
Abstract
In the present study, we examined the relationship between developmental modulation of socioaffective brain systems and adolescents’ preoccupation with social evaluation. Child, adolescent, and adult participants viewed cues indicating that a camera was alternately off, warming up, or projecting their image to a peer during the acquisition of behavioral-, autonomic-, and neural-response (functional MRI) data. Believing that a peer was actively watching them was sufficient to induce self-conscious emotion that rose in magnitude from childhood to adolescence and partially subsided into adulthood. Autonomic arousal was uniquely heightened in adolescents. These behavioral patterns were paralleled by emergent engagement of the medial prefrontal cortex (MPFC) and striatum-MPFC connectivity during adolescence, which are thought to promote motivated social behavior in adolescence. These findings demonstrate that adolescents’ self-consciousness is related to age-dependent sensitivity of brain systems critical to socioaffective processes. Further, unique interactions between the MPFC and striatum may provide a mechanism by which social-evaluation contexts influence adolescent behavior.
Keywords
Adolescence is a phase of the human life course defined by immense social change. Given that adolescents spend more time with peers than children and adults do (Brown, 2004), a unique feature of adolescent behavior is heightened attunement to, concern over, and reaction to perceived instances of peer evaluation. During adolescence, reported concern over social evaluation rises sharply from childhood (Westenberg, Drewes, Goedhart, Siebelink, & Treffers, 2004), reported daily self-consciousness peaks (Rankin, Lane, Gibbons, & Gerrard, 2004), and adolescents more frequently interpret themselves as being the target of social evaluation (leading to such phenomena as the imaginary audience; Elkind & Bowen, 1979).
An emerging viewpoint in neurodevelopmental research is that dynamic features of brain development influence unique aspects of behavior that emerge over the life course (Casey, Tottenham, Liston, & Durston, 2005; Somerville, Jones, & Casey, 2010). Despite the primary role of actual or perceived social evaluation in adolescents’ daily lives and well-being, little is known about the biological mechanisms that accompany phenomenological shifts in adolescent social concern. In the present study, we tested the hypothesis that experiential, autonomic, and socioaffective brain responses would change nonlinearly from preadolescence to postadolescence, even under the most basic of conditions—simply being looked at by a peer.
The current study focused on developmental modulation of the response properties and connectivity of the medial prefrontal cortex (MPFC). The MPFC is commonly engaged by social and emotional processes (Amodio & Frith, 2006; Roy, Shohamy, & Wager, 2012), and is a key node in neuroscientific models of the development of the adolescent self-concept (Sebastian, Burnett, & Blakemore, 2008). Given that the MPFC shows dynamic structural and connectivity-based maturation throughout the adolescent years (Shaw et al., 2008), we sought to evaluate the neurodevelopmental features of MPFC response and connectivity during instances of experimentally induced social evaluation. Sixty-nine human participants ranging in age from 8.0 to 22.9 years completed self-report, autonomic-arousal (galvanic skin response; GSR), and functional MRI (fMRI) measures to test (a) whether adolescents experience heightened emotional and autonomic responses to peer evaluation, (b) whether these responses extend to social-anticipation contexts, (c) whether such a behavioral profile is paralleled by distinct recruitment and connectivity patterns of the MPFC in adolescents, and (d) whether such potential effects subside or persist into early adulthood. In our analyses, we utilized age as a continuous variable to test for linear effects, quadratic effects (U or inverted-U shaped), and asymptotic effects (change during childhood and adolescence, stabilizing into adulthood) on responses to evaluation and anticipation periods.
Method
Participants
Sixty-nine healthy participants 8.0 to 22.9 years of age completed fMRI scanning. Participant volunteers were recruited from the New York City metropolitan area. See the Supplemental Material available online for demographics and inclusion and exclusion criteria. Adult participants provided informed written consent, and parental consent and subject assent were obtained for minors. The study was approved by the Weill Cornell Medical College Institutional Review Board.
Task
Participants were led to believe that a small, one-way video camera was embedded in the head coil of the fMRI scanner. They were told that this was a novel technology and that the experimenters wished to test it during their experimental session by cycling through its settings (off, warming up, on) several times. Participants were instructed to passively view the screen and, cued by low-level changes in the screen display, monitor the video camera’s status. The supposed video camera cycled among three phases (Fig. 1a). In the resting baseline condition, the display said “SYSTEM OFF.” In the anticipation condition, the display said “STARTING . . .,” and participants were told that the camera could turn on at any moment. In the evaluation condition, the display said “VIDEO ON,” and a red dot appeared.
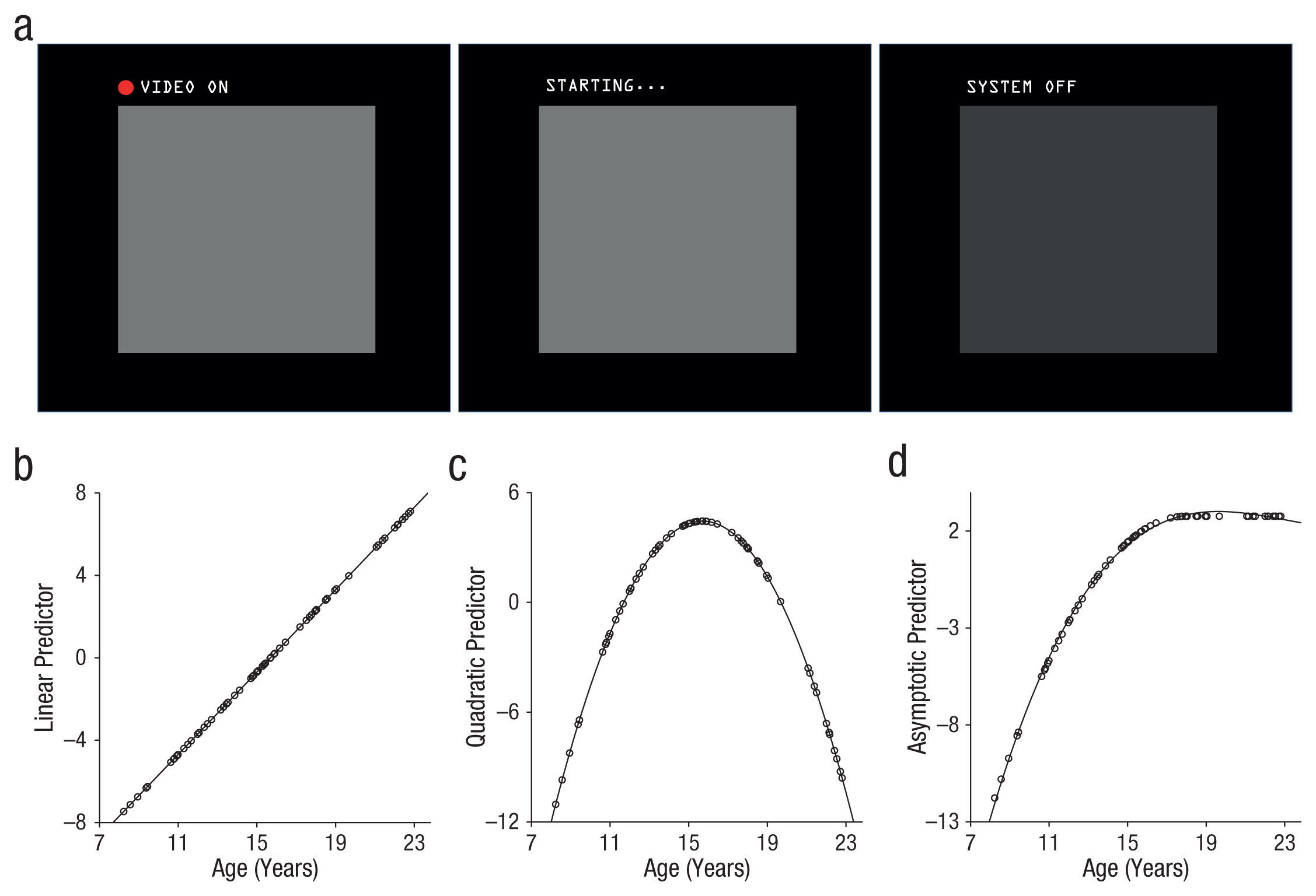
Experimental paradigm and predicted patterns of results. Participants were led to believe that a one-way video camera was embedded in the head coil of the functional MRI scanner. During different blocks of the experiment (a), participants were told that a camera was projecting an image of their face in real time (evaluation condition; left), warming up but not yet projecting (anticipation condition; middle), or off (baseline condition; right). The graphs show predicted linear (b), quadratic (c), and asymptotic (d) patterns as a function of participants’ age. Each dot represents a participant.
Participants were told that a same-sex peer of similar age would monitor the video feed during the participant’s scan and could see the participant’s face in real time whenever the camera was “on.” They were told the camera operated one way, and thus they should not expect to view a peer. Although there was, in fact, no camera, all participants had completed a separate peer-interaction task immediately prior to this study (Jones et al., 2011), which conveniently made the cover story more believable. Though it cannot be ruled out that the prior task influenced the present findings, the two tasks were held consistent across all participants, and each participant had a short break between the two studies.
The task was structured as a block design that pseudorandomly alternated among the rest, anticipation, and evaluation conditions. Participants saw a total of 12 blocks, 4 of each condition. To reduce predictability, we varied block length between 16 and 38 s. Across the task, participants spent an equal total duration in the anticipation and evaluation conditions (total time per condition: 92 s), and the mean duration of anticipation and evaluation blocks was matched (mean: 23 s). Participants viewed the resting baseline “camera off” stimulus for a total of 126 s in blocks averaging 31.5 s each.
Measures
Emotion
Immediately following each block, participants were asked to rate the extent to which they experienced the following emotions during that session: happiness, excitement, nervousness, worry, fear, and embarrassment. Participants rated each emotion category by bisecting a continuous line with the anchors “not at all” (far left) and “extremely” (far right). Ratings were completed for the anticipation and evaluation conditions separately and were not acquired for the rest blocks when participants believed the camera was off. See the Supplemental Material for further details.
Skin conductance
GSR was sampled simultaneously for 62 of the participants, with usable data acquired from 56 participants (see the Supplementary Material for further details). An MRI-compatible skin conductance recording system (GSR100C; Biopac Systems, Goleta, CA) together with AcqKnowledge 4.0 (Biopac) software continuously sampled skin conductance data at 100 Hz.
Neuroimaging
Participants were scanned with a General Electric (Milwaukee, WI) Signa 3.0 Tesla MRI scanner with a quadrature head coil. See the Supplemental Material for structural- and functional-acquisition sequences.
Data analysis
Age effects
Statistical analysis of each dependent variable (self-reported emotion, GSR, fMRI activity, fMRI connectivity) was used to assess the significance of three continuous age predictors, each assessing a distinct pattern of age-dependent change. The linear-age predictor with increasing age was modeled as a mean-centered linear age variable (Fig. 1b). The adolescent-specific predictor, used to detect U- or inverted-U effects for which adolescents differ from both children and adults, was modeled as a quadratic function (calculated by squaring the linear-age predictor; the quadratic peak fell at 15.94 years in the present sample; Fig. 1c). Finally, the adolescent-emergent predictor, which shows rapid change throughout adolescence and persists in magnitude into adulthood, was modeled with a mean-centered asymptotic predictor calculated by generating a quadratic function peaking at 18 years of age and retaining the maximum value for adult ages (Fig. 1d). The adolescent-emergent predictor closely mimicked a truncated cubic function for which the inflection point was fixed at 18 years of age.
Because the three predictors naturally share variance, group analyses consisting of a single “competing” statistical model incorporating all age predictors would be statistically invalid. Given that the objective of the present study was to assess age influences on self-conscious emotion and associated neural activity, every dependent variable was submitted to a triad of group statistical tests, each incorporating one continuous age predictor. From the triad of analyses, every age predictor that reached statistical significance is reported in the Results section and is represented by a fit line in the figures. Full analysis of variance (ANOVA) results for every age predictor are reported in the Supplemental Material. This approach mitigates model instability caused by multicollinearity and the need to engage in potentially biased experimenter choices regarding the importance of the three age predictors (e.g., choosing a testing order in stepwise group regressions or choosing to orthogonalize one predictor with respect to another). Though this approach does not permit direct quantitative comparison of the three age patterns, we believe it provides the most efficacious and unbiased method of identifying the age predictor (or predictors) that explains variance in the variables of interest.
Emotion
Self-reported emotion ratings were scored by recording the distance between the far left anchor and the point at which the participant bisected the line; greater values indicated greater endorsement of the emotion category. Raw scores were converted to proportions by dividing each score by the total line length, and then all measurements were divided by the total sum of scores for that participant.
Statistical analyses were conducted in IBM SPSS Statistics 19.0. A factor analysis on the self-report ratings indicated three latent variables corresponding to anxiety, positive arousal, and embarrassment (see the Supplemental Material). For each of the three emotion variables, a group ANOVA tested for effects of condition (anticipation, evaluation) and each of the three age predictors on self-reported emotion. Significant age effects were plotted to determine distribution, possible outliers, and directionality. Given the focus of the present article on developmental differences in task-evoked emotion, findings for the embarrassment ratings that yielded significant developmental differences are reported. Results for emotion ratings that did not show significant developmental differences are reported in the Supplemental Material. To account for independent tests of the three emotion variables, we interpreted each statistical test using an adjusted critical reliability coefficient (α = .0167; α = .05, Bonferroni corrected for three tests).
Skin conductance analysis
Skin conductance analysis in 56 usable participants was performed using AcqKnowledge 4.0 software and IBM SPSS Statistics 19.0. Skin conductance analyses focused on changes in response slope, or skin conductance level (SCL), per block. This standard analysis for block-design data (Dawson, Schell, & Filion, 2001) measures the signal-habituation rate during a task block, such that larger values correspond to less habituation, which is indicative of maintenance of autonomic arousal throughout the block. See the Supplemental Material for slope-calculation methods.
We used group analyses to test for effects of condition (anticipation, evaluation) and age on GSR, and we included baseline GSR as a covariate of no interest to account for task-independent variance in GSR reactivity across participants. Group analyses were conducted as described previously, with task time (first half of the experiment, second half of the experiment) additionally included as a within-subjects factor given the strong tendency for GSR effects to habituate over time (Andreassi, 2006; Dawson et al., 2001).
Neuroimaging
Functional-imaging data were preprocessed using Analysis of Functional NeuroImages (AFNI) software (Cox, 1996). See the Supplemental Material for details on preprocessing and first-level task-based general linear modeling (GLM). Following GLM estimation for each participant, we conducted random-effects group analysis, which consisted of a triad of linear mixed-effects group models with regressors dummy-coded to represent condition, participant, and each age predictor (Figs. 1b–d). Results yielded group statistical maps representing the main effect of condition, the main effect of each age predictor, and interactions of the condition-by-age predictor. Given the present focus on brain-behavior parallels, we focus in this article on age effects that persist for both anticipation and evaluation conditions as observed for embarrassment and GSR findings. However, a number of brain regions demonstrated significant condition-by-age interactions, which indicates differential age modulation for the anticipation and evaluation conditions. These regions and descriptions of age patterns are reported in Table S2 in the Supplemental Material.
Given dynamic changes in MPFC morphology and connectivity (Shaw et al., 2008), and motivated social behavior (Steinberg, 2004) from childhood to adulthood, we used group connectivity analyses to assess putative age modulation of coupling between the MPFC and systems of the brain critical to motivated behavior, such as the striatum (Robbins & Everitt, 1996). A whole-brain psychophysiological interaction (PPI) analysis (Friston et al., 1997) was carried out to identify selective MPFC task-based functional coupling that could be subsequently queried for age effects. (See the Supplemental Material for first-level PPI modeling methods.) Random-effects group analysis regressed voxel-wise PPI parameter estimates against each of the three age patterns of interest (linear, quadratic, and asymptotic). Resultant maps identified regions of the brain in which MPFC signal coupling during evaluation contexts fit each of the three age patterns.
All brain-imaging findings considered statistically significant exceeded correction for multiple comparisons to preserve a corrected reliability threshold (α ≤ .05) by using a p value-cluster size combination stipulated by Monte Carlo simulations run in the Clustsim subroutine in AFNI. The search space of the simulation constituted the spatial coverage obtained for functional images (42,341 voxels acquired; whole-brain coverage minus much of the occipital lobe). Thus, all imaging findings achieved a corrected significance level of p < .05 for the full acquisition space.
Significant age effects were plotted for inspection of distribution, possible outliers, and directionality by extracting parameter estimates for each participant from a 6-mm spherical region of interest around the cluster peak. These parameter estimates were also used in analyses to test possible sex differences and the relationship among dependent measures. They were also used to rule out potential age confounds in signal-to-noise ratio and motion (see the Supplemental Material).
Relationship among variables
Bivariate correlational analyses were conducted to quantify the degree of shared variance among self-reported embarrassment, GSR, and fMRI measures. Partial correlation analyses controlling for embarrassment and GSR were conducted to assess whether reported age effects in neural response remained significant when controlling for experiential measures. Results of this analysis (see the Supplemental Material) verified that the observed age differences could not be solely explained by covarying experiential differences across participants.
Results
The social-evaluation task elicited self-conscious emotion (e.g., increased ratings of embarrassment; Keltner & Haidt, 1999) and physiological arousal in adolescents. Repeated measures ANOVAs including condition (anticipation, evaluation) and each age predictor indicated that the adolescent-emergent age predictor yielded a significant main effect on embarrassment ratings, F(1, 67) = 6.07, p = .0163; η p 2 = .083; Bonferroni-adjusted critical α = .0167; (see the Supplemental Material and Fig. 2), with the adolescent-specific age predictor yielding trend-level prediction of embarrassment ratings, F(1, 67) = 5.52, p = .022; η p 2 = .076; Bonferroni-adjusted critical α = .0167, see the Supplemental Material. The estimated age of peak embarrassment ratings was 17.2 years.
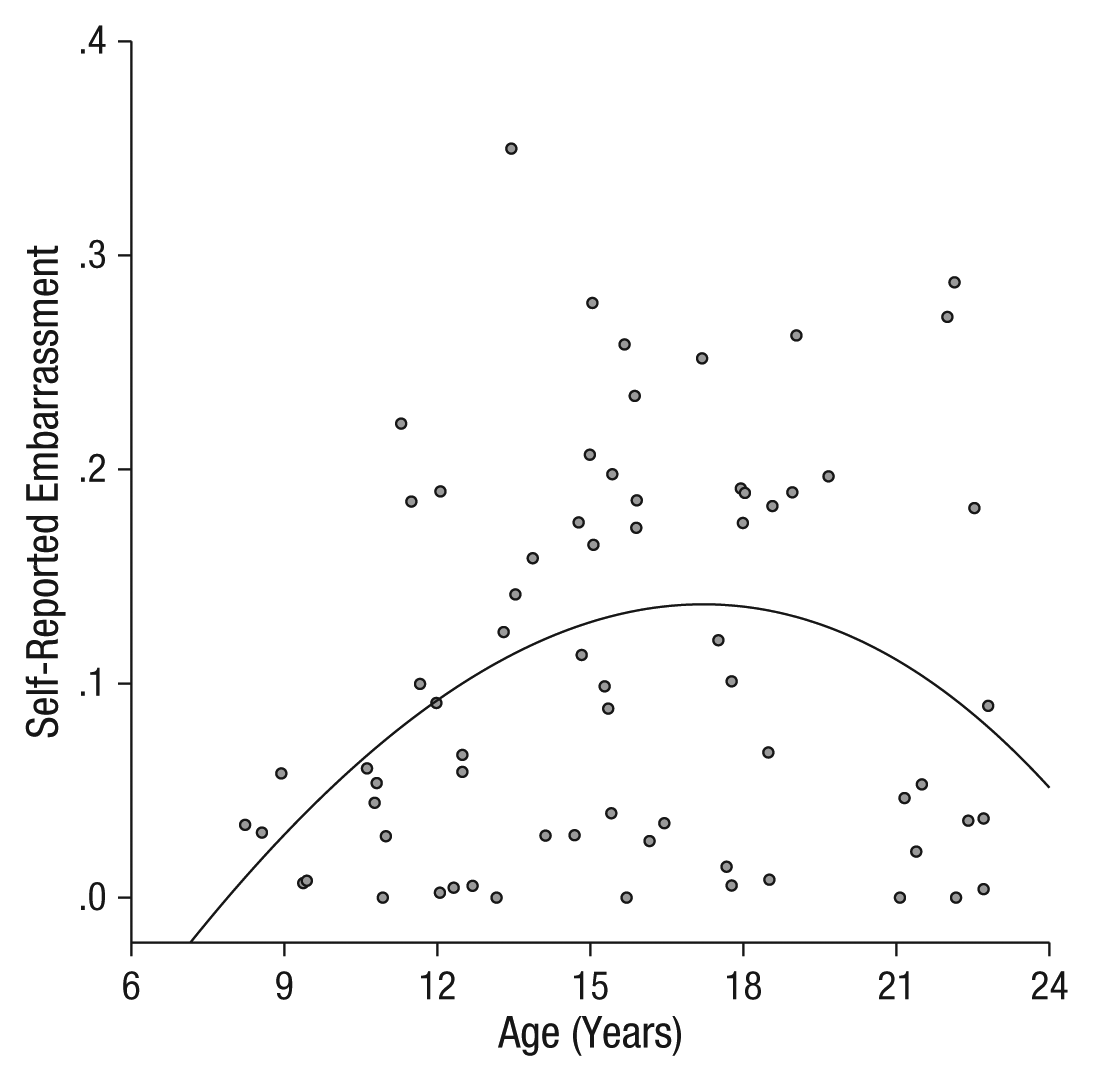
Scatter plot showing embarrassment ratings in the evaluation and anticipation conditions (collapsed) as a function of participants’ age. The fit line was derived from the adolescent-emergent predictor.
Given the significant adolescent-emergent effects, embarrassment ratings were further queried for modulation by condition. The adolescent-emergent age predictor yielded a trend-level effect for the anticipation condition, F(1, 67) = 3.57, p = .063; η p 2 = .051, and a significant effect for the evaluation condition, F(1, 67) = 7.14, p = .009; η p 2 = .096, which suggests consistency in age effects on embarrassment in both conditions but a more robust age difference during the evaluation phase. There was no main effect of condition on embarrassment ratings, F(1, 67) = 0.093, p = .76; Bonferroni-adjusted critical α = .0167.
Skin conductance data yielded a quadratic age-by-time interaction, F(1, 53) = 10.34, p = .002; η p 2 = .163. Post hoc analyses isolating the first half of the experiment indicated an adolescent-specific age effect, which can be described as greater autonomic arousal (less habituation of slope) in adolescence than in adulthood, F(1, 53) = 9.40, p = .003; η p 2 = .151 (see Fig. S1 in the Supplemental Material). The estimated age of peak GSR response was 14.38 years. There were no significant effects of condition or age during the second half of the experiment (ps > .2). Significant adolescent-specific age effects during the first half were evident for the anticipation and evaluation conditions when tested separately—anticipation: F(1, 53) = 4.77, p = .033, η p 2 = .083; evaluation: F(1, 53) = 6.95, p = .011, η p 2 = .12. These results suggest that minimal social-evaluative contexts are sufficient to induce heightened self-conscious emotion and physiological arousal that peaks in midadolescence.
A triad of voxel-wise, whole-brain, mixed-model ANOVAs were conducted with the repeated factor of condition (anticipation, evaluation) and each age predictor serving as a continuous covariate of interest. Guided by the behavioral findings, we focused fMRI analyses on revealing neural activations that were similarly engaged by the anticipation and evaluation conditions and differentially active as a function of the age predictors. A single region of the brain located in the MPFC (x = −13, y = 53, z = 6; 72 3 × 3 × 3 mm voxels; Brodmann’s area 32/10), mean cluster statistic: F(1, 67) = 11.84, p < .05 corrected (Fig. 3a), was significantly related to the adolescent-emergent age predictor. The estimated age of peak in MPFC activity was 15.25 years. The identical single region was also identified in an analysis of adolescent-specific age effects at a corrected threshold of p < .05, albeit smaller in size—mean cluster statistic: F(1, 67) = 10.38, 30 voxels. No regions demonstrated significant linear-age effects at whole-brain corrected thresholds. To summarize, mirroring the levels of experienced embarrassment and arousal, the MPFC demonstrated an elevated response in adolescents during both the anticipation and evaluation conditions that was partially matched in activity strength in young adults. Age differences in MPFC activity are plotted in Figure 3b for descriptive purposes.

Age differences in task-based functional brain activity. The statistical map identified functional activity showing heightened engagement of the medial prefrontal cortex (MPFC) through adolescence (relative to childhood) that persists into adulthood. The image threshold was p < .05, corrected for acquisition space. The scatter plot (b) shows MPFC responses in the evaluation and anticipation conditions (collapsed) as a function of participants’ age, for descriptive purposes. The solid fit line was derived from the adolescent-emergent predictor, and the dashed line was derived from the adolescent-specific predictor.
Given the powerful influence of social evaluation on motivated and affective behaviors in adolescence (Steinberg, 2008), we conducted PPI analyses to test the extent to which the MPFC demonstrates differentially selective connectivity during evaluation periods as a function of age (see the Supplemental Material for methodological details). Resultant statistical maps revealed MPFC coupling with the dorsal striatum that significantly fits an asymptotic age pattern—left caudate: x = −8, y = 20, z = 6; 35 3 × 3 × 3 voxels; mean cluster statistic: F(1, 67) = 10.24; p < .05, corrected (Fig. 4). No other activations were observed for this analysis. No regions demonstrated significant linear-age or adolescent-specific (quadratic) effects at whole-brain corrected thresholds. Thus, the transition from childhood through adolescence predicts the emergence of MPFC-striatum coupling during evaluation contexts, a pattern that persists into adulthood.

Age differences in task-dependent functional connectivity. Whole-brain connectivity analysis identified selective functional coupling between the medial prefrontal cortex (MPFC) and striatum during evaluation, which was greater in adolescence (relative to childhood) and persists into adulthood. The image threshold was p < .05, corrected for acquisition space.
Discussion
Using a simulated social-evaluation task, we observed that being watched by a peer was sufficient to generate nonlinear differences among children, adolescents, and young adults in self-conscious emotion and related physiological indices of emotional arousal. The nonlinear pattern of embarrassment and skin conductance findings supports the hypothesis that even subtle social-evaluative contexts—and anticipation of them—lead to heightened self-conscious emotion and arousal during adolescence. Self-conscious emotion rose in adolescents (relative to children) and stabilized in adults, whereas arousal (skin conductance) levels showed maximal levels in adolescents.
Numerous factors are thought to converge during adolescence and contribute to the central role of social evaluation in adolescents’ everyday experience. The transition to adolescence is typically marked by a rise in the frequency and intensity of peer interaction; this increase has been documented in both humans and animals (Cairns, Leung, Buchanan, & Cairns, 1995; Primus & Kellogg, 1989) and is often manifested among humans by high rates of digital communication (Wang, Iannotti, & Nansel, 2009). Such a shift in investment likely reflects a heightened motivation for peer acceptance, rendering social-evaluation contexts increasingly salient to adolescents.
In the present study, we evaluated whether adolescents instantiate unique neural response patterns to instances of social evaluation by peers that parallel phenomenological shifts in social sensitivity. We observed that behavioral shifts in adolescent social sensitivity were accompanied by nonlinear changes in MPFC response magnitude and selective MPFC-striatum connectivity, which were significantly higher in adolescents (relative to children) and slightly lower in early adulthood (relative to adolescence).
Though the present study was optimized to detect such age differences across the whole brain (except for the occipital lobe), nonlinear age effects were highly circumscribed to the MPFC. The specificity of effects to the MPFC converges with widely supported theories of the MPFC’s key role in social cognition and emotional valuation processes (Amodio & Frith, 2006; Blakemore, 2008). These findings extend existing accounts of MPFC function to suggest that it maintains persistent representation in even the most basic of social-evaluative contexts—being looked at.
Although the MPFC is frequently conceptualized as specialized for social cognition, emerging theoretical viewpoints have noted common recruitment of the MPFC during contexts that draw on affective valuation and assessment of significance to the self (Krienen, Tu, & Buckner, 2010; Roy et al., 2012)—which are often, but not exclusively, “social.” Indeed, the nonlinear age differences we observed were also evident in anticipatory situations during which participants believed they were not being viewed but would be imminently, which indicates that explicit evaluation was not necessary to invoke adolescent self-consciousness and its neural correlates. Following this view, we propose that MPFC activity in the present study served to incorporate salient contextual cues (in this case, imminent or perceived social evaluation) with emotional valuation processes. Thus, emergent heightened magnitude of MPFC activity in adolescence could result in assignment of heightened emotional value and self-relevance to instances of supposed social evaluation. This conceptualization is consistent with prior findings indicating that MPFC response to positive and negative social feedback is exaggerated in individuals for whom social feedback is particularly salient: individuals with low self-esteem (Somerville, Kelley, & Heatherton, 2010).
Robust MPFC signaling paired with increasing connectivity between MPFC and striatal regions could provide a mechanism by which peer-evaluation contexts come to increasingly modulate adolescent motivated behavior. Not only does social concern serve as a motivating force that drives adolescents to seek out social bonds (Steinberg & Morris, 2001), but adolescents are also more prone to engage in suboptimal choice behaviors (e.g., risky driving) when with peers than when alone (Gardner & Steinberg, 2005). The striatum plays a key role in incorporating motivational, control, and contextual signals to facilitate context-dependent learning and behavior (Alexander, DeLong, & Strick, 1986). Though tentative, the observed pattern of MPFC-striatum connectivity might selectively upregulate motivational signaling, effectively compelling adolescent behavior toward action or approach when being evaluated by peers. Although consistent with extant models of peer influence on adolescent decision making (Lenhart, Ling, Campbell, & Purcell, 2010; Somerville & Casey, 2010; Steinberg, 2008), findings of the current study illuminate the key role played by the MPFC in maintaining a representation of peer evaluation and its emotional qualities, whereas selective connectivity with the striatum may provide a means for integrating signals relevant to social context with motivational systems that govern goal-directed behavior.
It should be noted that the age effects observed for behavioral measures (embarrassment and GSR) partially correspond with the age-related changes observed for the brain-imaging measures. All measures showed a robust influx of response from childhood into adolescence, but measures diverged into young adulthood: GSR levels showed lower magnitude in young adults than in adolescents, whereas the other measures demonstrated a partial or no decline in adults relative to adolescents. Future research with a broader age range is needed to determine whether the divergence of age patterns in young adults is reliable.
A second feature of the reported findings is that physiological, MPFC, and self-conscious emotion demonstrate common maximal responding during adolescence, as indicated by analyses solving for the peak age of response using fit-line equations. Future research may assess whether the particular convergence of measures during adolescence plays a functionally significant role in promoting social sensitivity. There are also subtle and intriguing differences among the peak ages for GSR (14.38 years), MPFC activity (15.25 years), and self-reported embarrassment (17.2 years). Each measure contains its own profile of measurement error, so comparing the particular timing differences among variables should be interpreted cautiously. However, these findings provoke speculation that social sensitivity resonates in physiological and neural indices at an earlier age than when these emotions are most strongly labeled as self-conscious per se. Though even young children are capable of understanding embarrassment (Seidner, Stipek, & Feshbach, 1988), the current findings suggest that the process of attributing such physiological patterns as “embarrassment” might not manifest until later in adolescence, perhaps because of perspective-taking skills that continue to improve throughout adolescence (Crone & Dahl, 2012; Dumontheil, Apperly, & Blakemore, 2010) that may scaffold simulation of negative consequences of potential social transgressions that serve as a foundation of embarrassment (Keltner & Haidt, 1999). A complementary explanation is that though social evaluations are arousing across different stages of adolescence, they might be experienced as less specifically embarrassing in early and midadolescence relative to late adolescence. Comprehensive studies with broader emotion measures will be needed to address these possibilities.
In conclusion, waiting to be looked at and believing one is being looked at were sufficient to induce nonlinear changes in self-conscious emotion and related physiological indices among children, adolescents, and young adults. Nonlinear differences in response in the MPFC, and MPFC-striatum connectivity, parallel this behavioral shift, and we propose that these response differences influence adolescent social sensitivity. The functional properties of the MPFC are likely to be influenced by continued structural maturation (Mills, Lalonde, Clasen, Giedd, & Blakemore, 2012; Shaw et al., 2008) and subcortical and cortical connections (e.g., Asato, Terwilliger, Woo, & Luna, 2010) during adolescence. That said, future work will be needed to identify biological and experiential mechanisms that give rise to the functional differences illuminated by the present study. Together with other findings, the results of this study bridge examinations of psychosocial development and neurodevelopmental science to show how the emergent features of the adolescent social life can exert such a powerful influence over motivation, emotion, and well-being.
Footnotes
Acknowledgements
We thank Frederico Laurenço, Juan Molina, Joe Moran, Alea Skwara, and Lucas Wandrey for their contributions to data collection and statistical analyses, and the staff at the Biomedical Imaging Core Facility of the Citigroup Biomedical Imaging Center at Weill Cornell Medical College.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by K99 MH078713 (to L. H. S.) and the Mortimer D. Sackler family.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.