
Abstract
Metacognition can be defined as knowing what one knows, and the question of whether nonhuman animals are metacognitive has driven an intense debate. We tested 3 language-trained chimpanzees in an information-seeking task in which the identity of a food item was the critical piece of information needed to obtain the food. The chimpanzees could either report the identity of the food immediately or first check a container in which the food had been hidden. In two experiments, the chimpanzees were significantly more likely to visit the container first on trials in which they could not know its contents but were more likely to just name the food item without looking into the container on trials in which they had seen its contents. Thus, chimpanzees showed efficient information-seeking behavior that suggested they knew what they had or had not already seen when it was time to name a hidden item.
Keywords
Metacognition can be defined as knowing what one knows (Flavell, 1979; Koriat, 1993; Nelson, 1992; Schwartz, 1994). Humans use metacognition frequently, as when we panic upon seeing an acquaintance whose name we cannot recall or when we waver before answering the million-dollar question in a game show and try to decide whether to “phone a friend” or trust our own knowledge. In essence, a metacognitive system surveys what necessary or relevant information a cognitive system has or does not have and then generates responses accordingly. If one knows the answer to a question or one can decide what to call something, an answer is generated. But if information is incomplete or an answer is not clearly indicated, the metacognitive system generates responses to avoid responding, to seek additional information, or to request help.
Adult humans can show excellent metacognitive monitoring. Some nonhuman animal species have been reported to also show behavioral patterns that might reflect aspects of metacognition, such as information-seeking behavior or uncertainty monitoring (Basile, Hampton, Suomi, & Murray, 2009; Beran & Smith, 2011; Call & Carpenter, 2001; Foote & Crystal, 2012; Hampton, 2001; Kornell, Son, & Terrace, 2007; Smith, Beran, Redford, & Washburn, 2006; Smith et al., 1995; Smith, Shields, Allendoerfer, & Washburn, 1998; Smith, Shields, Schull, & Washburn, 1997), but the degree to which nonhuman animals’ performance reflects metacognition like that shown by humans is less clear and has driven an intense debate (see Carruthers, 2008, 2009; Crystal & Foote, 2009; Smith, 2009; Smith, Beran, Couchman, Coutinho, & Boomer, 2009; Smith, Beran, Coutinho, & Couchman, 2008; Smith, Couchman, & Beran, 2012; Staddon, Jozefowiez, & Cerutti, 2009).
One influential and useful paradigm in this area of inquiry is the information-seeking task. Typically, animals are exposed to two kinds of situations—one in which they are given the information they need to make an accurate response and one in which more information is needed before they can respond accurately. For example, Call and Carpenter (2001) presented orangutans, chimpanzees, and children with a test in which the goal was to reach into a container that held a prized reward. In some trials, the apes or children were shown exactly where the item was hidden. In other trials, the hiding was done out of view, so the apes or children could not know where the item was hidden without moving themselves into position to look into each of the possible hiding locations to find the item before reaching for it. The prediction was that if children and apes knew when they knew the location, they would reach without looking. And if they knew when they did not know where the item was, they should look first and then reach after locating it. This is exactly the pattern that emerged in the responses for all three species. Subsequently, other species were tested, and some showed the potentially metacognitive pattern of responding (gorillas, orangutans, chimpanzees, bonobos: Call, 2010; rhesus monkeys: Hampton, Zivin, & Murray, 2004), whereas others did not (capuchin monkeys: Basile et al., 2009; Paukner, Anderson, & Fujita, 2006; dogs: Bräuer, Call, & Tomasello, 2004).
Additionally, concerns were raised about the test itself and whether results found using the test required a metacognitive explanation. For example, Carruthers (2008) argued that animals could succeed—using only first-order mental states—in reaching immediately when they should and in looking further to collect information when they should. According to this argument, when an animal sees food in a tube, it believes food is in the tube. It also believes that if it selects that tube, food will be the result, and of course the animal desires food. Therefore, it reaches immediately for the tube associated with these beliefs about getting food. When the baiting is not observed, the animal has no food-related beliefs attached to any of the tubes, so it searches until such beliefs emerge because food is finally seen in one of the tubes. Put more simply, animals that lack information about the location of the food engage in search behavior until they locate the reward.
Using associative terminology, the parallel argument could be made that subjects learn a rule such as “If I see food, then I don’t look; if I don’t see food, then I do look” on the basis of reinforcement histories of these different behavior chains in different contexts. Crystal and Foote (2011) also offered alternative explanations of the typical “metacognitive” results in these types of experiments by positing that two relatively simple principles could guide an animal’s behavior without need of metacognition. According to the first principle, animals might have a “look before you go” response that supersedes random searches in space. The second principle assumes that spatially guided behavior should follow a rule of going to wherever something good is located. Together, these two principles could potentially account for performance on information-seeking tasks without a need for metacognition.
There are counterarguments to these criticisms (e.g., Call, 2010), but there are also ways to get around the problem by changing the task. For example, one can ensure that a subject always has the belief that there is a valuable item in a container but sometimes does not know what that item is. We used this approach in our first experiment with language-trained apes capable of communicating the identity of items.
Rather than requiring chimpanzees to determine where an item was hidden, we required them to name what was in a container so that there was always a belief that food was in the container and there was always a desire to get the food that was in the container. However, there was the restriction that items could be obtained only if they were properly named. Thus, if a chimpanzee knew what was in the container, it could name the item immediately. By contrast, if it did not, it would have to approach the container—a behavior that was costlier in terms of time and effort—determine what was inside, and then name the item. Thus, the missing information was not about where the food reward was and not about its value (all food rewards were valued), but about what the identity of the item was—a piece of information that had to be communicated to an experimenter. This method eliminated concerns about spatial gradients and default responses of looking before one chooses a container (Crystal & Foote, 2011) because the goal was not to find the item, but to name it.
Even success in this new variation of the task could be attributed to a multicomponent rule—for instance, chimpanzees may have learned to approach the container if they had not seen any food but to name the food immediately if they had seen it at the start of the trial. To control for this possibility, our second experiment involved always showing the food in one container without showing the food in a second container and then choosing one of the containers for placement in the enclosure where naming took place. Thus, chimpanzees could not simply name whatever they saw in the first part of the trial, because sometimes that item stayed in the container that was not moved to the naming area, and they could not use a rule such as “Look when no food has been seen, and name when any food has been seen,” because food was seen on every trial. Instead, the chimpanzees had to attend to whether the container that was moved was the one holding the item they had seen, and if it was, they could name that food item immediately. However, if the container that was moved had not been opened to show its contents, the chimpanzees would have to instead move to the container, look inside, and then name the item. High performance from the outset of experimental sessions would discount alternative explanations for the chimpanzees’ success in Experiment 1 and strengthen the conclusion that they used flexible information-seeking behaviors to accompany their knowledge states in a manner reflective of metacognitive monitoring.
Experiment 1
Method
Subjects
Three chimpanzees, Lana (female, 41 years old), Sherman (male, 38 years old), and Panzee (female, 25 years old) were tested. All 3 chimpanzees had been trained from an early age to use lexigram symbols to request and label objects, actions, locations, and individuals, as well as to respond to requests by humans using those symbols (Brakke & Savage-Rumbaugh, 1995; Rumbaugh, 1977; Rumbaugh & Washburn, 2003; Savage-Rumbaugh, 1986). All 3 chimpanzees had participated in a large number of previous experiments, some of which made use of lexigram symbols to assess various aspects of cognition (e.g., Beran, Savage-Rumbaugh, Pate, & Rumbaugh, 1999; Heimbauer, Beran, & Owren, 2011; Menzel, 1999).
The chimpanzees were tested in their home-cage area, which included indoor enclosures and outdoor yards (see Fig. 1). Each chimpanzee was tested while singly housed, and tests occurred two to four times per week between 10:30 a.m. and 1:00 p.m.
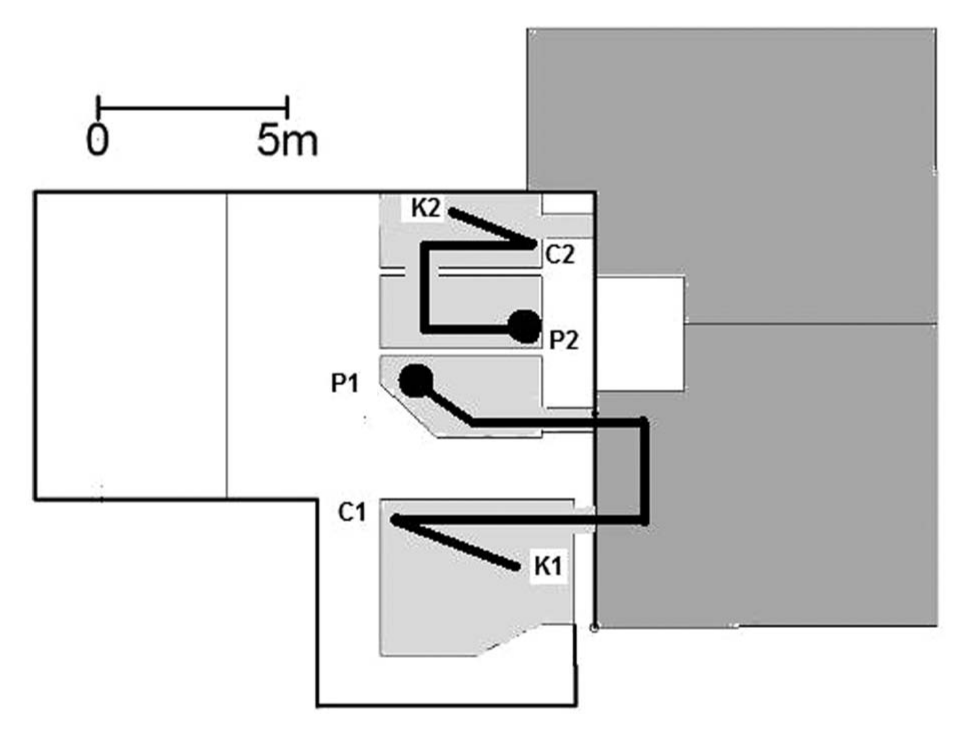
Schematic illustration of the test area. The areas shaded with light gray represent indoor-housing areas for the chimpanzees (Sherman, Panzee, and Lana). The areas shaded in dark gray represent outdoor yards for the chimpanzees. The experimenter either showed or did not show the contents of a container in an initial location (P1: Sherman and Panzee; P2: Lana) before moving the container to a second location (C1: Sherman and Panzee; C2: Lana). The chimpanzees could then use a keyboard (K1: Sherman and Panzee; K2: Lana) to indicate the contents of the container. The dark circles near the two initial locations represent the starting locations of the chimpanzees (circle closest to P1: Sherman and Panzee; circle closest to P2: Lana), and the dark lines show the routes the chimpanzees had to take if they chose to first view the contents of the container before they went to the keyboard to name the item in the container.
Design and procedure
At the outset of each trial, Experimenter 1 moved out of view of the chimpanzee and placed a single food item in an opaque container. This item was always a preferred food item that the chimpanzee could reliably name with its corresponding lexigram. Experimenter 1 then moved within view of the chimpanzee, removed the lid from the container, and made one of two motions—either tilting the container toward the chimpanzee so that it could see the contents of the container or tilting the container away from the chimpanzee so that its contents remained unknown. Experimenter 1 then moved to a location adjacent to the area the chimpanzee was currently in and placed the container on a chair so that its contents could be viewed only from a particular angle in that second location (Fig. 1).
Experimenter 1 left the test area for the remainder of the trial. The chimpanzee then had to leave its original area and move through space such that it lost visual contact with the container for a period of time before arriving in the second location. Once there, the chimpanzee could take one of two routes—moving directly to the lexigram keyboard to indicate the contents of the container or moving instead toward the container to a position from which its contents could be viewed. Critically, the chimpanzee could not see what was in the container from the keyboard location. Thus, moving to the container was costly, adding approximately 15 s of delay, on average, beyond the time it took to just move to the keyboard and select a lexigram.
At the outset of each trial, Experimenter 2 stood in an area of the laboratory from which he could read any responses made by a chimpanzee using a lexigram keyboard on which there were 256 individual lexigrams. Experimenter 2 was unaware of what was in the container and did not know whether the chimpanzee had been shown the contents of the container or not. This experimenter recorded whether the chimpanzee first approached the container or the keyboard and what lexigram was indicated using the keyboard (on all trials, the chimpanzees indicated only a single item using the keyboard). After a keyboard response, Experimenter 2 moved to the container and determined if the item inside matched what the chimpanzee had indicated on the lexigram board. If it did, that item was given to the chimpanzee. If it did not, the item was removed and the chimpanzee did not receive anything.
Each session consisted of four trials, two in which the contents of the container were shown to the chimpanzee and two in which the chimpanzee received no information about the item in the container. Only a subset of highly preferred food items with corresponding lexigrams was used, including bananas, juice, oranges, M&M’s candies, apples, bread, soda, sweet potatoes, kiwis, raisins, peanuts, grapes, strawberries, and primate biscuits. Foods were chosen at random across trials. Each chimpanzee completed 10 sessions, for a total of 40 trials. Because of experimenter error, Panzee and Sherman were given one extra nonvisible trial, for a total of 41 trials. Those additional trials were included in the analyses.
Data analysis
We conducted a chi-square test of independence to assess the relationship between presentation condition (visible vs. nonvisible) and subject’s first response (moving to the container vs. moving to the keyboard). We expected that subjects would more often visit the container first when they had not seen its contents (nonvisible condition) and that they would more often visit the keyboard first when they had seen the contents of the container (visible condition).
Results
Figure 2 shows the performance of each chimpanzee. All chimpanzees were statistically more likely to visit the keyboard first in the visible condition and to visit the container first in the nonvisible condition—Sherman: χ2(1, N = 41) = 23.89, p < .001; Panzee: χ2(1, N = 41) = 21.39, p < .001; Lana: χ2(1, N = 40) = 26.67, p < .001. This behavioral pattern emerged in the very earliest trials: Lana made the predicted response in seven of the first eight trials, Panzee did so for six of the first eight trials, and Sherman did so for six of the first eight trials.

Responses (moving to the keyboard first or to the container first) of each chimpanzee as a function of food-presentation condition (visible vs. nonvisible) in Experiments 1 (a) and 2 (b).
Across the 112 trials in which the chimpanzees either saw the food item and moved first to the keyboard or went to the container before naming, they correctly indicated the name of the food item on 109 trials. However, across the 10 trials in which the chimpanzees erroneously went to the keyboard and named the food item without seeing the contents of the container, they correctly named the item only four times.
Experiment 2
Method
Subjects
The same chimpanzees were tested as in Experiment 1.
Design and procedure
The general design of Experiment 2 was similar to that of Experiment 1 in task requirements and in how the chimpanzees moved through space to make responses in naming the item in the container. The only difference concerned the actions of Experimenter 1.
In this experiment, Experimenter 1 first baited two opaque containers that were out of view of the chimpanzee with two preferred items, each of which had a corresponding lexigram. Both containers were covered and then placed on the floor in front of the chimpanzee. The first container was then lifted, the lid was removed, and either the contents were shown to the chimpanzee or the container was tilted away from the chimpanzee so that the contents remained unknown. The experimenter then repeated this process with the second container. On each trial, one container’s contents were revealed to the chimpanzee, and the other’s were not (counterbalanced for order across trials). The experimenter next moved one of the two containers to the second location, as in Experiment 1. This meant that on each trial, the chimpanzee saw one type of food in one container, but the contents of the other container remained unknown. The chimpanzee then had to attend to which container was moved to the second location so that, presumably, it would know whether it had seen the contents of that container or whether those contents remained unknown. Experimenter 1 left the test area, and trials proceeded exactly as they had in Experiment 1, with the chimpanzee moving to the second location and either looking in the container first or selecting a lexigram on the keyboard without having looked in the container.
Each session consisted of four trials, two in which the contents of the container moved to the second area had been revealed and two in which the contents of the container moved to the second area had not been revealed. The order of these two trial types was randomized within the session. Each chimpanzee completed five sessions in this experiment, for a total of 20 trials, 10 in each presentation condition (visible vs. nonvisible).
Results
All chimpanzees were significantly more likely to visit the keyboard and name first in the visible condition and visit the container first in the nonvisible condition— Sherman: χ2(1, N = 20) = 10.77, p = .001; Panzee: χ2(1, N = 20) = 5.05, p = .025; Lana: χ2(1, N = 20) = 16.36, p < .001 (Fig. 2).
For the first eight trials of Experiment 2, Lana and Sherman made the predicted response in seven of eight trials, and Panzee did so for five of eight trials. Thus, performance did not change across the course of either experiment, as would be expected if the chimpanzees had had to learn what to do through experience.
When the chimpanzees either saw the food item and moved first to the keyboard or went to the container before naming, they were correct in indicating the name of the food item on 55 of 56 trials. However, when they erroneously went to the keyboard and named without seeing the contents of the container, they correctly named the item only one time in four such trials.
General Discussion
In both experiments, the behavioral patterns of the chimpanzees were clear and consistent. In Experiment 1, when they saw the food early in the trial, they named it right away without looking again in the container. When they had not seen the item at the start of the trial, they approached the container, viewed its contents, and then went to the keyboard to name the food item. This pattern emerged in the earliest sessions; thus, the chimpanzees spontaneously reacted to the test with proficient responses on the basis of what information they had (or did not have) when they left their start position and moved to the response area.
As noted, it was possible that a simple behavioral rule could have underlain this performance, even in the early trials. Because these chimpanzees routinely request and name items (most often food items) during their daily interactions with humans, they may have learned over time that they should not name things they have not seen. An argument against this is that the chimpanzees most often use lexigrams with humans (mainly caregivers) to request foods that are not visible, so it is not uncommon for them to request food that is not present. However, given the novel requirement of having to move from one cage to another to name an item in a container, it is possible that the chimpanzees were behaving according to such a rule.
Experiment 2 discounted this possibility. In that experiment, the chimpanzees always saw a food item that they could name. However, that item was not always in the container that was moved to the response area, so they had to track and remember whether the container that was moved contained the item they could name or whether they needed to look into the container first. Again, they responded proficiently, even on the earliest trials. Thus, seeing food (or not seeing food) could not serve as a cue for which behavioral response to make, yet chimpanzees effectively sought information when it was needed and made a response immediately when information was already available.
These results indicate that chimpanzees know what they have seen and know when they can or cannot accurately name an item that is otherwise not visible at response time. This form of information-seeking behavior on trials in which the chimpanzees did not see what food was in the relevant container required them to leave the initial area, move to the container, and look in it—a relatively high-cost response, given that such movements more than doubled the time it took to name and obtain the item in the container. That the chimpanzees made this response only when they did not know what was in the container suggests that they recognized that they had no way to name the item without first viewing it, and then rectified that lack of information.
Our study made use of the special ability of these chimpanzees to use symbols that represent, in this case, food items to indicate what was in a container once they knew that information. Given their unique rearing histories, these chimpanzees’ ability to use symbols constitutes perhaps the strongest evidence for representational symbol use in any nonhuman animals (see Rumbaugh & Washburn, 2003), but the level of “language training” they underwent may not be requisite for giving nonhuman animals this kind of test. Our paradigm could be adapted for any species that had even a small number of associations between arbitrary symbols and real-world items, provided that those species were trained to indicate the “label” that went with each specific item once they could discern what that item was through their own information-seeking behaviors.
The outcome of this study supports previous arguments (e.g., Call, 2010; Call & Carpenter, 2001) that great apes, and perhaps other species, share with humans a metacognitive capacity for dealing with incomplete information. The information-seeking behavior of great apes is indicative of metacognition rather than learned rules for responding, although the metacognitive capacities of nonhuman animals are likely not equivalent to those reflected by the varied forms of information-seeking behaviors and other responses that humans make when they are doubtful or uncertain. That said, the existence of any metacognitive capacities in nonhuman animals has important implications for understanding the evolution of other aspects of “mind.” For example, there is an ongoing debate about the relative emergence of metacognition and of theory of mind in human evolution, with some arguing that metacognition likely emerged prior to theory of mind (e.g., Couchman, Coutinho, Beran, & Smith, 2009; Smith et al., 2008) and others arguing that the emergence of mind reading (theory of mind) was prior (e.g., Carruthers, 2008). Regardless of which view of the relative emergence of these capacities one takes, the present data suggest that the metacognitive processes that support information-seeking behavior are shared across species and that some species can engage controlled, decisional cognitive processes in the face of difficult problems for which only incomplete information is available.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by Grants HD-061455 and HD-060563 from the National Institute of Child Health and Human Development and by Grant BCS-0956993 from the National Science Foundation.