
Abstract
Despite the central role of learning in visual recognition, it is largely unknown whether visual form learning is maintained in older age. We examined whether training improved performance in both young and older adults at two key stages of visual recognition: integration of local elements and global form discrimination. We used a shape-discrimination task (concentric vs. radial patterns) in which young and older adults showed similar performance before training. Using a parametric stimulus space that allowed us to manipulate global features and background noise, we were able to distinguish integration and discrimination processes. We found that training improves global form discrimination in both young and older adults. However, learning to integrate local elements is impaired in older age, possibly because of reduced tolerance to external noise. These findings suggest that visual selection processes, rather than global feature representations, provide a fundamental limit for learning-dependent plasticity in the aging brain.
Successful visual recognition relies on the ability to extract structure from noisy sensory inputs, integrate local features into global forms, and discriminate among similar objects. Learning has been shown to play a key role in facilitating performance in a wide range of perceptual skills and in optimizing visual form recognition in young adults (for reviews, see Fine & Jacobs, 2002; Goldstone, 1998). In particular, previous work has shown that learning facilitates the detection and recognition of targets in clutter (e.g., Brady & Kersten, 2003; Dosher & Lu, 1998; Gold, Bennett, & Sekuler, 1999) by enhancing the integration of relevant features and their segmentation from noisy backgrounds.
Training has been shown to enhance perceptual skills not only in young but also in older adults, despite age-related decline of visual functions (Owsley, 2011). For instance, recent studies have shown that training enhances both young and older adults’ performance in brightness-discrimination (Ratcliff, Thapar, & McKoon, 2006), acuity (Fahle, 1993), texture-discrimination (Andersen, Ni, Bower, & Watanabe, 2010), and motion-direction-discrimination (Ball & Sekuler, 1986; Bower & Andersen, 2012) tasks. However, it remains largely unknown whether training facilitates the recognition of global forms beyond the processing of local features in older adults.
In the study reported here, we examined whether learning improves both young and older adults’ perceptual skills related to two key stages of visual recognition: the integration of local elements embedded in noise and the discrimination of global forms (Wilson & Wilkinson, 1998). To address this question, we designed a stimulus space and a model that allowed us to distinguish learning-dependent improvement in these two processes within the context of the same task. We used parametric manipulations of Glass patterns, which comprise oriented dot dipoles (Fig. 1a). For these stimuli, small local changes to the dot patterns have a predictable influence on the perception of global forms (Fig. 1a: concentric vs. radial patterns). These stimuli were ideally suited for our purpose, as previous studies had shown that performance in discriminating global forms in these patterns is similar (without training) in young and older adults (Habak, Wilkinson, & Wilson, 2009). Thus, we were able to compare learning between age groups while avoiding potentially confounding performance differences.
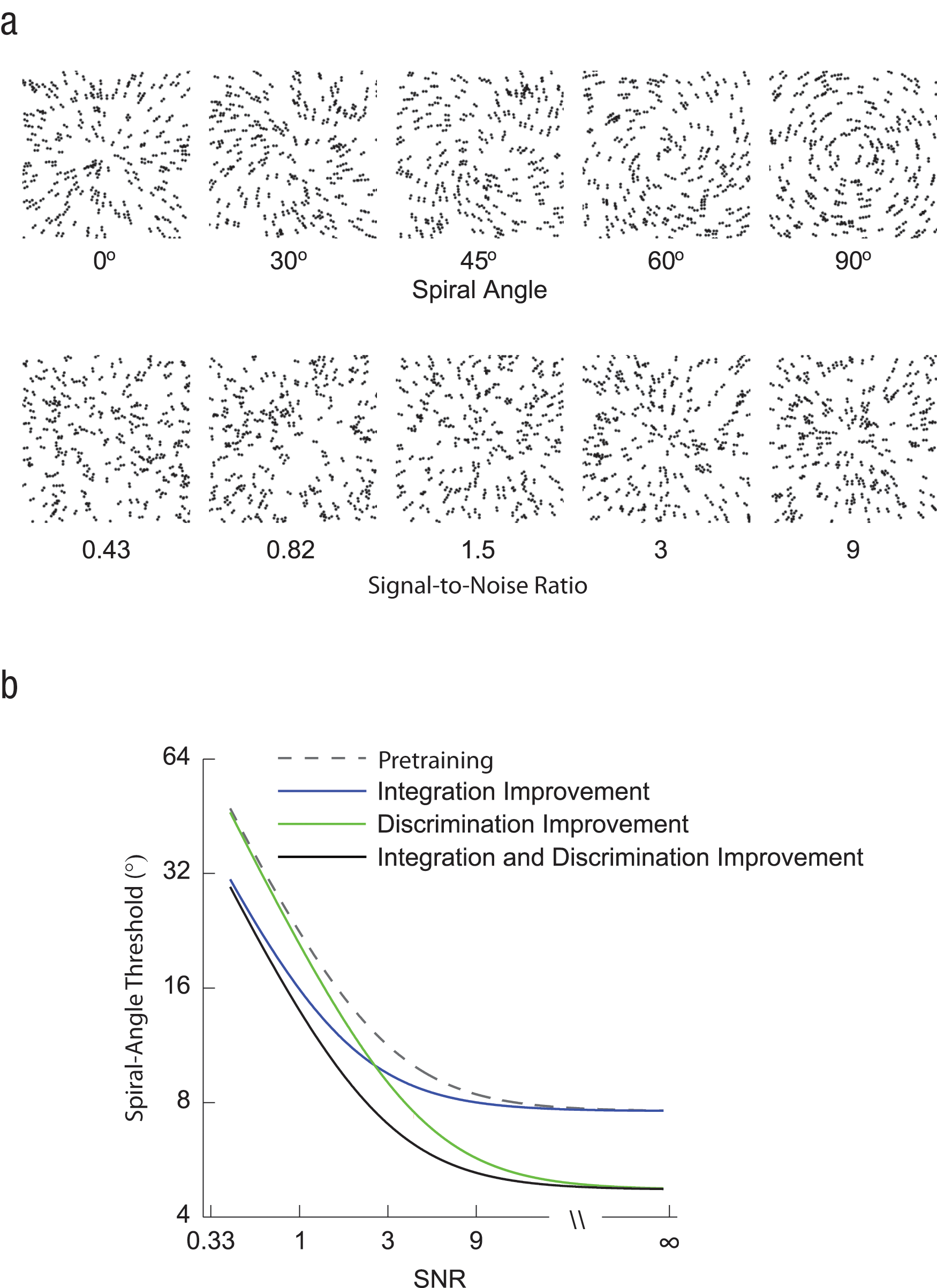
Examples of the Glass-pattern stimuli (a) and model predictions (b). In (a), the stimulus parameters have been adjusted for demonstration purposes (e.g., in the experiment, the stimuli comprised white dots presented against a black background). The top row shows Glass patterns with different spiral angles, ranging from 0° (radial pattern) to 90° (concentric pattern). The bottom row shows radial Glass patterns at different levels of signal-to-noise ratio (SNR; from 0.43 to 9). We introduce a model that distinguishes the separate contributions of local integration sensitivity and global form-discrimination sensitivity to performance improvement in posttraining relative to pretraining. The graph illustrates how improvement in these processes might relate (separately and in combination) to changes in spiral-angle threshold, across SNR levels.
Moreover, the parametric nature of the stimuli enabled us to distinguish processes of local integration and global form discrimination. In particular, we manipulated (a) the amount of background noise (i.e., randomly oriented dipoles) and (b) the similarity between global forms, using linear morphing between concentric and radial patterns. Measuring performance when background noise was at a high level enabled us to assess sensitivity in integrating local orientations (i.e., extracting from noise and grouping signal dipoles that belonged to the global form patterns), whereas measuring performance when background noise was at a low level enabled us to uncover sensitivity to global configurations (i.e., sensitivity to differences in curvature between concentric and radial patterns). We developed a model that allowed us to distinguish local integration and global form-discrimination processes (Fig. 1b) and tested how training changes behavioral performance related to these processes in young and older adults.
Our findings demonstrate a dissociation in the ability to learn visual forms in older adults. In particular, learning to discriminate global form differences is maintained, whereas learning to extract local elements from noise and to integrate these elements is impaired. These results provide novel insights into learning-dependent plasticity in older age. In particular, we propose that visual selection processes, rather than global feature representations, provide the fundamental limit for learning-dependent plasticity in the aging brain.
Method
Observers
Twenty-four healthy young adults (median age = 21 years, range = 18–26) and 24 older adults (median age = 72 years, range = 62–81) participated in the study. All observers were naive to the purpose of the experiments, had normal or corrected-to-normal vision (20/20 performance), and gave written informed consent. We measured (after optical corrections) far (Snellen chart) and near (Landolt C) visual acuity. All older adults scored within the range of normal cognitive ability (median = 30, range = 26–30) in the Mini-Mental State Examination (Folstein, Robins, & Helzer, 1983). The study was approved by the University of Birmingham Ethics Committee.
Stimuli
We used Glass-pattern stimuli defined by 600 pairs of white dots (dipoles) displayed within a square aperture (7.5º × 7.5º) on a black background (Fig. 1a). Each dipole comprised two 0.0375° dots separated by 0.26°. These parameters were chosen on the basis of pilot psychophysical studies and in accordance with previous work (Wilson & Wilkinson, 1998) showing that coherent form patterns are reliably perceived in stimuli with these parameters. We generated concentric and radial Glass patterns by placing dipoles tangentially (concentric stimuli) or orthogonally (radial stimuli) to the circumference of a circle centered on the fixation dot. Further, we generated patterns intermediate between these two Glass-pattern types by parametrically varying the spiral angle of the pattern between 0º (radial pattern) and 90º (concentric pattern). For each dot dipole, the spiral angle was defined as the angle between the dot dipole’s orientation and the radius from the center of the dipole to the center of the stimulus aperture. We also manipulated signal-to-noise ratio (SNR; i.e., the ratio of signal dipoles to noise dipoles, which were randomly positioned and oriented), presenting stimuli at 30%, 45%, 60%, 75%, 90%, and 100% signal—corresponding to SNRs of 0.43, 0.82, 1.5, 3, 9, and ∞, respectively (Fig. 1a). We set the lowest SNR at the detection threshold of Glass patterns in noise (29.8 ± 1.59% signal) indicated by our pilot studies. A new pattern was generated for each trial; consequently, stimuli were locally jittered in their position.
Procedure
All observers were familiarized with the task and stimuli during a short practice session (100 trials). Observers participated in one pretraining and one posttraining session without feedback and four to five training sessions with auditory feedback. On each trial, a stimulus pattern was presented for 200 ms, followed by a 500-ms blank screen. Observers were instructed to report whether the pattern was radial or concentric. We measured observers’ performance using a 3-down/1-up staircase method, which resulted in 79.4% convergence level. We measured spiral-angle thresholds by averaging the spiral angles at the last two thirds of reversals in each staircase. In the pre- and posttraining sessions, we measured participants’ performance using three to five staircases with 9 or 10 up-down reversals at each SNR. In each training session, participants received training on 1,200 to 2,000 trials; participants in Experiment 1 were trained with all six SNR levels (200–300 trials per SNR), and participants in Experiment 2 were trained with an SNR of 0.82 or 9.
Data analysis
To distinguish local integration and global form-discrimination processes, we developed a two-process model to fit spiral-angle discrimination thresholds across SNR levels (Fig. 1b). We reasoned that thresholds (S th) are determined by the signal required to (a) extract and integrate local dipoles (Si ) and (b) discriminate fine shape differences (Sd ):

To normalize the shape-discrimination thresholds (to a range from 0 to 1), we divided the measured thresholds  by the reference spiral angle (45°):
by the reference spiral angle (45°):

We assumed that Si
is inversely related to SNR. Therefore, Si
can be written as
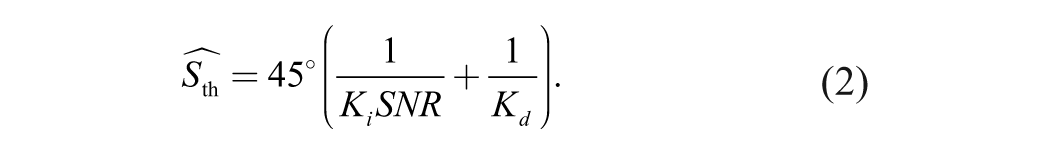
Further, to evaluate the speed of learning, we used a single-parameter model to fit thresholds across sessions:
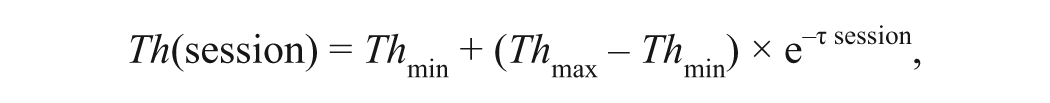
where Th(session) is the mean shape-discrimination threshold across sessions, Th max and Th min are the highest and lowest thresholds, and τ represents the learning rate (LR).
In addition, we quantified the effect of training using the following two indices:
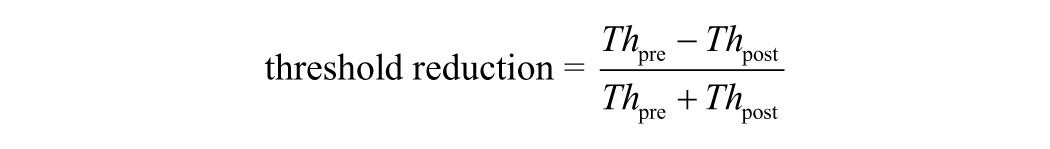
and

where Th pre and K pre are threshold and sensitivity in pretraining, and Th post and K post are threshold and sensitivity in posttraining.
We conducted repeated measures analyses of variance (ANOVAs; Greenhouse-Geisser corrected) on mean performance thresholds, threshold reduction, and sensitivity improvement to assess differences in visual form learning in young and older adults.
Results
Experiment 1: training with stimuli across multiple signal levels
In the first experiment, participants (8 young and 8 older adults) were trained to discriminate concentric and radial Glass patterns presented at six different SNR levels (0.43, 0.82, 1.5, 3, 9, and ∞). We found that training improved performance (i.e., reduced thresholds across sessions) for both young and older adults (Fig. 2a). However, older adults showed less improvement in local integration than in global form-discrimination sensitivity.
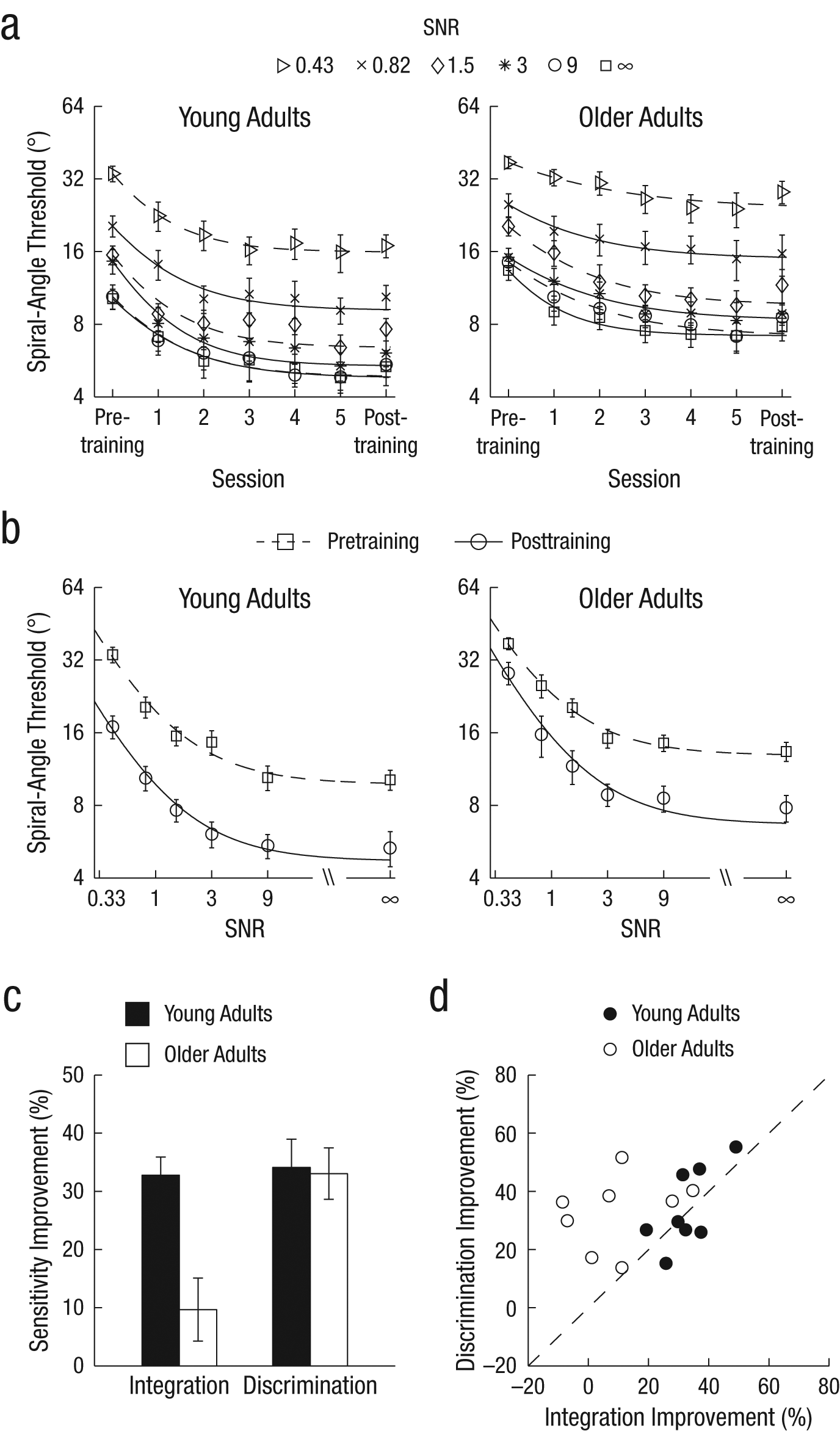
Results from Experiment 1: (a) spiral-angle threshold as a function of session for young (left) and older (right) participants at each signal-to-noise ratio (SNR), (b) discrimination threshold as a function of SNR for young (left) and older (right) participants before and after training, (c) percentage improvement in local integration and global form-discrimination sensitivity after training for young and older participants, and (d) improvement in discrimination sensitivity as a function of improvement in integration sensitivity for individual participants. The thresholds in (b) were fitted with Equation 2 (R = .97). In all plots, error bars indicate standard errors of the mean across participants.
Specifically, to determine how training changed local integration and global form-discrimination processes, we fit a two-process model (Fig. 1b) to the pre- and posttraining thresholds obtained at different SNR levels for both young and older adults (Fig. 2b). This model showed that young adults improved in both local integration and form discrimination, whereas older adults improved mainly in global form discrimination. That is, young adults’ performance improved across all SNR levels, whereas older adults’ performance improved more at high than at low SNR levels, as indicated by a significant interaction between session (pre- vs. posttraining), age (young vs. older participants), and SNR (six levels; repeated measures ANOVA on thresholds), F(2.9, 40.9) = 4.97, p = .005.
Using the model fits, we compared improvement in integration and discrimination sensitivity in young and older adults (Figs. 2c and 2d). Older adults showed similar improvement in global form discrimination but weaker improvement in local integration compared with young adults. A repeated measures ANOVA showed a significant interaction of session (pre- vs. posttraining), age (young vs. older participants), and process (integration vs. discrimination), F(1, 14) = 5.14, p = .04. Young and older participants showed similar improvement in discrimination sensitivity (i.e., young adults: range = 15.2% to 55.2%; older adults: range = 13.7% to 51.7%), as indicated by the lack of a significant interaction between session and age, F(1, 14) = 1.17, p = .298. However, older adults showed weaker improvement in local integration (less than 10% improvement for 4 of 8 participants) than young adults did, as indicated by a significant interaction between session and age, F(1, 14) = 12.96, p = .003.
Experiment 2: training with stimuli at high versus low SNR
Is it possible that older adults’ weaker improvement in local integration relative to young adults was due to limited training with low-SNR stimuli in Experiment 1? To test this possibility, in Experiment 2 we trained each participant at one SNR level for 1,200 to 2,000 trials per session, rather than at each SNR level for 200 to 300 trials per session. First, we trained young and older participants (8 per group) to discriminate concentric and radial Glass patterns at a low SNR level. We chose the SNR of 0.82, to avoid floor effects (i.e., training at very high noise levels may be too difficult to show performance improvement). We then trained different groups of young and older participants (8 per group) at a high SNR level. We used the SNR of 9, as improvements appeared to saturate at this level in Experiment 1 (Fig. 2b).
We reasoned that training at a low SNR would facilitate the ability to extract and integrate local elements, whereas training at a high SNR would facilitate discrimination of global form differences. We measured discrimination thresholds in the pretraining, posttraining, and training sessions and tested for learning-dependent improvement at the trained SNR and transfer of learning to untrained SNR levels.
Training at the low SNR (0.82) resulted in weaker improvement at that SNR for older than for young adults (Fig. 3a), as indicated by a significant interaction between session and age (repeated measures ANOVA on performance thresholds), F(1, 14) = 7.23, p = .018. This result is consistent with Experiment 1, demonstrating weaker improvement in local integration sensitivity for older than for young adults. Moreover, comparing the learning rate at the trained SNR between age groups (Fig. 3b) showed that older adults learned more slowly than young adults (young adults: mean LR = 1.03, SD = 0.15; older adults: mean LR = 0.42, SD = 0.06), t(14) = 3.88, p = .002.

Results for participants trained at the low signal-to-noise ratio (SNR; 0.82) in Experiment 2: (a) pre- and posttraining spiral-angle thresholds as a function of SNR for young (left) and older (right) participants, (b) thresholds at the trained SNR for young and older adults as a function of session, (c) percentage improvement in local integration and global form-discrimination sensitivity after training for young and older participants, and (d) improvement in discrimination sensitivity as a function of improvement in integration sensitivity for individual participants. The arrows in (a) indicate the SNR used for training. The thresholds in (b) were fitted with Equation 2 (R = .96). In all plots, error bars indicate standard errors of the mean across participants.
We used our two-process model to compute sensitivity improvement across SNRs in young and older adults trained at the low SNR. We found that training at this SNR enhanced local integration sensitivity more in young than in older adults (Figs. 3c and 3d). Integration improvement ranged from 4.8% to 52.6% for young participants but from −8.8% to 33.2% for older participants, with 1 young and 3 older participants showing improvement lower than 10%. Performance improvement generalized to global form-discrimination sensitivity for both young adults (range: −0.5% to 50.8%) and older adults (range: −2.9% to 30.1%), with only 1 young and 1 older participant showing improvement lower than 10%. A repeated measures ANOVA on sensitivity improvement confirmed this result, showing significant differences between age groups, F(1, 14) = 10.99, p = .005, but no significant effect of process, F(1, 14) = 0.3, p = .592, and no significant interaction between age and process, F(1, 14) = 0.15, p = .703. Further, performance improvement transferred from the trained (0.82) to the untrained SNR levels in both young and older adults (Fig. 3a). Comparing threshold reduction across SNRs revealed no significant effect of SNR, F(5, 70) = 1.07, p = .383, and no significant interaction between SNR and age, F(5, 70) = 0.82, p = .543.
The participants who were trained at the high SNR level (9) showed a different pattern of results. Specifically, we found similar performance improvement for young and older adults (Fig. 4a). A repeated measures ANOVA on performance thresholds at the trained SNR showed a significant effect of session, F(1, 14) = 123.59, p < .001, but no significant difference between age groups, F(1, 14) = 1.11, p = .31. Further, there was no significant difference in the learning rate at the trained SNR (Fig. 4b) between young and older adults (young adults: mean LR = 0.77, SD = 0.14; older adults: mean LR = 0.68, SD = 0.14), t(14) = 0.43, p = .677.

Results for participants trained at the high signal-to-noise ratio (SNR; 9) in Experiment 2: (a) pre- and posttraining spiral-angle thresholds as a function of SNR for young (left) and older (right) participants, (b) thresholds at the trained SNR for young and older adults as a function of session, (c) percentage improvement in local integration and global form-discrimination sensitivity after training for young and older participants, and (d) improvement in discrimination sensitivity as a function of improvement in integration sensitivity for individual participants. The arrows in (a) indicate the SNR used for training. Thresholds are fitted with Equation 2 (R = .96). In all plots, error bars indicate standard errors of the mean across participants.
We used our two-process model to compute sensitivity improvement in young and older adults trained at the high SNR. We found that training at this SNR enhanced global form-discrimination sensitivity similarly in young adults (range: 4.9% to 48.2%) and older adults (range: 6.1% to 47%), with only 1 young and 1 older adult showing improvement lower than 10% (Figs. 4c and 4d). However, improvement in global form discrimination did not appear to generalize to local integration sensitivity for either young adults (range: −8.8% to 55.5%) or older adults (range: −25.3% to 15.5%), with 5 young and 4 older participants showing improvement lower than 10%. A repeated measures ANOVA on sensitivity improvement showed stronger improvement in form discrimination than in integration sensitivity, F(1, 14) = 17.91, p = .001, but no significant interaction between process and age, F(1, 14) = 0.38, p = .548. This lack of learning transfer to local integration sensitivity is also evident in Figure 4a, which shows weak transfer from the trained SNR (9) to lower, untrained SNR levels in both young and older adults; age and SNR did not have a significant interactive effect on performance threshold, F(5, 70) = 0.68, p = .641. Further, comparing threshold reduction across SNRs revealed a significant effect of SNR, F(5, 70) = 5.24, p < .001. Threshold reduction was significantly smaller at the SNR of 0.43 compared with other SNR levels, F(1, 14) = 15.26, p = .002, which suggests weak transfer of learning from a high to low SNRs.
In sum, comparing training at low and high SNRs corroborated our findings from Experiment 1: Compared with young adults, older adults showed weaker improvement in local integration but similar improvement in global form discrimination. Comparison of the results of the two experiments indicated that older adults’ weaker improvement in local integration sensitivity could not have been due to limited training. In particular, more training at the low SNR in Experiment 2 did not result in enhanced local integration sensitivity for older adults in that experiment compared with Experiment 1. At the low SNR only (0.82), there was no significant difference between experiments in older adults’ performance threshold, F(1, 14) = 2.66, p = .125, or local integration sensitivity, F(1, 14) = 0.24, p = .634. Interestingly, we found an asymmetry in the transfer of learning between trained and untrained SNR levels. Training at the low SNR improved not only local integration but also global form-discrimination sensitivity in both young and older adults. In contrast, training at the high SNR yielded improvement that was more specific to global form-discrimination sensitivity.
Could the differences in improvement between age groups have been due to differences in performance before training? Our experiments are consistent with recent studies showing that shape discrimination is preserved in older age (Habak et al., 2009; Wang, 2001), as we observed similar pretraining performance across SNRs for young and older adults (Fig. 5a). In particular, repeated measures ANOVAs on pretraining thresholds showed neither significant main effects of age—Experiment 1: F(1, 14) = 2.67, p = .124; Experiment 2, low-SNR training: F(1, 14) = 0.01, p = .907; Experiment 2, high-SNR training: F(1, 14) = 3, p = .105—nor significant interactions between age and SNR—Experiment 1: F(2.7, 37.7) = 1.22, p = .314; Experiment 2, low-SNR training: F(2.5, 35.4) = 0.79, p = .488; Experiment 2, high-SNR training: F(2.5, 34.5) = 0.16, p = .89.

Pretraining performance in Experiments 1 and 2 combined: (a) spiral-angle threshold as a function of signal-to-noise ratio (SNR) for young and older participants and (b) mean local integration sensitivity and global form-discrimination sensitivity for young and older adults. Error bars indicate standard errors of the mean across participants.
Further, we analyzed behavioral sensitivity before training in the two experiments, and found that local integration and global form discrimination were similar for young and older adults (Fig. 5b). Repeated measures ANOVAs showed no significant main effects of age—Experiment 1: F(1, 14) = 1.77, p = .205; Experiment 2, low-SNR training: F(1, 14) = 0.01, p = .92; Experiment 2, high-SNR training: F(1, 14) = 2.4, p = .143—nor significant interactions between age and process—Experiment 1: F(1, 14) = 2.45, p = .14; Experiment 2, low-SNR training: F(1, 14) = 0.79, p = .389; Experiment 2, high-SNR training: F(1, 14) = 0.08, p = .781. These results suggest that the posttraining performance differences between age groups reflect differences in learning ability rather than processing capacity before training.
Discussion
Our study demonstrates a dissociation in visual form learning in older adults: Learning to discriminate global form differences is maintained, whereas learning to extract and integrate local elements is impaired. Our methodological approach has two main advantages. First, using a shape-discrimination task in which young and older participants showed similar pretraining performance allowed us to compare behavioral improvement between young and older adults without confounding differences between age groups before training. Second, we developed a parametric stimulus space and a two-process model that allowed us to distinguish learning-dependent improvement in local integration and global form-discrimination processes. Our model is consistent with previous studies (Kurki, Laurinen, Peromaa, & Saarinen, 2003; Wilson & Wilkinson, 1998) in assuming two processes, one that involves extracting signal from random elements by grouping elements of similar orientation, and another that identifies fine differences between global configurations (e.g., differences in curvature between concentric and radial patterns).
An alternative approach suggests that Glass-pattern discrimination could be explained by a single process (Dakin, 1997) that computes the mean local orientation across a given number of dipoles. This model is consistent with our prediction for improvement in both local integration and global form discrimination as a result of learning (Fig. 1b) and with the results we observed for young participants when they were trained across all SNR levels. However, this model does not predict greater improvement in global form discrimination than in local integration (i.e., greater improvement at high than at low SNRs), as we observed in older participants and young participants trained at a high SNR only. Thus, our results suggest that a two-process model is more appropriate for understanding the limits (i.e., reduced tolerance to external noise) of learning-dependent improvement in older adults.
Our results advance understanding of learning-dependent plasticity in older age in three main ways. First, previous studies investigating visual learning in older adults have focused on visual feature (e.g., texture, motion) learning. These studies have shown similar learning-dependent improvements for young and older adults (Andersen et al., 2010; Bower & Andersen, 2012). Here, we have provided evidence that learning molds not only local feature processing but also global form perception in older adults. Previous studies investigating age-related differences in the visual recognition of global forms (Del Viva & Agostini, 2007; Habak et al., 2009; McKendrick, Weymouth, & Battista, 2010; Roudaia, Bennett, & Sekuler, 2008; Roudaia, Farber, Bennett, & Sekuler, 2011) have not distinguished local integration and global form- discrimination processes. Here, we have demonstrated that young and older adults differ in their ability for local integration but not global form-discrimination learning.
Second, previous studies have suggested that aging may result in reduced efficiency (Bennett, Sekuler, & Ozin, 1999), reduced tolerance to external noise (Bower & Andersen, 2012), or increased internal noise (Bennett, Sekuler, & Sekuler, 2007). However, it has been suggested that learning in young adults enhances performance efficiency (Gold, Sekuler, & Bennett, 2004), improves exclusion of external noise, and reduces internal noise (Dosher & Lu, 1999). In our study, we could not discriminate the effects of internal and external noise, as we measured threshold for a stimulus dimension (spiral angle) different from the noise dimension (signal vs. noise dot dipoles). However, it is likely that the high external noise used when measuring local integration sensitivity resulted in reduced learning-dependent improvement in older adults. This suggestion is consistent with previous studies showing that the ability to integrate contours in clutter (Del Viva & Agostini, 2007; Roudaia et al., 2008) and suppress the background (Betts, Sekuler, & Bennett, 2009; Betts, Taylor, Sekuler, & Bennett, 2005) deteriorates in older age. It is possible that this deterioration is due to weakening of inhibitory processes (Hua et al., 2010; Leventhal, Wang, Pu, Zhou, & Ma, 2003) or attentional functions (Ball, Roenker, & Bruni, 1990; Kane, Hasher, Stoltzfus, Zacks, & Connelly, 1994) that mediate the ability to suppress irrelevant information and enhance the salience of behaviorally relevant targets in cluttered scenes. In light of these previous studies, our findings suggest that visual form learning is limited by visual selection processes in aging. Although our results did not show any significant differences in Glass-pattern discrimination between young and older adults before training, it is possible that age-related decline in visual selection processes hinders the ability of training to improve extraction of local elements from noise and integration of those elements.
Third, the hallmark of perceptual learning has been that improvement is specific to trained features (Ball & Sekuler, 1982; Fahle, 2004; Karni & Sagi, 1991). However, previous studies have shown that transfer of orientation training can depend on the difficulty of the training (Dosher & Lu, 2005; Liu & Weinshall, 2000) or the transfer (Jeter, Dosher, Petrov, & Lu, 2009) task. Transfer of learning appears to be stronger between two easy conditions or from a hard to an easy condition than between two difficult conditions or from an easy to a hard condition. Those results may appear at odds with our results showing stronger transfer of shape training from local integration (low SNR) to global form discrimination (high SNR) than vice versa. These differences between studies could be due to the different stimulus paradigms used, as previous studies did not train sensitivity for local integration and for global form discrimination separately. The learning transfer from local integration to global form-discrimination sensitivity that we observed may relate to transfer from an easy to a difficult task based on top-down processing mechanisms (Ahissar & Hochstein, 1993). In particular, global forms are easy to discriminate once detected at a low SNR. Global form features extracted during training at a low SNR may then guide training in the more difficult fine-feature discrimination at a high SNR.
In sum, our findings suggest that visual form learning in older adults is limited by visual selection processes rather than by fine feature processing. It has been suggested that processes related to the integration and detection of global patterns (e.g., texture, forms) develop late (Braddick & Atkinson, 2007; Kiorpes & Bassin, 2003; Kovacs, 2000). This may relate to late maturation of cortical connections, possibly lateral connections within primary visual cortex or feedback connections from parietal circuits involved in attentional salience and target detection. It is possible that these circuits decline earlier in older age, and thus provide a fundamental limit to learning-dependent visual plasticity. Future neuroimaging and physiology studies investigating these circuits are needed to test how developmental changes in brain connectivity determine the ability for visual learning and successful object recognition across the life span.
Footnotes
Acknowledgements
We are grateful to Aimee Goldstone, Jessica Sandall, and Hayley Wolstencroft for help with the data collection and to Dorita Chang, Lynne Kiorpes, and Andrew Welchman for helpful comments.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by grants to Z. K. from the Biotechnology and Biological Sciences Research Council (D52199X, E027436) and the European Community’s Seventh Framework Programme (FP7/2007-2013) under Agreements 255577 and 214728.