same conclusion, as distractors along the movement path slowed the participants’ reaching more than distractors outside the path. This was true regardless of the starting location of the hand, and hence of the direction of the movement. An action-centered reference frame may be adopted because perihand space has a distinct neural representation (Filimon et al., 2009; Rizzolatti, Luppino, & Matelli, 1998), and using the hand to perform an action such as reaching engages specialized brain areas. The use of an action-centered reference frame might allow the movement path to be planned more accurately and may facilitate the detection of obstructions that could interfere with the movement. However, older adults exhibit declines in the cortical areas that underlie such representations (Kochunov et al., 2005; Lehmbeck et al., 2006). These declines may make the use of an action-centered reference frame difficult or impossible. Indeed, in the present study, older adults showed the longest TRTs when the distractor was near their bodies, even when the hand movement did not traverse that region. This pattern reveals their use of a body-centered reference frame.
The pattern of performance for older adults was consistent with a decline in hand-centered representations of peripersonal space. Specialized perihand representations must be spatially accurate to support actions such as reaching, grasping, and object manipulation (Graziano, Yap, & Gross, 1994). Individuals give attentional priority to objects near their hands because they may want to interact with or avoid these objects, and this may be why young adults appear to automatically code the location of objects in reference to their hands when reaching (Meegan & Tipper, 1998; Tipper et al., 1992). If perihand representations become less flexible or less spatially accurate with age, the dominant reference for spatial coding might move from the hands to the trunk. This change would lead to older adults adopting a body-centered reference frame, which in turn would influence how they attend to and interact with their environment. Older adults are known to have problems with tasks requiring manual dexterity and goal-directed hand movements (Carmeli et al., 2003; Pratt, Chasteen, & Abrams, 1994). Although previous studies have found age-related declines in sensory and musculoskeletal function (Ranganath, Siemionow, Sahgal, & Yue, 2001), the present findings suggest that declines in perihand representations are also a factor that should be considered as a contributor to older adults’ impaired movements.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by Grant AG0030 from the National Institute on Aging.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.