
Abstract
Mature amygdala-prefrontal circuitry regulates affect in adulthood but shows protracted development. In altricial and semialtricial species, caregivers provide potent affect regulation when mature neurocircuitry is absent. The present investigation examined a potential mechanism through which caregivers provide regulatory influences in childhood. Children, but not adolescents, showed evidence of maternal buffering, such that maternal stimuli suppressed amygdala reactivity. In the absence of maternal stimuli, children exhibited immature amygdala-prefrontal connectivity. However, in the presence of maternal stimuli, children’s connectivity was more mature, resembling adolescents’ connectivity. Children showed improved affect-related regulation in the presence of their mothers. Individual differences emerged, with greater maternal influence on amygdala-prefrontal circuitry associated with stronger mother-child relationships and maternal modulation of behavioral regulation. These findings suggest a neural mechanism through which caregivers modulate children’s regulatory behavior by inducing more mature connectivity and buffering against heightened reactivity. Maternal buffering in childhood, but not adolescence, suggests that childhood may be a sensitive period for amygdala-prefrontal development.
Keywords
Adults rely on intrinsic sources of regulation to manage their emotions and effectively navigate social interactions and the environment (Gross, 1998). At the neural-systems level, emotion regulation is supported by mature amygdala-prefrontal circuitry in healthy adults (Banks, Eddy, Angstadt, Nathan, & Phan, 2007; Hariri, Mattay, Tessitore, Fera, & Weinberger, 2003; Kim, Somerville, Johnstone, Alexander, & Whalen, 2003). However, children have yet to develop these internal mechanisms of regulation, as amygdala-prefrontal circuitry undergoes protracted development through adolescence (Gee, Humphreys, et al., 2013; Guyer et al., 2008; Hare et al., 2008; Monk et al., 2003). Because of this relative immaturity, external sources of regulation (primarily caregivers) play a critical role in children’s emotion regulation (Hofer, 1994; McCoy & Masters, 1985).
Early in life, the caregiver, often the mother, is one of the most potent stimuli for altricial and semialtricial species and plays an essential role in typical behavioral and neural development (Tottenham, 2012). The mother 1 provides widespread regulatory influences in childhood that include physiological, thermal, and nutritional regulation (Hofer, 1994). The affective mother-child bond is intense and learned, resulting in preference for the mother and wariness of strangers in infants, and greater approach-related behavior (Zarbatany & Lamb, 1985), greater exploration (Ainsworth & Bell, 1970), and less fear (Campos, Emde, Gaensbauer, & Henderson, 1975) in children when the mother is present than when the mother is absent. These studies suggest that the mother is uniquely relevant as an external source of regulation when the child approaches and engages with the environment.
In nonhuman animals and humans, the maternal stimulus is distinguished from nonmaternal stimuli at the neural level as well (Caldji et al., 1998; Hostinar, Sullivan, & Gunnar, 2014; Moriceau & Sullivan, 2006; Plotsky et al., 2005; Todd, Evans, Morris, Lewis, & Taylor, 2011; Tottenham, Shapiro, Telzer, & Humphreys, 2012). In rodents, maternal presence reduces amygdala activity in the pup, thus promoting attachment behaviors and reducing fear (Moriceau & Sullivan, 2006). Given the functional homologies between rats and humans early in life, it is possible that the mother exerts a similar influence on amygdala circuitry in humans, although the latter remain dependent on maternal care for a much longer period than do the former. Thus, the present study focused on neural mechanisms underlying the mother’s external regulation of emotion in human development.
Recently, we characterized developmental changes in the amygdala-prefrontal circuit that supports emotion regulation in adulthood, with evidence suggesting a shift from an immature to a mature phenotype of amygdala-prefrontal circuitry around the transition between childhood and adolescence. Specifically, children exhibit positive connectivity between the amygdala and medial prefrontal cortex (mPFC) and higher amygdala reactivity, whereas adolescents and adults exhibit negative amygdala-mPFC connectivity and lower amygdala reactivity (Gee, Humphreys, et al., 2013). For children, the normative state is having access to their caregiver, who can provide external regulation of developmentally normative heightened amygdala reactivity. We hypothesized that maternal stimuli might induce phasic modulation of amygdala-mPFC circuitry to promote emotion regulation in childhood by modulating amygdala-mPFC connectivity (instantiating connectivity akin to that in mature individuals) and reducing amygdala and affective reactivity. We anticipated that this effect would be absent in adolescence, a time of increasing independence from the mother. In order to examine the potential mechanism through which the mother provides affect regulation, the present study used a functional MRI (fMRI) task involving maternal and nonmaternal stimuli. In addition, an affect-related regulation task was used to examine whether the presence of the mother effectively modulates amygdala-prefrontal connectivity and associated regulatory behavior.
Method
Participants
Participants in the fMRI session (N = 53) were 23 children (ages 4–10 years old, mean age = 7.34, SD = 2.14; 14 females, 9 males) and 30 adolescents (ages 11–17 years old, mean age = 14.21, SD = 1.87; 18 females, 12 males). A maximum sample size of 60 participants was determined a priori to account for expected data loss due to excessive motion during fMRI scanning. We aimed to obtain at least 50 participants with usable scanning data to ensure sufficient sampling across the age range of 4 to 17 years. Data collection was stopped following completion of the targeted enrollment. Seven participants were excluded because of excessive motion in their fMRI data. All participants were physically and psychiatrically healthy (no medical or psychiatric disorders), as confirmed by a telephone screening prior to participation. The Child Behavior Checklist (CBCL; Achenbach, 1991) was used to assess clinical symptoms, and all participants fell within the normal range on the CBCL Total Problems, Internalizing Problems, and Externalizing Problems subscales.
Of the original 53 participants, 47 also completed a go/no-go task, which measured regulatory behavior in an affective context. These 47 participants had a mean age of 11.44 years (SD = 3.86, range = 4–17). Of the 6 participants who did not complete the behavioral session, 3 were children and 3 were adolescents.
Participants were from European American (37.7%; 34.8% of children, 40% of adolescents), African American (32.1%; 26.1% of children, 36.7% of adolescents), Asian American (15.1%; 26.1% of children, 6.7% of adolescents), American Indian (5.7%; 4.3% of children, 6.7% of adolescents), and other (9.4%; 8.7% of children, 10% of adolescents) backgrounds. Among participants, 22.6% identified as Latino (21.7% of children, 23.3% of adolescents). Cognitive ability was assessed using the Wechsler Abbreviated Scale of Intelligence (Wechsler, 1999) for participants between the ages of 6 to 17 years (assessments were conducted with 48 participants). The average full-scale IQ of the sample was 110 (SD = 16). Data on household income were obtained for the families of 49 participants; modal income was between $55,001 and $70,000, and there was no difference between the child and adolescent groups. Data on primary caregiver’s education was obtained for families of 52 participants: There was a modal education level of a 4-year college degree for both the child and adolescent groups. All participants were right-handed. The protocol was approved by the University of California, Los Angeles Institutional Review Board. Participants provided informed consent or assent (parental informed consent for minors).
Self- and parent-report measures
Anxiety was measured using the parent report form of the Revised Child Anxiety and Depression Scale (Chorpita, Moffitt, & Gray, 2005). The current study was primarily interested in the subscale for separation anxiety. Attachment-related behaviors were measured using the Security Scale (Kerns, Klepac, & Cole, 1996). Attachment security is difficult to measure after infancy (during which, measurement relies on the Ainsworth Strange Situation task); however, the Security Scale assesses children’s perceptions of security in parent-child relationships in middle childhood and early adolescence. The instrument provides scores for three subscales: (a) children’s belief that their attachment figure is responsive and available, (b) children’s reliance on the attachment figure in times of stress, and (c) children’s ease and interest in communicating with the attachment figure.
Procedure
Participants came to the laboratory for two sessions. In the first session, behavioral measures were collected and children performed an affect-related regulation task in the presence of their mother and separately in the presence of a stranger (an experimenter). Children were also acclimated to the scanner environment with an MRI replica. During the second visit, which occurred on a separate day, the mother/stranger task was administered in the MRI scanner. Participants completed the fMRI session an average of 3.81 months (SD = 4.34, range = 0–20) following the behavioral session.
Mother/stranger fMRI task
Participants completed a block-design mother/stranger task while in the MRI scanner. Based on rodent studies that present olfactory cues associated with the mother to the developing rodent (reviewed in Moriceau, Roth, & Sullivan, 2010), the present task used visual cues (the mother’s face) representative of the mother. Participants viewed pictures of their mother and an ethnicity-matched unfamiliar individual, who was the mother of another child (a stranger), in alternating blocks of 28 s each. Both mothers and strangers posed with happy and neutral expressions, and all models wore white material around their necks; thus, for each participant, there were two mother stimuli and two stranger stimuli. These images were obtained in the laboratory and standardized for size and luminance. The face images were in full color with a vertical visual angle of approximately 15°. Participants were instructed to respond quickly to the happy facial expression (regardless of model), which was presented 50% of the time in the same random order to all participants.
There were four blocks of mother stimuli (M), four blocks of stranger stimuli (S), and three blocks of fixation (+), which were presented with mother and stranger blocks alternating (+MSMS+SMSM+). The starting block (mother or stranger) was counterbalanced across subjects. Each block contained 18 images of either the participant’s mother (happy and neutral expressions) or a stranger (happy and neutral expressions), which yielded a total of 144 stimulus presentations—72 mother trials and 72 stranger trials. Each face stimulus was presented for 500 ms followed by 1,000 ms of fixation. Thus, participants were allowed approximately 1,500 ms to respond, which they did by pressing a button with their index finger. Participants viewed images through VisuaStim Digital video goggles (Version 8; Resonance Technology). A 932 fORP response box (Current Designs, Philadelphia, PA) was used for recording behavioral responses. The entire task lasted 4:54 min. Prior to scanning, participants were given the opportunity to practice to ensure that they understood and could perform the task.
Scanning parameters
Participants were scanned with a Siemens Trio 3-T fMRI scanner (Siemens, Erlangen, Germany). A whole-brain, high resolution, T1-weighted anatomical scan (magnetization-prepared rapid-acquisition gradient echo; 192 × 192 in-plane resolution, 250-mm field of view; 176 mm × 1 mm sagittal slices) was acquired for each subject for transformation and localization of functional data into Talairach space (Talairach & Tournoux, 1988). For the functional run of the mother/stranger task, we collected 143 T2*-weighted echo-planar images—34 slices, slice thickness = 4 mm (skip 0), repetition time = 2,000 ms, echo time = 30 ms, flip angle = 90°, matrix size = 64 × 64—at an oblique angle of approximately 30°.
fMRI preprocessing
Functional imaging data were preprocessed and analyzed with the Analysis of Functional NeuroImages (AFNI) software package (Medical College of Wisconsin, Milwaukee, WI). All included data were free of movement greater than 2.5 mm in any direction. After slice-time correction, images were registered to the first image volume after the high-resolution anatomical data set with rigid body transformations and smoothed with an anisotropic 6-mm Gaussian kernel. Time series were normalized to percentage signal change to allow comparisons across runs and individuals by dividing signal intensity at each time point by the mean intensity for that voxel and multiplying the result by 100. The model included regressors for each of the two stimulus types (mother and stranger) by convolving the stimulus-timing files using a canonical hemodynamic response function. Six motion parameters were included as separate regressors, for a total of eight regressors. General linear modeling (GLM; random effects) was performed to fit the percentage signal change time courses to each regressor. Linear and quadratic trends were modeled in each voxel time course to control for correlated drift. Group-level analyses were conducted on the regression coefficients from the individual analysis after transformation into the standard coordinate space of Talairach and Tournoux (1988) with parameters obtained from the transformation of each subject’s high-resolution anatomical scan. Talairach-transformed images had a resampled resolution of 3 mm3.
Task-dependent functional connectivity
A whole-brain psychophysiological-interaction (PPI) analysis was conducted to examine whether the mother-versus-stranger context modulated functional connectivity with the amygdala. The PPI analysis tested the extent to which the amygdala covaried with other brain regions more during the viewing of mother faces than during the viewing of stranger faces. A GLM analysis was performed in AFNI for each participant, with regressors for task, seed-region time series, interaction of task and time series, accuracy, and the six motion regressors. Two psychological (task) regressors modeled whether a given trial consisted of viewing the mother’s face or the stranger’s face. The physiological (seed-region time series) regressor comprised the time series for the right anatomically defined amygdala. Two interaction regressors modeled the interaction of the psychological regressors and the physiological regressor, such that each interaction regressor identified regions whose time series correlated in a task-dependent manner with the amygdala time series. The GLM analyses fit the percentage-signal-change time series to each regressor, and linear and quadratic trends were modeled for each voxel’s time series to control for correlated drift. The individual-level regression coefficients were then submitted to random-effects, group-level analyses.
Statistical and behavioral data analysis
Participants were grouped by age (4–10 years vs. 11–17 years) on the basis of prior studies demonstrating a valence switch in amygdala-mPFC coupling around age 10 (Gabard-Durnam et al., 2014; Gee, Gabard-Durnam, et al., 2013). We performed a 2 × 2 repeated measures analysis of variance (ANOVA) to test for effects of age group (children vs. adolescents) and condition (mother vs. stranger) on beta weights for amygdala reactivity and functional connectivity. Individual differences in anxiety and attachment were examined using univariate ANOVAs with factors for age group and valence of amygdala-mPFC connectivity (positive or negative to the mother, relative to implicit baseline).
The right anatomical amygdala, as defined by the Talairach & Tournoux Atlas in AFNI, was selected as a region of interest (ROI). We were particularly interested in the right amygdala because literature on hemispheric asymmetry suggests that the left hemisphere is more involved in positive emotion, whereas the right hemisphere is more threat sensitive and involved in negative emotion (Canli, Desmond, Zhao, Glover, & Gabrieli, 1998; Compton, Heller, Banich, Palmieri, & Miller, 2000), even in infants (Davidson & Fox, 1982). Thus, we hypothesized that maternal buffering would occur via the right amygdala, which would normally mediate the monitoring of threat presence or absence. In addition, we have previously shown that the presence of the mother potentiates activation in the left amygdala (Tottenham et al., 2012). Results for the left amygdala (also defined anatomically using the Talairach & Tournoux Atlas in AFNI) are presented in the Supplemental Material available online. The mPFC ROI consisted of z = 10–20 of the anatomically defined anterior cingulate cortex in the Talairach & Tournoux Atlas in AFNI. This particular region was selected based on prior work demonstrating its role in affective tasks (Bush, Luu, & Posner, 2000), as well as changes in its connectivity with the amygdala across typical development (Gabard-Durnam et al., 2014; Gee, Humphreys, et al., 2013); beta weights were extracted from the PPI analysis for this ROI.
For each participant, we calculated accuracy and mean reaction time (RT) for each condition (mother, stranger). We calculated accuracy as the difference between the total number of trials (72) and total errors. Total errors equaled the sum of false alarms to neutral faces (errors of commission; i.e., responding to a neutral face) and misses to happy faces (errors of omission; i.e., not responding to a happy face). We calculated the mean RT for correct responses (hits) to happy faces and for false alarms to neutral faces.
Motion correction
Systematic procedures were implemented to reduce motion, particularly in younger participants, and to ensure that children remained still throughout the duration of the task. Before the scanning session, children participated in a mock scanning session to help them acclimate to the scanning environment and feel comfortable with the scanning procedures. In addition, this step provided an opportunity for children to practice and receive feedback on lying still in order to optimize children’s ability to remain still during actual data collection. During data collection, an air-vacuum pillow (Siemens Comfort Pack) was used to pad and secure the child’s head in a comfortable, steady position. Additional padding was placed around the child’s head. In addition, all participants were provided with feedback and reminders regarding motion throughout the scanning session.
Volumes with motion greater than 2.5 mm in any direction were excluded (via censoring), and all participants had fewer than 27% of total volumes censored (mean censored volumes = 1.4%; mode = 0%). Preprocessing included standard spatial realignment to correct for motion. Motion regressors were included in our imaging analyses (at the subject level, motion in all six directions at the trial-by-trial level). In addition, multiple analyses were conducted to rule out potential effects of motion. Average motion (across all six directions) did not correlate with age, amygdala reactivity, or amygdala-mPFC functional connectivity and did not differ between the two age groups (allps > .05).
Given recent advances in methods for controlling for motion, we also conducted a secondary analysis in which we reanalyzed our functional-connectivity data controlling for different motion levels across participants (Van Dijk, Sabuncu, & Buckner, 2012). The mean absolute-displacement value was calculated for each participant and was entered as a covariate into the repeated measures ANOVA examining functional connectivity in response to mothers versus functional connectivity in response to strangers as a function of age group. Results of this secondary analysis replicated our original finding of the Condition × Age Group interaction, F(1, 49) = 3.98, p = .052. Moreover, mean displacement value was not associated with age, amygdala reactivity, or amygdala-mPFC functional connectivity and did not differ between the two age groups (ps > .05).
Affect-related regulation task
At least 1 day prior to completing the mother/stranger task, 47 participants completed an affect-related regulation task (Hare et al., 2008; Tottenham, Hare, & Casey, 2011) outside of the scanner. In this go/no-go task, participants viewed facial expressions of emotion, pressing a button when a target facial expression (go) appeared and inhibiting this behavioral response when a distractor facial expression (no-go) appeared. Stimuli consisted of color images of four female faces from the Karolinska Directed Emotional Faces database, each of which was shown with happy, sad, and neutral expressions (Lundqvist, Flykt, & Öhman, 1998). The face images were in full color with a vertical visual angle of approximately 12°. The three task blocks consisted of the following: happy face as the go expression and sad face as the no-go expression, happy face as the go expression and neutral face as the no-go expression, and neutral face as the go expression and happy face as the no-go expression. Participants were instructed to press a response button quickly for the go face and were not told which expression would be the no-go stimulus. Blocks were counterbalanced across participants. The go facial expression was presented 66.6% of the time to create a prepotent tendency for responding. Stimuli were presented in the same random order to all participants. Each face stimulus was presented for 500 ms followed by 1,000 ms of fixation to ensure enough time for responding.
The task was administered twice, once in the presence of the participant’s mother and once in the presence of a stranger (a research assistant). During each administration, the mother or stranger sat next to the participant while he or she completed the task. Participants were instructed to focus on the task in both conditions, and mothers were instructed not to speak with the participant during the task but instead to occupy themselves with paperwork. The order of administration (mother or stranger first) was counterbalanced.
For each participant, we calculated accuracy as the difference between the number of hits in response to go faces and the number of false alarms (i.e., responding to a no-go face). We calculated the mean RT for hits in response to neutral faces in the context of happy and sad no-go faces. Repeated measures ANOVAs assessed differences in behavioral performance when participants were in the presence of the mother versus the stranger. A repeated measures ANOVA tested the effects of amygdala-mPFC connectivity valence (between-subjects factor) and mother or stranger presence (within-subjects factor) on behavior. To assess whether amygdala reactivity related to how children performed in the presence of their mother or a stranger, we tested whether children with greater suppression of the amygdala in response to mothers than to strangers differed from children with less amygdala suppression to mothers on false alarm rate when in the presence of their mother or a stranger.
Results
fMRI session: does the maternal stimulus alter amygdala-mPFC circuitry?
Effect of the mother on amygdala reactivity
Beta weights corresponding to blood-oxygen-level-dependent (BOLD) signals were extracted from the right amygdala. A Condition × Age Group interaction demonstrated that differential activation to mother versus stranger stimuli depended on age, F(1, 53) = 4.065, p = .049, η2 = .071 (medium effect size; Cohen, 1988). Specifically, right amygdala reactivity in children, which is typically high (Gee, Humphreys, et al., 2013), was suppressed when participants viewed their mother, relative to when they viewed a stranger (Fig. 1). By contrast, adolescents did not show differential amygdala reactivity to mothers versus strangers. Figure S1 in the Supplemental Material shows amygdala reactivity to mothers and strangers versus the implicit baseline for children and adolescents.
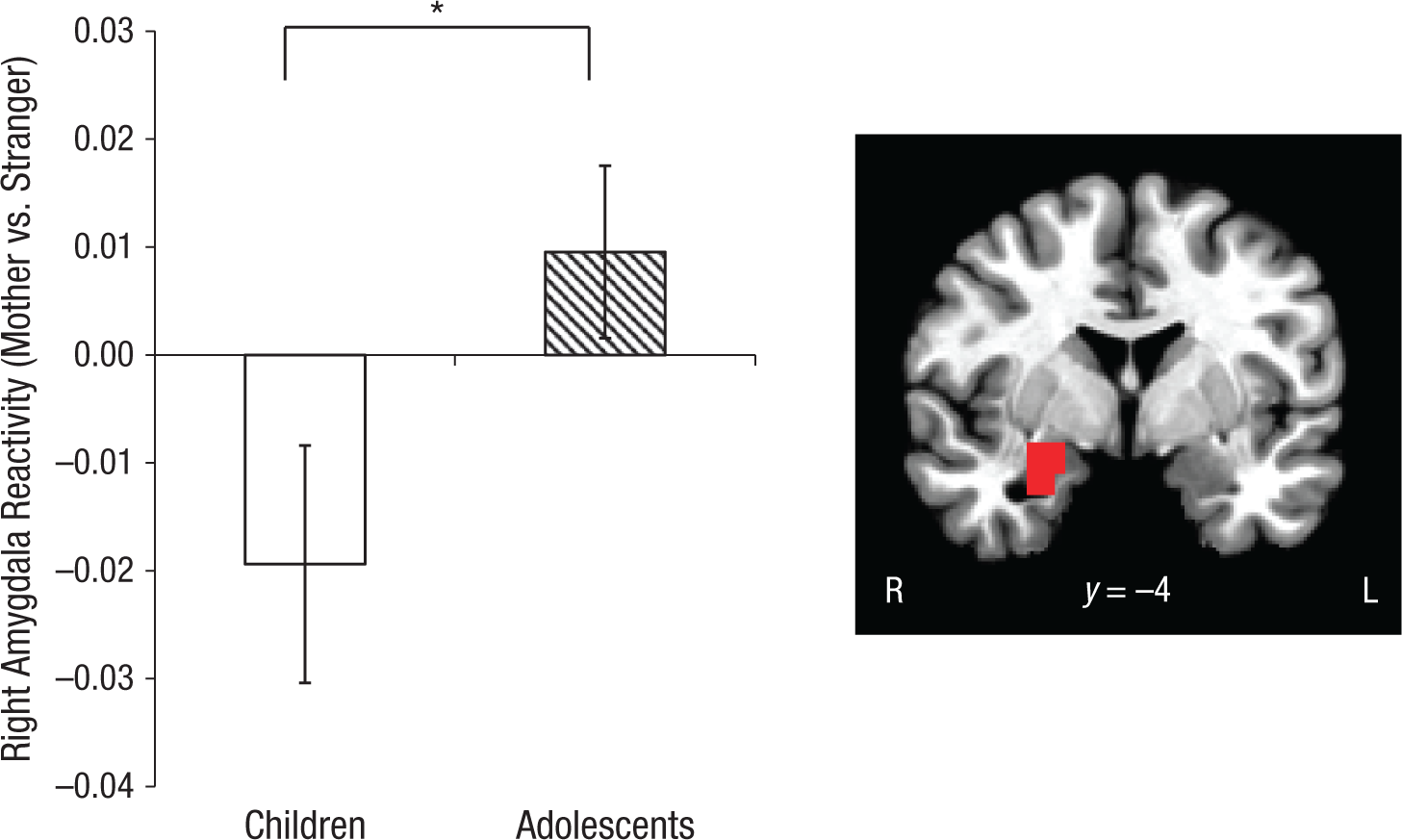
Results for the right amygdala. The graph shows mean beta weights in the mother condition relative to the stranger condition, separately for children and adolescents. The asterisk indicates a significant difference between groups (p < .05). Error bars show ±1 SEM. The brain image shows the region from which the results were taken (Montreal Neurological Institute coordinate y = −4). R = right, L = left.
Effect of the mother on functional connectivity
A voxel-wise PPI analysis, with the seed region of the right anatomically defined amygdala, was conducted to examine how the mother and stranger tasks modulated functional connectivity between the amygdala and the mPFC. Results demonstrated a Condition × Age Group interaction for amygdala-mPFC functional connectivity, F(1, 51) = 4.737, p = .034, η2 = .085 (medium effect size). Whereas adolescents showed negative amygdala-mPFC functional connectivity (relative to baseline) that did not differ between the mother and stranger conditions, t(28) = 0.139, p = .89, children showed differential connectivity to the mother and stranger conditions, t(23) = 2.496, p = .02, η2 = .213 (large effect size; Fig. 2). Amygdala-mPFC functional connectivity in children was negative relative to implicit baseline (inversely correlated) only for mothers, t(23) = 2.52, p = .019, d = 0.51 (medium effect size), but was not significantly different from zero for strangers relative to implicit baseline, t(23) = 0.70, p = .494. Connectivity for mothers did not differ between children and adolescents, t(51) = 0.48, p = .634. For more information about connectivity valence and amygdala reactivity, see the Supplemental Material.
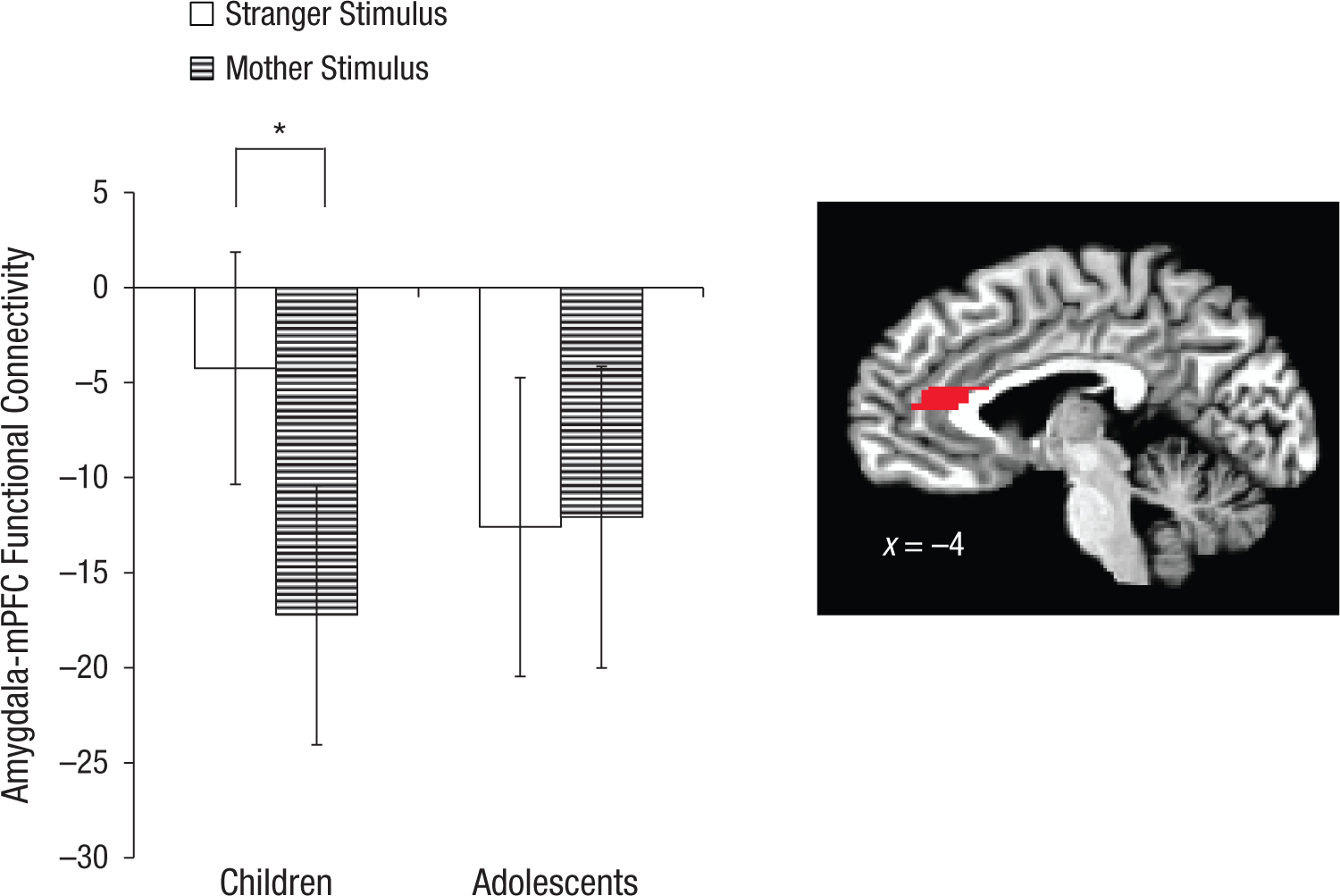
Mean connectivity between the amygdala and the medial prefrontal cortex (mPFC) as a function of age group and stimulus type. The asterisk indicates a significant difference between groups (p < .05). Error bars show ±1 SEM. The brain image shows the region from which the results were taken (Montreal Neurological Institute coordinate x = −4).
Amygdala-mPFC connectivity and individual differences
In order to understand the relationship between the maternally induced neural phenotype and individual differences, we grouped participants into those showing positive amygdala-mPFC connectivity relative to implicit baseline (n = 17; indicative of childlike neural response) and those showing negative connectivity relative to implicit baseline (n = 32; indicative of adultlike neural response). Participants who demonstrated negative amygdala-mPFC functional connectivity in response to mothers (vs. implicit baseline) had lower separation anxiety than those who exhibited positive connectivity in response to mothers, F(1, 48) = 6.12, p = .021, η2 = .21 (large effect size; Fig. 3a). As anticipated, adolescents had lower separation anxiety than children, F(14, 48) = 3.11, p = .008, η2 = .65 (large effect size). There was no interaction between age group and connectivity valence on separation anxiety, F(1, 48) = 1.11, p = .394. Participants with negative amygdala-mPFC functional connectivity in response to mothers (vs. implicit baseline) also had higher attachment security, as measured by the Security Scale’s subscale for youth’s reliance on their attachment figure in times of stress, F(1, 41) = 4.20, p = .047, η2 = .11 (medium-large effect size; Fig. 3b). There was no main effect of age group, F(1, 41) = 0.15, p = .70, and no interaction effect between age group and connectivity valence on reported attachment, F(1, 41) = 2.17, p = .149.
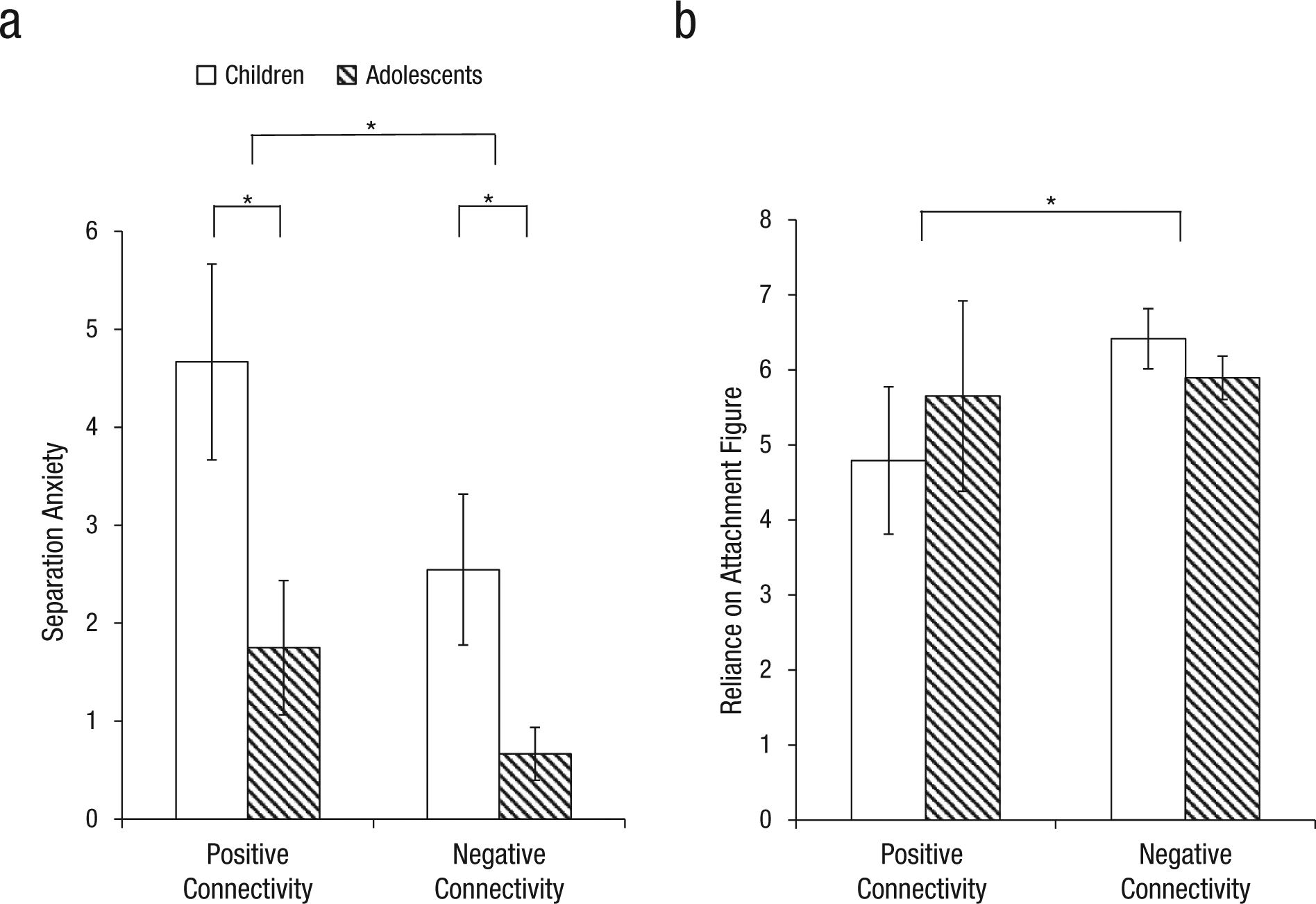
Mean separation-anxiety (a) and attachment-security (b) score as a function of connectivity between the amygdala and medial prefrontal cortex, separately for children and adolescents. An asterisk indicates a significant difference between groups (p < .05). Error bars show ±1 SEM. Analyses controlled for age.
Behavioral session: does maternal presence alter regulation in affective contexts?
Behavioral performance was analyzed using Bonferroni-adjusted alpha levels of .0167 (.05/3) per test to control for tests of false alarm rate, miss rate, and reaction time. There was a Condition (mother vs. stranger) × Age Group interaction, F(1, 46) = 6.35, p = .015, η2 = .12 (medium-large effect size) for false alarm rate, such that children (but not adolescents) exhibited fewer false alarms in the presence of mothers compared with strangers, t(21) = 2.24, p = .037, d = 0.35 (small effect size; Fig. 4). There were no main effects of or interactions with condition on false alarm rate or RT. This laboratory-based regulatory behavior was associated with maternal buffering of the amygdala during childhood (see the Supplemental Material). For information about performance on the mother/stranger fMRI task, see the Supplemental Material.
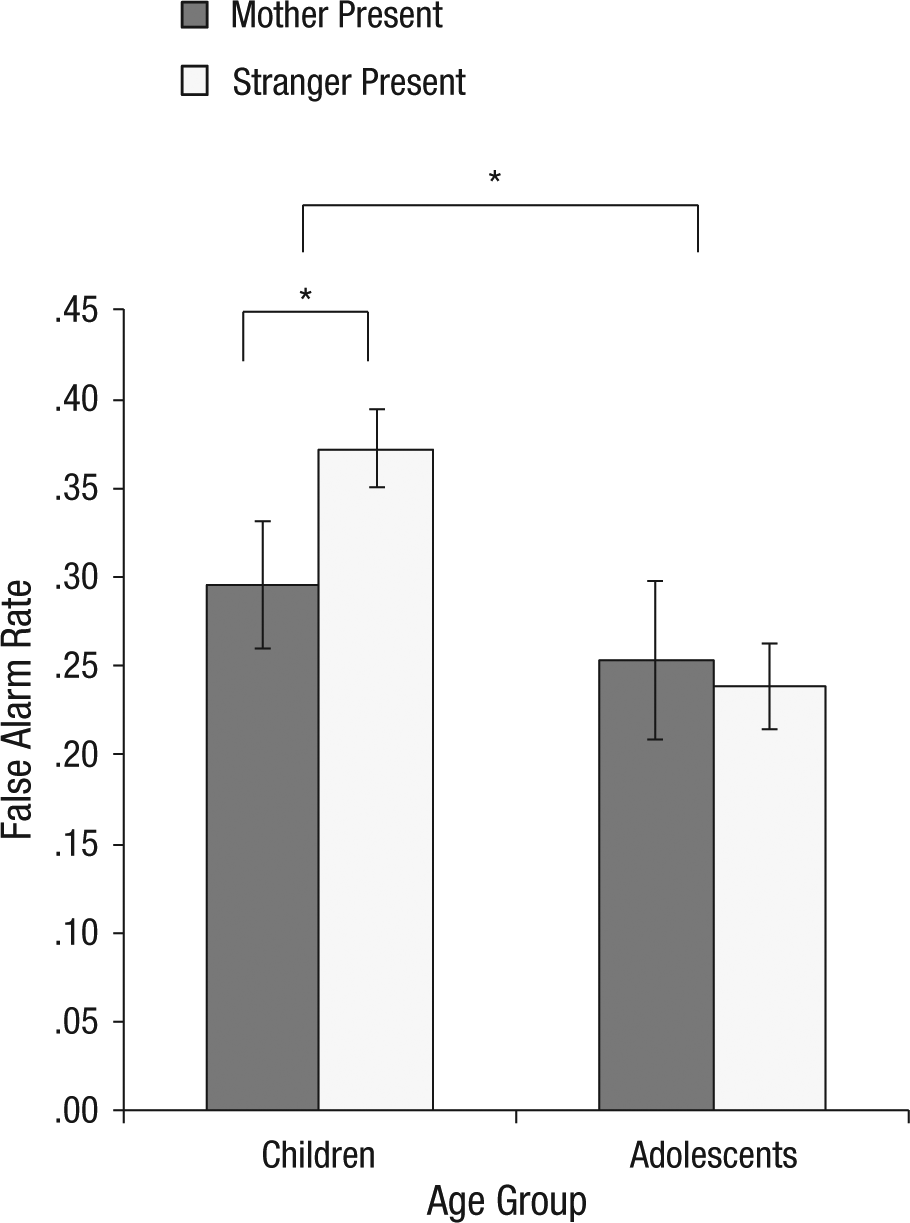
Results of the behavioral task: mean proportion of false alarms as a function of age group and whether the mother or a stranger was present during the task. An asterisk indicates a significant difference between groups (p < .05). Error bars show ±1 SEM.
Discussion
The present study examined the effects of mothers (via both presentation of their image and their physical presence) on amygdala-prefrontal circuitry and regulatory behavior in children and adolescents. The maternal stimulus induced a mature pattern of negative amygdala-prefrontal coupling and suppressed amygdala reactivity in children, whereas adolescents showed the expected pattern of mature connectivity to both the maternal and stranger conditions and did not show reduced amygdala reactivity to their mother. Moreover, children, but not adolescents, performed with better regulatory behavior in the presence of their mother than in the presence of a stranger, whereas adolescents’ performance was unaffected by maternal presence. These findings demonstrate that the maternal stimulus can modulate amygdala-mPFC circuitry and affect behavior, which supports the hypothesis that mothers serve as an external source of regulation through buffering effects on amygdala-prefrontal circuitry in childhood.
The finding that maternal stimuli buffer amygdala reactivity among children provides a neurobiological basis for the regulatory effects of caregiver presence early in life. Suppressed amygdala reactivity to the mother in this study parallels prior findings of maternal suppression of amygdala reactivity in rodents (Moriceau & Sullivan, 2006). Evidence shows that maternal presence also reduces corticosterone in rodents (Levine, 2001) and cortisol in nonhuman primates (Bayart, Hayashi, Faull, Barchas, & Levine, 1990) and humans (Gunnar & Donzella, 2002). Thus, the mother may buffer against stress through direct effects on the hypothalamic-pituitary-adrenal (HPA) axis, which acts on the developing amygdala. This potential mechanism has been explored in early childhood, and evidence suggests that social regulation through maternal buffering serves a regulatory function in humans (Gunnar & Donzella, 2002). In the present study, the regulatory effects of mothers on children were different from those on adolescents: The latter did not exhibit maternal neural modulation, which possibly reflects decreased behavioral relevance of caregivers as independence increases. Adults do not show differential amygdala activation to the face of their mother compared with the face of a stranger (Taylor et al., 2009), which suggests that childhood is a unique time during which the mother can modulate neurocircuitry. A similar period in rats has been identified as a “conditional sensitive period” for the amygdala (Moriceau & Sullivan, 2006, p. 655). Though understanding the function of maternal amygdala suppression requires further investigation in humans, participants with reduced amygdala reactivity to their mother made fewer false alarms in the presence of their mother, compared with participants without maternal amygdala suppression. Thus, maternal presence may act on amygdala circuitry to enhance regulatory behavior.
Adolescents exhibited negative amygdala-mPFC connectivity (vs. baseline) regardless of condition, a finding consistent with evidence for a switch from positive to negative amygdala-mPFC connectivity around the transition to adolescence (Gee, Gabard-Durnam, et al., 2013; Gee, Humphreys, et al., 2013). In children, connectivity to their mother was negative relative to baseline, but connectivity did not differ from baseline in response to a stranger’s face. Thus, children’s connectivity resembled the pattern of connectivity in adolescents only for the maternal condition. Given the known regulatory role of the mPFC in adults (Hariri et al., 2003; Kim et al., 2003), the observation in children of negative amygdala-mPFC connectivity with reduced amygdala reactivity to mothers suggests that the presence of the mother might buffer against amygdala reactivity through modulation of mPFC coupling with the amygdala. Though PPI analyses can compare how two tasks modulate functional connectivity, it is important to understand how each task modulates connectivity relative to implicit baseline. For adolescents, the mother and stranger conditions modulated amygdala-mPFC connectivity in similar ways; by contrast, children showed modulation by the mother (more negative than baseline) but not by strangers. Thus, the modulation of amygdala-mPFC functional connectivity by a maternal or stranger context changed with age. It is important to note the phasic nature of maternal neural modulation in the present study, compared with prior evidence suggesting that prolonged maternal separation accelerates amygdala-prefrontal development (Callaghan & Richardson, 2011; Gee, Gabard-Durnam, et al., 2013). Thus, while extended maternal deprivation may induce early development of mature connections with deleterious consequences, strong parent-child attachment may promote regulation through phasic amygdala-prefrontal modulation in the mother’s presence.
The finding that maternal buffering was present in children but absent in adolescents suggests that childhood may be a sensitive period for amygdala-prefrontal development. Identifying sensitive periods, during which the environment has particularly strong influences on neurodevelopment and thus lasting effects on behavior, is critical to understanding the developmental timing of neuroplasticity and how adult phenotypes are constructed. Sensitive periods have been shown in animal models of affective neurodevelopment (Callaghan & Richardson, 2011; Moriceau & Sullivan, 2006; Yang, Lin, & Hensch, 2012) but have been elusive in humans. The present study provides initial evidence for a sensitive period in human amygdala-mPFC development, during which the mother can modulate the system.
Physical maternal presence also affected performance on the affect-related regulation task, which has been used previously to measure behavioral regulation in an emotional context (Hare et al., 2008). Specifically, children made fewer false alarms in their mother’s presence than in a stranger’s presence, which may mean that the mother enables better behavioral regulation. Notably, children’s false alarm rate, which indexes a failure to regulate a response to an emotional stimulus, but not their miss rate, was modulated by maternal presence. These findings suggest that maternal presence does not improve general performance but specifically enhances affect-related behavioral regulation. Consistent with the increased behavioral relevance of the mother in childhood and our finding that access to maternal stimuli affected amygdala-prefrontal circuitry in children but not in adolescents, these results showed that only children performed differently in the maternal context.
Within children, individual differences in maternal amygdala-prefrontal modulation related to behavior. Those children who benefited from maternal buffering against amygdala reactivity were able to better regulate their affect-related regulatory behavior than children whose amygdala reactivity was not reduced in the presence of their mothers. Moreover, the adolescent-like neural phenotype elicited by maternal stimuli may reflect positive aspects of the parent-child relationship. Specifically, children who displayed negative connectivity to their mother (relative to implicit baseline) reported lower separation anxiety and more secure attachment than children who displayed positive connectivity to their mother. It may be that the nature of a secure attachment relationship results in the mother’s ability to regulate amygdala-prefrontal circuitry in children.
Though the fMRI task lacked an explicit demand for emotion regulation, it provides a unique opportunity to test the hypothesis that the mother buffers against amygdala reactivity in childhood. Prior work suggests normatively heightened amygdala reactivity in childhood (Decety, Michalska, & Kinzler, 2012; Gee, Humphreys, et al., 2013; Swartz, Carrasco, Wiggins, Thomason, & Monk, 2014; Vink, Derks, Hoogendam, Hillegers, & Kahn, 2014). In novel situations (e.g., an fMRI scanner), young children can typically access their caregiver, who would buffer against developmentally normative high amygdala reactivity. Employing threat-related stimuli or acute stressors will be important to further test maternal buffering. Though mother/stranger physical presence was manipulated in the behavioral task, visual images were manipulated in the fMRI task. It may be that the mother’s physical presence would produce even stronger neural effects of maternal buffering. A limitation of our fMRI task is the lack of stimuli normed by independent raters for valence, arousal, or perceived age. Moreover, because we purposefully compared amygdala reactivity to mothers versus strangers, we cannot rule out the possible influence of facial familiarity. This possibility seems less likely, however, because the effect was absent in adolescents (who should have even greater familiarity than children with their mothers). Future research would benefit from the inclusion of a familiar nonmaternal control condition. Finally, because there was a wide age range in the present study, future studies that focus on a group of similarly-aged participants in different stages of puberty (e.g., Forbes, Phillips, Silk, Ryan, & Dahl, 2011) would help to disentangle influences of age and puberty. (See the Supplemental Material for information about gender and pubertal status in the present study.)
Caregivers are one of the most salient stimuli early in life and provide many types of regulation for developing youth, including emotion regulation. Our findings provide a neuromechanistic framework for how caregivers regulate children’s emotional reactivity. Specifically, maternal presence appears to buffer against amygdala reactivity and induce phasic modulation of amygdala-mPFC connectivity, reflecting positive attachment and reduced anxiety. Moreover, for children who experience neural regulatory effects of their mother, maternal presence enables enhanced regulatory behavior. Thus, caregivers may serve an external regulatory function while circuitry supporting emotion regulation develops in childhood. With age, this maternal effect may become internalized, which allows for independent regulation via mature neural systems. The present findings provide evidence for how emotion regulation is supported in humans prior to mature central-nervous-system development.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This project was supported by National Institute of Mental Health Grants R01MH091864 (to N. Tottenham) and P50MH078105, by the Dana Foundation (to N. Tottenham), and by National Science Foundation Graduate Research Fellowship awards to D. G. Gee and L. Gabard-Durnam.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.