
Abstract
Facial expressions may have originated from a primitive sensory regulatory function that was then co-opted and further shaped for the purposes of social utility. In the research reported here, we tested such a hypothesis by investigating the functional origins of fear expressions for both the expresser and the observer. We first found that fear-based eye widening enhanced target discrimination in the available visual periphery of the expresser by 9.4%. We then found that fear-based eye widening enhanced observers’ discrimination of expressers’ gaze direction and facilitated observers’ responses when locating eccentric targets. We present evidence that this benefit was driven by neither the perceived emotion nor attention but, rather, by an enhanced physical signal originating from greater exposure of the iris and sclera. These results highlight the coevolution of sensory and social regulatory functions of emotional expressions by showing that eye widening serves to enhance processing of important environmental events in the visual fields of both expresser and observer.
Darwin (1872/1998) theorized that facial expressions originated from a direct egocentric benefit to the expresser, whose signals then served as an indirect allocentric benefit to the observer. One salient feature of facial-expression appearance, the widening of the eyes (e.g., raised eyebrows and opened eyelids in expressions of fear; Ekman, Friesen, & Hager, 2002), may have been shaped to directly modulate sensory intake (Susskind et al., 2008). This egocentric sensory benefit may have originated in fear’s service of vigilance (Darwin, 1872/1998), in the face of uncertainty in the environment (Whalen, 1998), to gather immediate information about potential threats. The function of eye widening in fear expressions also extends beyond the self by serving as a powerful social signal (Marsh, Adams, & Kleck, 2005; Whalen et al., 2004). This function likely is facilitated by the additional contrast of white sclera in humans, which is unique among primates and is thought to have coevolved with humans’ social nature (Kobayashi & Kohshima, 1997). Indeed, the eyes alone can convey rich information about people’s innermost mental states (Baron-Cohen, Wheelwright, Hill, Raste, & Plumb, 2001), as well as signal where to direct attention through eye gazes (Friesen & Kingstone, 1998).
Prior work has examined the interaction between fear and eye gazes, showing that fear expressions facilitate observers’ judgments of averted versus direct gaze in expressers (Adams & Franklin, 2009) and, inversely, that averted gaze in fear expressers enhances the intensity of fear perceived by observers (Adams & Kleck, 2005). Directional eye gazes in fear expressions have also been shown to deploy additional attention in the context of an attentional-cuing paradigm (Putman, Hermans, & van Honk, 2006; Tipples, 2006). Although these effects illustrate how fear expressions benefit the observer by evoking a congruent state of vigilance, it is important to note that they are hinged to the communicated emotion, the state of alarm that may reverberate in the observer to act as the catalyst for the benefits (e.g., Harrison, Singer, Rotshtein, Dolan, & Critchley, 2006).
In the present study, we asked whether the benefits of fear expressions are transmitted interpersonally at a more basic, physical level. The retraction of the eyelids and eyebrows in fear expressions has likely resulted from multiple selective pressures, only one of which may have been the need to communicate a particular emotion or mental state (Shariff & Tracy, 2011; Susskind et al., 2008). Our thesis, which we examined in the study reported here, was that these actions may have coevolved to enhance processing of events in the expresser’s visual field while simultaneously augmenting the expresser’s gaze signal to aid the observer in locating the same events. At a physical level, the conjunction of the increased iris-to-sclera contrast of the human eye and the eye’s widening to enhance that contrast may serve as the most expedient social signal of a significant event’s location. Thus, the potential personal sensory benefit of eye widening would be directly conferred interpersonally, prior to and without the need for the communicated emotion of the expresser.
To examine our thesis, we first tested the personal benefit of eye widening in fear expressions, hypothesizing that it would enhance visual encoding of the expresser’s surroundings, as reflected by the expresser’s discrimination of stimuli farther out in the periphery. Although our prior study showed evidence that fear-based eye widening enhanced detection of simple spots of light (Susskind et al., 2008), the present study provided a more rigorous test of the effect of fear expressions on discrimination in the eccentric visual field. We then tested the interpersonal benefit of fear-based eye widening at a basic signal level, in the absence of social perception (i.e., communication) of fear. Specifically, we examined whether the observer’s discrimination of the expresser’s gaze direction would be enhanced. We predicted not only that fear would expand the expresser’s sensitive visual field but also that the functional essence of this benefit would be passed on to the observer through transmission of a clearer “look-here” gaze signal.
Fear signals, however, play an important role in attracting attention (Vuilleumier, Armony, Driver, & Dolan, 2001). To limit the influence of the perceived emotion from the physical signal of the eyes without altering their physical form, we created simple schematic-outline stimuli of just the eyes extracted from images depicting full fear expressions. We further examined these eyes’ effects when inverted, hypothesizing that perceived fear would be reduced (McKelvie, 1995) but that the enhanced gaze signal would be maintained. Such a result would provide support for the notion that the physical signal rather than the perception of fear is responsible for the enhanced gaze signal. Finally, we used these eye-gaze stimuli in an attentional-cuing paradigm to test whether gaze-signal enhancement facilitated participants’ location of peripheral targets and to simultaneously examine their capacity to alter covert attention. In all experiments, we contrasted the eyes of expressive fear with the eyes of expressive disgust and expressionless, neutral eyes. Disgust was selected as an emotional expression because it has the narrowest eye aperture, in opposition to fear, which has the widest aperture; thus, we spanned the largest possible variance in a continuum of expressive eye opening (Susskind et al., 2008).
Experiment 1: Personal Sensory Benefit of Fear’s Eye Widening
To test how eye widening in fear enhances the visual encoding of the expresser’s surroundings, we had participants pose fear, neutral, and disgust expressions while they judged Gabor-grating orientations in their visual periphery. We hypothesized that fear expressions, compared with neutral or disgust expressions, would enhance discrimination farther out in the periphery.
Method
Twenty-eight undergraduates provided informed consent and participated in return for $10. All had normal or corrected-to-normal vision. To access the visual periphery, a 1,280 × 1,024-pixel-resolution image from a personal computer using E-Prime Version 1.1 (Psychology Software Tools, Sharpsburg, PA) was projected to a size of 160 × 120 cm. Participants were seated and stabilized via chin rest 30 cm from the projection screen. Participants fixated on a 1-cm central cross at eye level.
During the posed expression, observers responded to a target Gabor grating (sinusoidal grating enveloped by a Gaussian function at 0.5 cycles per degree, full contrast; created using MATLAB Version 7.04; The MathWorks, Natick, MA) that appeared along one of three meridians (one vertical and two oblique; see Fig. 1a). The Gabors appeared at varying eccentricities from fixation, depending on calibration and practice phases, and always subtended 7.8° (the correct pixel-sized Gabor was selected based on eccentricity).

Time course of trials (a), stimuli (b), and results (c, d) of Experiment 1. While maintaining fixation at the bottom of a projected image, participants posed and held a fear, neutral, or disgust expression. After a delay (randomized duration of 700 to 1,100 ms), they judged the orientations of Gabor gratings (the 0° orientation is shown) presented along one of three meridians (the dashed lines showing these three meridians did not appear in the experiment). At target onset, a 400-ms tone was played over desktop speakers (500 Hz sinusoidal) because the target may not have been visible. As feedback, a bar (green for a correct response or red for an incorrect response) was displayed at eye level across the bottom of the screen. The four possible orientations of the Gabor stimuli (clockwise from top left: 0°, 45°, 90°, and 135°) are shown in (b). The graph in (c) shows results for the vertical meridian for 1 participant. The percentage of correct responses is plotted as a function of eccentricity of the Gabor stimulus for each of the three expressions; modeled logistic regression curves are also shown. For each combination of expression and meridian, each participant’s accuracy was computed at every eccentricity, and these values were then submitted to a separate logistic regression analysis with a single predictor (eccentricity) and a constant term. The lower bound was set to chance (25%). Psychometric threshold eccentricities (62.5%; marked by the dashed horizontal line in the graph) were computed for each logistic regression function. The rightward shift of the regression curves from disgust to neutral to fear indicates that successful discrimination extended farther out in the periphery. The bar graph (d) shows the mean psychometric threshold eccentricities for each of the three expressions. Error bars represent standard errors of the mean.
Experiment 1 employed a 3 (expression) × 3 (meridian) × 6 (eccentricity) within-subjects factorial design. Each trial (Fig. 1a) began with a delay of 700 to 1,100 ms (randomized in 100-ms increments), followed by the presentation of the target for up to 4,000 ms or until response. Participants responded to one of four possible Gabor orientations (Fig. 1b). Each trial ended with a 200-ms presentation of a green or red bar at eye level on the screen as feedback indicating a correct response or an incorrect/missed response, respectively (for additional details, see Experiment 1 in the Supplemental Material available online).
Results
The threshold eccentricities for each participant’s expression (see Fig. 1c) were submitted to a 3 (expression) × 3 (meridian) repeated measures analysis of variance (ANOVA). As hypothesized, there was a main effect of expression, F(2, 36) = 39.7, p < .001, η p 2 = .69 (Fig. 1d), such that participants discriminated the Gabor-grating orientations at farther eccentricities when posing fear expressions compared with neutral expressions, F(1, 18) = 9.0, p < .01, η p 2 = .33, and disgust expressions, F(1, 18) = 69.6, p < .001, η p 2 = .80. It is important to note not only that eye closure in disgust resulted in the expected visual-field reduction associated with its putative defensive function (Chapman, Kim, Susskind, & Anderson, 2009; Susskind et al., 2008) but also that fear expressions increased the effective visual field by 9.4% into the available periphery (maximum of 90°) compared with neutral expressions.
Experiment 2a: Interpersonal Benefit of Eye Widening in Fear Expressions
Because the expressers in Experiment 1 received the sensory benefit of discriminating stimuli farther out in the periphery, we next tested the signaling benefits that may be transmitted to onlookers of fear expressions. In Experiment 2a, participants discriminated the direction of eye gazes across four eye sizes ranging from the narrowest, associated with disgust, to the widest, associated with fear. We hypothesized that wider eye gazes would be more accurately discriminated because they would provide a stronger “look-here” signal. To limit the influence of the full emotional expression, we used impoverished schematic eye stimuli that did not show the rest of the face. The eye shapes were created using a statistical model (Cootes, Edwards, & Taylor, 2001) of posed fear and disgust expressions from 19 individuals (Susskind et al., 2008). Using the model, we computed average fear eyes (Size 4) and disgust eyes (Size 1), along with eyes of two intermediate sizes (Sizes 2 and 3; see Fig. 2b).

Time course of trials (a), stimuli (b), and results (c, d) of Experiments 2a and 2b. A trial began with fixation (randomized duration of 1,000 to 1,400 ms). An eye-gaze stimulus then appeared for 300 ms, after which it was masked. The rectangular mask appeared for up to 2,000 ms or until the participant responded to indicate the eyes’ gaze direction. A red square appeared for 200 ms as feedback if the participant failed to respond. Stimuli consisted of schematic drawings of differently sized eyes (b), which were modeled using participants who were instructed to pose disgust expressions (top images; Size 1) and fear expressions (bottom images; Size 4). Intermediate Sizes 2 and 3 were interpolated linearly from Size 1 to Size 4 in equal steps of vertical aperture. Eyes in the right column are inverted versions of eyes in the left column and were used only in Experiment 2b. All degrees of gaze shown are 0.063° to the left of center. The graph in (c) shows the pattern of correct gaze-discrimination responses as a function of degree of gaze (L = left of center, R = right of center) and eye size in Experiment 2a (each circle represents the average of all participants across all trials at a given degree of gaze); modeled logistic regression curves are also shown. The steeper logistic regression slope for fear eyes compared with disgust eyes indicates greater gaze-direction discriminability. The graph in (d) shows a plot of mean maximum logistic regression slopes computed using separate logistic regression models for each eye size for each participant. Slope values were standardized for the upright and inverted conditions within each experiment (“Exp.”). Close overlap across experiments indicates replicated effects of increased discrimination with increased eye size. Error bars represent standard errors of the mean.
Method
Twenty-six undergraduates who had not taken part in the previous experiment provided informed consent and participated in return for $10 or course credit. All had normal or corrected-to-normal vision. Participants were seated and stabilized via chin rest 40 cm from an LCD monitor with a display resolution of 1,280 × 1,024 pixels (60 Hz). Like Experiment 1, Experiment 2a was run from a personal computer using E-Prime software (Version 1.1).
Participants fixated on a 0.2° central cross and responded to a target pair of eyes subtending 7.2° × 0.47° (smallest) to 7.2° × 1.0° (largest). For each eye size, eleven degrees of gaze stimuli were created by shifting each iris from its center by 0.25°, 0.13°, 0.063°, 0.031°, or 0.016° to the left, 0°, or 0.016°, 0.031°, 0.063°, 0.13°, or 0.25° to the right.
Experiment 2a employed a 4 (eye size) × 11 (degree of gaze) within-subjects factorial design. There were 528 trials, with conditions balanced across four blocks and randomized. Each trial (Fig. 2a) began with a fixation period of 1,000 to 1,400 ms (randomized in 100-ms increments), was followed by the presentation of a pair of eyes and fixation cross for 300 ms, and then a mask (rectangular mask subtended 11.1° × 2.8°) for up to 2,000 ms or until response. Participants responded during the presentation of the mask, indicating whether they perceived the eyes gazing left or right by pressing the “Z” key or the “/” key on the keyboard with their left or right index fingers, respectively, in a two-alternative forced-choice task. A red square appeared in the center of the screen for 200 ms as feedback if no response was given to keep participants on task (see Experiments 2a and 2b in the Supplemental Material for additional details).
Results
Prior to the main analysis, we examined the overall fidelity of gaze discrimination. Participants discriminated the smallest degree of gaze from center (0.016°) with an accuracy rate of 62.7%, significantly greater than chance level, t(25) = 8.6, p < .0001.
The two-alternative forced-choice data (“left” coded as 0; “right” coded as 1) from all participants were submitted to a logistic regression model of three predictors (degree of gaze, eye size, and Degree × Eye Size interaction; Collett, 1991) and a constant term. Degree of gaze was a highly statistically significant predictor of left/right responses (likelihood ratio = 6,780.0, p < .0001), which indicates that response accuracy improved as gaze shifted away from center in either direction. Eye size alone was not significant in predicting left/right responses (likelihood ratio = 0.41, p > .5). The Degree × Eye Size interaction was statistically significant (likelihood ratio = 23.8, p < .0001), which indicates that different eye sizes exerted different logistic regression curves. Specifically, the logistic regression curve slopes were steeper as eye size increased (Figs. 2c and 2d), which supported our hypothesis that participants should be better able to discriminate direction of gaze as eye size increased.
Experiment 2b: Physical Versus Emotional Eye Signals
Despite the fact that we used schematic eyes, it is likely that the larger eyes still expressed some degree of fear, which may have altered attention and discrimination accuracy. To examine this possible influence, we designed Experiment 2b to be identical to Experiment 2a except that we included a set of inverted eyes (using the same schematic eye stimuli; see Fig. 2b), which provided the same amount of physical gaze information as the noninverted set of eyes. We hypothesized that the change in eye configuration (e.g., the greater exposure of white sclera below the iris in inverted fear expression eyes) would reduce perceived fear for the larger eye sizes, as measured by subjective ratings, but would leave their enhanced gaze discrimination intact.
Method
Twenty-five undergraduates who had not taken part in the previous experiments provided informed consent and participated in return for $10. All had normal or corrected-to-normal vision. All aspects of the experiment were the same as in Experiment 2a except that we added four pairs of vertically inverted eyes, thereby doubling the number of trials (1,056). The experiment thus employed a 4 (eye size) × 11 (degree of gaze) × 2 (inversion) within-subjects factorial design.
Afterward, participants rated all eight pairs of eyes for how much they expressed each of six basic emotions: anger, disgust, fear, happiness, sadness, and surprise. Each pair of eyes (gazing straight ahead) was shown in random order, after which participants rated it for how much it conveyed each of the six emotions, using scales from 1 (not at all) to 9 (very strongly). Participants were informed that there were no right or wrong answers and that a pair of eyes could appear to express multiple emotions or no emotion.
Results
The ratings of fear were submitted to a 4 (eye size) × 2 (inversion) repeated measures ANOVA. There was a main effect of eye size, F(3, 66) = 9.0, p < .0001, η p 2 = .29, such that wider eyes were perceived as expressing more fear, F(1, 22) = 18.2, p < .001, η p 2 = .45. A main effect of inversion, F(1, 22) = 9.6, p < .01, η p 2 = .31, confirmed that inverted eyes were rated as expressing less fear than upright eyes. For example, inverted eyes of Size 3 were perceived to express as much fear as upright eyes of Size 1, the smallest size, t(22) = 0.04, p > .9 (see Fig. S1).
The gaze-discrimination data for all participants were split into upright and inverted sets and submitted to two separate logistic regression models, each identical to that used in Experiment 2a. Replicating the finding from Experiment 2a, there was a Degree × Eye Size interaction for upright eyes (likelihood ratio = 26.7, p < .0001). As hypothesized, gaze-discrimination enhancement was unaffected by perceived emotionality, as seen in a similar, and numerically greater, Degree × Eye Size effect for inverted eyes compared with upright eyes (likelihood ratio = 28.6, p < .0001). Analysis of the full data set as one logistic regression model with the inverted condition as an additional parameter did not significantly modify degree of gaze (neither interaction was significant—Degree × Inversion: likelihood ratio = 0.18, p > .6; Degree × Eye Size × Inversion: likelihood ratio = 0.04, p > .8).
Experiment 3: Peripheral Target Localization
We next examined whether eye widening in fear expressions would facilitate observers’ responsiveness in locating peripheral stimulus events (i.e., to “look here”), thus potentially extending the benefit of a clearer gaze-signal transmission. We used our schematic eyes in an attentional gaze-cuing paradigm in which fear expressions have been shown to direct more attention to cued locations than neutral gazes (Putman et al., 2006; Tipples, 2006). On one hand, if enhanced gaze discrimination reflects a special capacity for these eyes to interact with attention, we would expect larger eye sizes to interact with attentional-cuing effects. On the other hand, a lack of interaction would make a stronger case for a purely physical gaze enhancement. We predicted the latter on the basis of our thesis and prior evidence of an attention interaction dependent on communicated emotion (i.e., using full fear expressions, with the effect related to anxiety traits; Fox, Mathews, Calder, & Yiend, 2007; Putman et al., 2006).
In addition, using the same statistical-expression model, we maintained individual differences in eye shapes to create four eye sizes for each of the 19 exemplars (see Fig. S2) to generate broader variances in measurable eye features (Fig. 3a). We then related these features to response times to explore which might be the most influential functional features in eye widening.
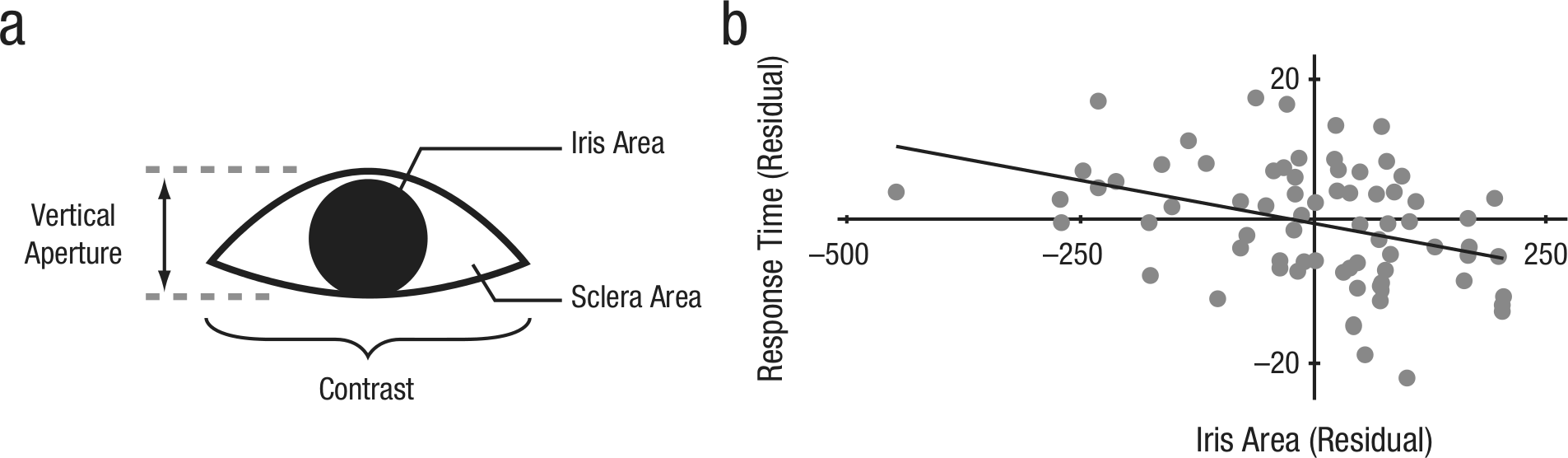
Revealed iris area predicts response time to correctly cued peripheral targets. The illustration (a) shows the four features extracted from each unique pair of schematic eyes. The graph (b) shows the relationship between the visible iris area revealed by eye widening and response times for correctly cued targets (r = −.378). Axes represent residuals, after controlling for baseline vertical-gaze perception using data collected in a separate experiment.
Method
Twenty-six undergraduates who had not taken part in the previous experiments provided informed consent and participated in return for $10 or course credit. All had normal or corrected-to-normal vision. The setup and apparatus were identical to those used in Experiment 2a.
Participants fixated on a 0.45° central cross and viewed one pair of the schematic eyes (one of the 19 exemplars in one of the four sizes) subtending 14.4° × 0.27° to 14.4° × 2.3°, which gazed straight ahead and then to either the left or the right by 1.0°. Participants then responded with their left or right index fingers to indicate the location of a target asterisk (0.55°), which appeared to the left of right of the eyes (8.6° left or right from fixation) in a position congruent or incongruent with gaze direction.
The experiment employed a 2 (cue congruence) × 4 (eye size) × 19 (exemplar) within-subjects factorial design. There were 304 trials, with eye-size and congruence conditions balanced across four blocks and randomized. Each trial began with a fixation period of 700 to 1,100 ms (randomized in 100-ms increments), and participants were instructed to fixate on the central location throughout the trial. The cross was replaced by a pair of eyes gazing straight ahead for 1,000 ms. The eyes then shifted gaze either left or right. After 200 ms, the target asterisk appeared for up to 800 ms or until response while the eyes continued to hold their gaze. A green (for a correct response) or red (for an incorrect response or no response) square appeared in the center of the screen for 200 ms as feedback to keep participants on task. (See Experiment 3 in the Supplemental Material for additional details.)
Results
Mean response times for only correct responses were submitted to a 2 (cue congruence) × 4 (eye size) repeated measures ANOVA. Consistent with an effect of attention, there was a main effect of cue congruence, F(1, 23) = 42.1, p < .0001, η p 2 = .65, such that participants responded faster to congruent cues than to incongruent cues, a result that replicated previous gaze-cuing findings (Friesen & Kingstone, 1998). The Congruence × Eye Size interaction was not statistically significant, F(3, 69) = 0.83, p > .4, η p 2 = .035, which indicated that eye widening did not modify this attentional-cuing effect. However, there was a main effect of eye size, F(3, 69) = 7.98, p = .0001, η p 2 = .26, such that increasingly wider eyes facilitated responses to peripheral targets, F(1, 23) = 12.7, p < .01, η p 2 = .36.
Our thesis was that enhanced gaze-direction decoding is brought about by physical parameters of the iris and sclera in widened eyes. To explore this relationship, we first extracted four features for all 76 pairs of eyes: vertical aperture, contrast, visible area of iris, and visible area of sclera (Fig. 3a). We then ignored eye-size groups and correlated each feature with response times for trials with congruent cues on which participants correctly responded; response times were averaged across all participants. After controlling for a vertical-directionality gaze effect (see Experiment 3 in the Supplemental Material), we found iris area to have the largest correlation to faster responses, r(72) = −.378, p = .001 (Fig. 3b), followed by contrast, r(72) = −.339, p = .003. Both of these factors survived statistical correction for multiple comparisons and were likely driven by the same variable, given their close correspondence, r(72) = .683, p < .0001.
Discussion
Eye widening not only increased the effective visual field of the expresser but also enhanced the discriminability of eye gazes for the observer and facilitated the speed with which observers used gaze cues to locate peripheral stimuli. These results were largely due to an augmented physical signal of the eyes, without the fearfulness signaled by the complete expression, and were retained under conditions of eye inversion, when fear perception was significantly diminished. An analysis of eye features suggested that the mechanism may be due to increased iris exposure and associated increased local iris-to-sclera contrast found in wider eyes. On the basis of these results, we argue that expressions of fear, because of their characteristic eye widening, provide a transfer of function from sensory to social, toward locating a common environmental event.
The visual benefit for fear expressers in Experiment 1 shows that wider fear eyes enhance stimulus discrimination and not just detection (Susskind et al., 2008), in that they preserved discrimination performance 9.4% farther out in the available periphery compared with narrower neutral eyes. This egocentric sensory function of fear is likely aligned with the increase in sympathetic autonomic tone (Levenson, 1992) associated with pupil dilation (Beatty & Lucero-Wagoner, 2000) and the conjunctive retracting of the eyelids through the involuntary, sympathetically innervated Müller’s muscle (Brunton, 1938).
The social importance of directional eye gaze is signified in the unique morphology of our eyes, which seemingly evolved to maximize its physical signal via the additional contrast of white sclera that likely facilitates observers’ ability to discriminate gaze direction (Kobayashi & Kohshima, 1997). The social utility of this information is indicated by the separate representations of left and right directional gazes in the superior temporal sulcus (Calder et al., 2007), which are proximal to a region important for the decoding of others’ mental states (Saxe & Powell, 2006). How aptly we process this information can be seen behaviorally, reflected in the accurate perception of minute changes in eye gaze (e.g., the smallest shift of gaze used in the present experiments, 0.016°, or half of one pixel in our experimental setup, possible through anti-aliasing, was reliably discriminated at a rate above chance level despite its brief, masked display) and in our remarkable sensitivity to different contexts of head orientation in correctly discriminating gaze direction (Jenkins, Beaver, & Calder, 2006; Langton, Watt, & Bruce, 2000).
Given that mere greater exposure of eye whites can activate the amygdala (Whalen et al., 2004) and that widened eyes are sufficient to signal recognizable fear (M. L. Smith, Cottrell, Gosselin, & Schyns, 2005), the recruitment of emotional circuitry, as well as some degree of emotion contagion (Harrison et al., 2006), to the wider eyes is possible. Thus, some emotional modulatory influence on gaze processing in the wider eyes cannot be completely ruled out. To obtain support for our physical- signal argument, we tested the same eyes inverted, because inverted fear expressions have been shown to reduce amygdalar activity (Sato, Kochiyama, & Yoshikawa, 2011) and attentional orienting toward fear (Bocanegra & Zeelenberg, 2009; Phelps, Ling, & Carrasco, 2006), although they also have been shown to retain some of their affective influences (e.g., Lipp, Price, & Tellegen, 2009). We showed that inverted eyes caused pronounced reductions in fear perception (Fig. S1) but exerted a pattern of gaze-discrimination enhancement equivalent to that of upright eyes (Fig. 2d), consistent with a physical rather than an emotional signal enhancement. Although it remains possible that inverted eyes evoked some emotional response in perceivers, it is unclear how distinct emotional responses may have enhanced (in the case of fear) or impaired (in the case of disgust) gaze discrimination. Rather, the most parsimonious account is the evident difference in physical signals originating from the iris and sclera.
Eye widening in emotional expressions may reflect some evolutionary or learned adaptation to enhance the physical signal of the eye. Fear and disgust expressions both communicate emotional states and are thus equal potential targets for attention, but the salience of the signal transmitted by their eyes is biased toward fear. This asymmetric salience of eye widening may enhance detection of these expressions from afar (F. W. Smith & Schyns, 2009), in addition to transmitting a clearer “look-here” signal, both of which functions are aligned with fear’s selective pressures toward rapid processing of distal, moving threats, as compared with the selective pressures of disgust, which tracks proximal, stationary threats (Anderson, Christoff, Panitz, De Rosa, & Gabrieli, 2003). In other words, fear and disgust are different kinds of threat responses, with the former appropriate for a potential predator or looming threat and the latter appropriate for a potential disease vector (e.g., Chapman et al., 2009).
Finally, we note some of the other adaptive effects of fear expressions. For the expresser, the pedomorphic features of widened eyes in fear expressions may solicit more socially sympathetic responses from neighbors (Marsh et al., 2005). For the observer, full fear expressions have been demonstrated to improve early vision (Phelps, Ling, & Carrasco, 2006), specifically along lower spatial-frequency channels (Bocanegra & Zeelenberg, 2009). This effect is further aligned with fear’s prioritized perception and action via projection of the magnocellular pathway to the dorsal stream (Vuilleumier, Armony, Driver, & Dolan, 2003; West, Anderson, Bedwell, & Pratt, 2010). Although these effects of full fear expressions are not incompatible with the physical signaling benefits shown here, predictions may diverge when it comes to other expressions that share the wide-eyed appearance of fear, such as surprise. Given that our findings reflect iris exposure and not fear perception, we would predict similar gaze-discrimination enhancements for surprise expressions but perhaps not the same attentional biases as fear. Although fear and surprise are placed into distinct categories and have distinct forms of attentionally salient features, such as mouth opening (Schyns, Petro, & Smith, 2009), it has been argued that similar forms of eye opening in fear and surprise reveal shared stimulus appraisals (Scherer, 2001). We hypothesized that this eye widening represents an underlying dimension of facial appearance related to sensory acquisition (Chapman et al., 2009; Susskind & Anderson, 2008; Susskind et al., 2008), reaching as far back in evolutionary ancestry as to be observed in dairy cattle (Sandem, Janczak, Salte, & Braastad, 2006). The additional prominent flashing of white sclera, unique to humans among primates (Kobayashi & Kohshima, 1997), suggests a physical augmentation toward optimizing the transmission of social signals (F. W. Smith & Schyns, 2009). And through this simple physical mechanism, a single expressive action links visual processing in expresser and observer.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.