
Abstract
Although responses to different stressors are sometimes assumed to be similar, recent research has demonstrated that certain types of stress, such as targeted rejection, are particularly potent. To test such associations in a chronic-disease model, we examined how noninterpersonal, interpersonal, and targeted-rejection major life events predicted changes in gene expression and symptom severity in 121 youths with asthma who were assessed every 6 months for 2 years. Youths who had recently experienced targeted rejection had lower messenger RNA expression for signaling molecules that control airway inflammation and obstruction (specifically, the glucocorticoid receptor and β2-adrenergic receptor) than youths who had not experienced targeted rejection. These associations were specific to targeted rejection and stronger for youths higher in subjective social status. Higher-status youths exposed to targeted rejection (but not other types of stress) also reported more asthma symptoms. These data demonstrate stressor-specific associations with molecular-signaling pathways and the severity of asthma, and they suggest that threats to the social self may be particularly deleterious.
Stress research has been heavily shaped by generality theories, which posit that biological responses are uniform across different stressors (Selye, 1973). Although influential, this perspective has been criticized by theorists who argue that, to be adaptive, stress responses must be sensitive to the heterogeneous demands of different situations (Kemeny, 2003; Mason, 1971; Weiner, 1992). The latter view is supported by evidence demonstrating that different stressful situations have distinct physiological signatures (Dickerson, Gable, Irwin, Aziz, & Kemeny, 2009; Dickerson & Kemeny, 2004), but this evidence is derived almost exclusively from brief laboratory studies. Consequently, at least two important questions remain unanswered: Does stressor specificity extend to stressful life events that people experience in the real world? If so, are there corresponding implications for health?
To address these questions, we examined how a specific stressor, targeted rejection, relates to the health of children with asthma. Targeted rejection is a unique stressor in that it entails the intentional rejection of a person by another person or group of people (Slavich, Thornton, Torres, Monroe, & Gotlib, 2009). Examples of targeted rejection include having a romantic partner end one’s relationship or being fired from a job. Compared with similar events with different social-psychological characteristics—for example, choosing to end a relationship or being laid off from work along with other employees—targeted rejection elicits depression more rapidly and provokes greater activity in the hypothalamic-pituitary-adrenal axis and the sympathetic nervous system, along with increased inflammatory activity (Slavich & Irwin, 2014; Slavich, O’Donovan, Epel, & Kemeny, 2010).
Targeted rejection’s links with these systems may be a vestige of evolutionary history. Humans have a fundamental need to belong (Baumeister & Leary, 1995), and social exclusion elicits biological responses (Dickerson & Kemeny, 2004). Theorists believe that these responses prepare the body to manage threats, including injuries and infections, that accompanied exclusion in socially hostile ancestral environments (Dickerson, Gruenewald, & Kemeny, 2011; Irwin & Cole, 2011; Slavich et al., 2010). Although adaptive in the ancestral context, and perhaps still adaptive today, these responses may compromise health when evoked repeatedly. For example, persistent inflammation is known to contribute to the development of many chronic diseases (Nathan & Ding, 2010).
Building on these observations, we examined how targeted rejection relates to health in a multiwave study of youths with asthma. We also compared the impact of targeted rejection with that of other similarly severe stressors. Indeed, targeted rejection may have an especially large impact during later childhood and adolescence. During these periods, social activities become increasingly important sources of identity development (Hansen, Larson, & Dworkin, 2003), and youths report increased affiliation with peers and concerns about their position within social networks (Larson & Richards, 1991; Spear, 2000). Asthma is ideal to study among children and adolescents because it is well understood (Wright, 2011), is a major cause of disability and impairment, and poses a significant economic burden (Akinbami, Moorman, Garbe, & Sondik, 2009). The pathogenesis of asthma involves inflammation and airway obstruction that results in coughing, wheezing, shortness of breath, and chest tightness. Among the molecules that regulate airway inflammation and obstruction are the hormones cortisol, epinephrine, and norepinephrine, which act by binding to the glucocorticoid receptor (GR) and the β2-adrenergic receptor (β2-AR). Accordingly, we hypothesized that targeted rejection would be associated with decreased expression of both GR and β2-AR and a corresponding increase in the severity of asthma symptoms.
To examine the specificity of these associations, we compared the relative impacts of targeted rejection and a general category of interpersonal stressors. This enabled us to better isolate the unique effects of rejection. To evaluate the generality theories that have pervaded stress research (Selye, 1973), we also examined the impact of a broader category of noninterpersonal stressors. On the basis of the research discussed earlier, we hypothesized that targeted rejection would relate to biological and clinical outcomes in asthma more strongly than would similarly severe but contextually different interpersonal or noninterpersonal life events.
People possess unique resources that moderate their biological responses to stressful encounters and subsequent health outcomes (Weiner, 1992). Consequently, we also tested for possible moderation. One factor that is likely to moderate associations between targeted rejection and health is subjective social status—in this case, where youths believe they are situated within their peer hierarchy (Goodman et al., 2001). Although in the aggregate, higher-status individuals generally exhibit better health than their lower-status peers (Adler et al., 1994; Goodman, 1999), mounting evidence suggests that stature-threatening events take a disproportionate toll on these individuals. For example, in a study of young adults exposed to negative social evaluation, cortisol responses were significantly larger in high- than in low-status participants (Gruenewald, Kemeny, & Aziz, 2006). We recently replicated and extended these findings in adolescents, focusing on how social status moderates the effects of targeted rejection. Overall, adolescents who experienced targeted rejection showed increased expression of genes involved in immune-system regulation, but adolescents higher in subjective social status exhibited the largest targeted-rejection-related changes in gene expression (Murphy, Slavich, Rohleder, & Miller, 2013).
These findings are consistent with studies of nonhuman male primates showing that social instability evokes larger biological stress responses and more coronary artery disease in high- than in low-ranking animals (Manuck, Marsland, Kaplan, & Williams, 1995). These exaggerated responses are thought to reflect the disproportionate cost of social instability for high-status animals, in terms of access to resources and safety within the peer group. High-status adolescents who experience targeted rejection may incur similar costs; in addition, rejection may violate their sense of justice and undermine their self-esteem more than it does among low-status adolescents (Ramos, Correia, & Alves, 2014). Drawing on this work, we hypothesized that targeted rejection’s association with asthma outcomes would be moderated by subjective social status. Specifically, we expected that individuals who viewed themselves as higher in social status would have steeper declines in GR and β2-AR and greater increases in the severity of asthma symptoms after experiencing targeted rejection.
Method
Participants and procedures
Data were collected as part of a longitudinal study of youths with asthma. Participants were recruited using newspaper advertisements and flyers posted at schools and community centers and from asthma clinics in and around Vancouver, British Columbia. To be eligible, individuals were required (a) to be 9 to 18 years old, (b) to be fluent in English, (c) to have received a diagnosis of asthma from a physician, (d) to be free of acute respiratory illnesses at the time of their first visit (participants who were suffering from an acute illness at the time of their subsequent study visits were rescheduled), (e) to be medically stable (defined as not having had a course of oral steroids for the 2 weeks preceding their first visit), and (f) to have no chronic psychiatric or physical illnesses other than asthma. A total of 121 individuals met these criteria and were enrolled in the study (83% had allergic asthma). The project was approved by the University of British Columbia Research Ethics Board. At least one parent provided written consent for each youth, and youths provided assent (see Table 1 for an overview of the sample; see Tables S1 and S2 in the Supplemental Material available online for complete descriptive information for the sample at each visit and a correlation matrix of baseline study variables).
Descriptive Characteristics of the Sample (N = 121)
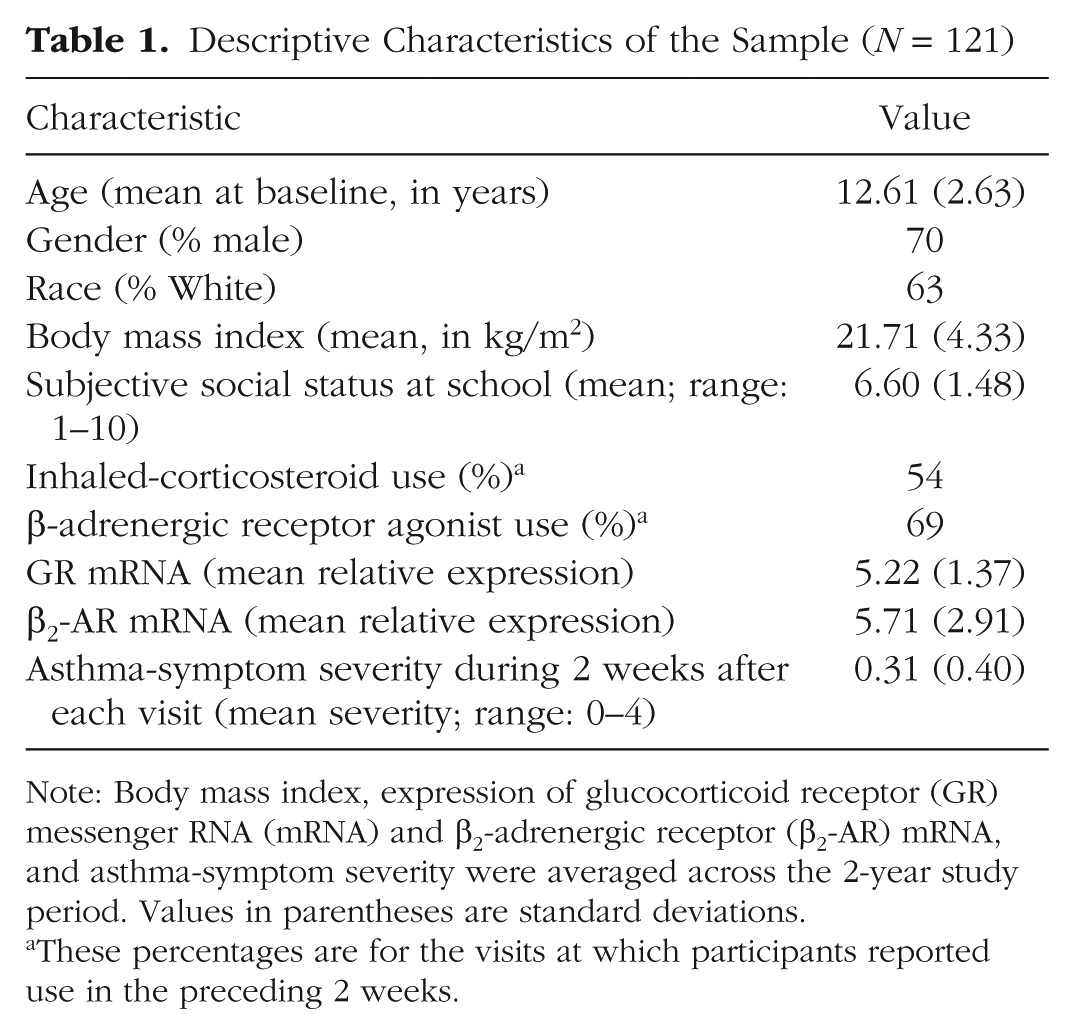
Note: Body mass index, expression of glucocorticoid receptor (GR) messenger RNA (mRNA) and β2-adrenergic receptor (β2-AR) mRNA, and asthma-symptom severity were averaged across the 2-year study period. Values in parentheses are standard deviations.
These percentages are for the visits at which participants reported use in the preceding 2 weeks.
Participants came into the laboratory every 6 months for 2 years, which provided up to five waves of data. At each wave, they completed questionnaires, were interviewed, and underwent a blood draw. After each visit, individuals reported on their asthma symptoms daily for 2 weeks. Overall, 104 participants (86%) completed at least three waves, and the majority of participants (n = 86, 71%) completed all five waves. Analyses were based on all available data.
Life stress and targeted rejection
We used the Life Stress Interview (Hammen, 1991) to identify the different types of life events that each participant experienced during the study period. Trained interviewers probed participants to understand the precise nature and contextual features of the stressors they had experienced during the 6 months before each study visit. The interviewers presented this information to a team of at least three independent raters, being careful not to disclose details that could have biased the ratings, such as participants’ emotional responses to the events. The rating team then made a consensus judgment regarding the severity of each event, basing their judgment on the degree to which the event would be expected to affect a typical person, given a similar context. Ratings were made in half-point increments on a scale from 1 (no negative impact) to 5 (severe impact).
Research has shown that the impact of stressful life events follows a threshold model and that there is a distinction between major and minor life events (Monroe & Simons, 1991; Monroe, Slavich, & Georgiades, 2009). Most researchers who have used the Life Stress Interview to conduct this research have coded events falling at or above the midpoint of the severity rating scale (i.e., 2.5) as major events (e.g., Miller & Chen, 2006; Murphy et al., 2013; Slavich, Tartter, Brennan, & Hammen, 2014; Uliaszek et al., 2012; Vrshek-Schallhorn et al., 2013). We followed this empirical precedent and standard practice in the present study by focusing on life events that were rated 2.5 or higher in severity. These events were subsequently coded as interpersonal or noninterpersonal by author M. L. M. Murphy; interpersonal events were defined as events that mainly involved difficulties with another person or group of people. Finally, interpersonal events were also coded for targeted rejection by author M. L. M. Murphy, according to criteria outlined by Slavich et al. (2009). Specifically, an interpersonal event was judged to involve targeted rejection if it (a) happened primarily to the participant, (b) involved the rejection of the participant by another person or group, (c) involved a clear intent to actively reject the participant, (d) directly affected the participant, and (e) resulted in the severing of a social bond between the participant and the other person or group.
For this project, we conducted more than 500 interviews, from which we identified 139 major life events in three mutually exclusive categories: 51 were identified as noninterpersonal major life events, 71 were identified as interpersonal major life events, and 17 were identified as targeted-rejection major life events. Of the targeted-rejection events, 7 involved a romantic breakup, 6 involved problems with friends or other peers at school, and 4 involved problems at school, such as expulsion (for examples of these events, see Table S3 in the Supplemental Material). The majority of these targeted-rejection major life events (n = 15) were experienced by different participants, and the maximum number of targeted-rejection life events experienced by any participant was 2.
Although working with a larger pool of events would be ideal, targeted rejection is a stressor that appears to occur relatively infrequently. In a previous study using a separate, independent sample of adolescents, we conducted more than 750 interviews, from which we identified 165 major life events (Murphy et al., 2013). Of these, 20 met criteria for targeted rejection. The base rates in the current and previous work are consistent with those reported originally by Slavich et al. (2009), who examined associations between targeted rejection and onset of depression.
A randomly selected subset of events (n = 10; 5 events originally coded as interpersonal, 5 events originally coded as noninterpersonal) was independently evaluated by author G. M. Slavich, who was blind to both the original ratings and the number of life events coded as either interpersonal or noninterpersonal. This evaluation was performed to examine the reliability of coding events as interpersonal. The same procedure was followed to examine the reliability of coding for targeted-rejection events. Author G. M. Slavich evaluated a subset of events to establish reliability (n = 10; 5 events originally coded as targeted rejection, 5 events originally coded as either interpersonal or noninterpersonal, but not targeted rejection). Agreement between coders was excellent for both categories (κ = 1.00 for interpersonal events; κ = .80 for targeted rejection).
Subjective social status
Participants’ subjective social status within their peer groups was assessed at baseline using the MacArthur Scale of Subjective Social Status, Youth Version (Goodman et al., 2001). Participants were shown a picture of a ladder with 10 rungs and were asked to indicate where they felt they fit on the ladder compared with their peers at school. They were told that higher rungs corresponded to people in their school who were respected, had friends, and did well academically, whereas lower rungs corresponded to peers who were not respected, did not have friends, and received poor grades.
Asthma symptoms
Participants reported their asthma symptoms for 2 weeks after each study visit. At waking, they reported the extent to which they had experienced coughing, wheezing, chest tightness or pain, and shortness of breath during the previous night. At bedtime, they reported the extent to which they had experienced these same symptoms during the day. The rating scale ranged from 0 (none) to 4 (really bad). Symptom reports were averaged across the 2 weeks.
GR and β2-AR
Whole blood (2.5 ml) was collected from antecubital veins at each visit to measure expression of messenger RNA (mRNA) for the α isoform of GR and for β2-AR in leukocytes. Total RNA was extracted from blood using PAXgene Blood RNA Kits (PreAnalytix, Hombrechtikon, Switzerland). Real-time reverse transcription-polymerase chain reaction was carried out using commercially available one-step assays from Applied Biosystems (TaqMan Gene Expression Assay developed in partnership with Applied Biosystems and based on RefSeq NM_000176 for GRα; TaqMan Gene Expression Assay Hs00240532 for β2-AR) on an Applied Biosystems Sequence Detection System (Applied Biosystems, Foster City, CA). Using the delta-cycle-threshold method, values of each target mRNA were adjusted for expression of the housekeeping gene 18S, which was measured in parallel (TaqMan Gene Expression Assay Hs99999901). Results were expressed as relative quantities of mRNA; higher values indicated greater expression of the target mRNA relative to the sample range. Each unit difference in relative quantity indicated a twofold difference in expression.
Why are GR and β2-AR of specific interest? As described earlier, asthma is characterized by inflammation and obstruction of the airways, brought about by exaggerated physiological responses to allergens, infections, and other stimuli. Normally, the body uses hormones it produces endogenously to help keep these processes in check. Cortisol regulates inflammation by acting on GRs located inside cells of the immune system, and the catecholamines epinephrine and norepinephrine regulate the tone of the airways by acting on β2-AR located on smooth muscle cells in the lungs. Dysregulation of these signaling pathways is a central pathological feature of many cases of asthma, which is why physicians often treat asthma with medications that target these receptors. Corticosteroids are the principle controller medication for many patients with asthma, because they help reduce inflammation by acting on GRs inside cells of the immune system. β2-AR agonists, in turn, serve as rescue medications for many patients with asthma when they are in the midst of an acute attack. These agents widen the airways and ease breathing by binding to β2-ARs on airway smooth muscle cells; they also act on immune cells in ways that reduce airway inflammation.
Alternative explanations
Exposure to major life events varies across demographic groups, as do asthma outcomes. To evaluate potential demographic confounds, we collected data on age, gender, and race (coded as White or other), and included these variables as covariates in analyses. Major life events may also lead to weight gain, or vice versa, and obesity affects inflammatory processes and asthma outcomes (Mokdad et al., 2003; Pirkola et al., 2010). To evaluate the possibility of confounding by obesity, we measured body mass index (BMI) at each visit. BMI was highly stable across the study (intraclass correlation coefficient = .90), so we used average BMI as a covariate. In addition, recognizing that life events may alter use of asthma medications and that these medications directly modulate physiological processes related to asthma, we assessed the number of days that participants reported using the two most common categories of asthma medications (i.e., inhaled corticosteroids or β2-AR agonists) during the 2 weeks before each visit. This enabled us to test whether life events were related to health outcomes, above and beyond the effects of medication use. 1 Finally, to evaluate whether links between life-event exposure and mRNA levels were confounded by variations in the composition of the circulating leukocyte pool, we covaried cell subset numbers (i.e., for monocytes, neutrophils, and lymphocytes) obtained from a complete blood count with differential at each visit (ADVIA 70 Hematology System; GMI Inc., Holliston, MA).
Analytic approach
Analyses were based on all available life-stress data, gene-expression data (specifically, levels of GR and β2-AR mRNA), and asthma-symptom levels assessed up to five times over a 2-year period. We also assessed participants’ subjective social status at baseline. Our predictors of interest included participants’ baseline subjective social status and whether they had experienced a noninterpersonal, interpersonal, or targeted-rejection major life event during the 6 months preceding each of the five study visits. The main outcomes of interest were within-person changes in levels of GR and β2-AR mRNA expression, which were measured at each study visit, as well as within-person changes in participants’ asthma symptoms, which were assessed daily over a 2-week period after each study visit and then averaged over that period to form an index of asthma-symptom severity at each time point.
To test our hypotheses, we evaluated a series of multilevel models using Hierarchical Linear and Nonlinear Modeling (HLM) software (Version 6.08; Raudenbush, Bryk, & Congdon, 2004). For the within-person portion of the models (Level 1), in separate equations, 2 the outcome variable of interest was predicted by within-person changes (i.e., the variables were person centered) in the occurrence of either (a) a noninterpersonal major life event, (b) an interpersonal major life event, or (c) a targeted-rejection major life event. Time since baseline (in months), inhaled-corticosteroid use, β2-AR agonist use, and a random intercept term were also included at Level 1 in all models. For the between-participants portion of the models (Level 2), the Level 1 slopes and intercept were predicted by individual differences in subjective social status as well as age, gender, race, and BMI. Subjective social status, age, and BMI were mean centered. The two key coefficients of interest in these analyses were (a) the Level 2 intercepts for the stress variables, representing the extent to which the occurrence of each type of life event was associated with within-person changes in the outcome of interest, and (b) the cross-level interaction between subjective social status and stress occurrence. A significant interaction would indicate that subjective social status moderated the strength of the association between within-person changes in the occurrence of a major life event and the outcome of interest.
Simple-slopes analyses for the cross-level interactions at the mean (6.60) and 25th percentile (5.99; i.e., lower-status individuals) and 75th percentiles (7.99; i.e., higher-status individuals) of the sample distribution of subjective social status were calculated using techniques outlined by Preacher, Curran, and Bauer (2006). Finally, multilevel-model effect sizes, ds, were obtained using methods described by Tymms (2004). Table S4 in the Supplemental Material contains the regression coefficients, standard errors, and p values for each variable in the models.
We use bar graphs to depict associations between exposure to a recent targeted-rejection major life event and the outcomes of interest for participants reporting different levels of subjective social status. The bars in these graphs do not represent means; rather, they represent the predicted values of the dependent variables, plotted from the multilevel-model regression coefficients. Therefore, the predicted values shown in the bar graphs are based on all available data.
Results
Comparing the severity of different types of major life events
Asthma-related outcomes could be more strongly related to targeted-rejection life events than to interpersonal and noninterpersonal life events simply because targeted-rejection events are more severe than the other two types of life events. To address this possibility, we compared the objective team-rated severity ratings of noninterpersonal, interpersonal, and targeted-rejection major life events. We found no evidence of differences in severity across these categories of events (noninterpersonal: M = 3.0, SD = 0.52; interpersonal: M = 3.0, SD = 0.53; targeted rejection: M = 3.1, SD = 0.57), F(2, 138) = 0.47, p = 0.63. Consequently, any differences in the associations of these three types of life events with asthma-related outcomes are probably due to differences in the events’ underlying social-psychological characteristics as opposed to differences in their basic stressfulness or severity.
Major life events, subjective social status, and GR mRNA
We found significant associations between within-person changes in the occurrence of a targeted-rejection event and relative expression of GR mRNA (see Fig. 1). As predicted, participants’ GR mRNA expression was significantly lower when they had experienced targeted rejection within the preceding 6 months than when they had not experienced targeted rejection (b = −1.62, SE = 0.65, p = .01, d = −0.42). In addition, individual differences in subjective social status moderated the strength of the association between targeted rejection and GR mRNA (b = −0.46, SE = 0.19, p = .02). Simple-slopes analyses indicated that after a recent targeted-rejection major life event, GR mRNA expression declined more sharply in higher-status individuals (b = −2.26, SE = 0.78, p = .004, d = −0.59) than in lower-status individuals (b = −1.34, SE = 0.62, p = .03, d = −0.35). (Subjective social status was included in all models as a centered variable with a mean of 0; therefore, at that mean, the association between targeted rejection and GR mRNA was identical to the association between within-person changes in targeted-rejection events and the relative expression of GR mRNA.) This pattern remained in a separate model that adjusted for variations in the distribution of circulating leukocytes (b = −1.77, SE = 0.65, p = .008, for the association; b = −0.51, SE = 0.19, p = .007, for the interaction).
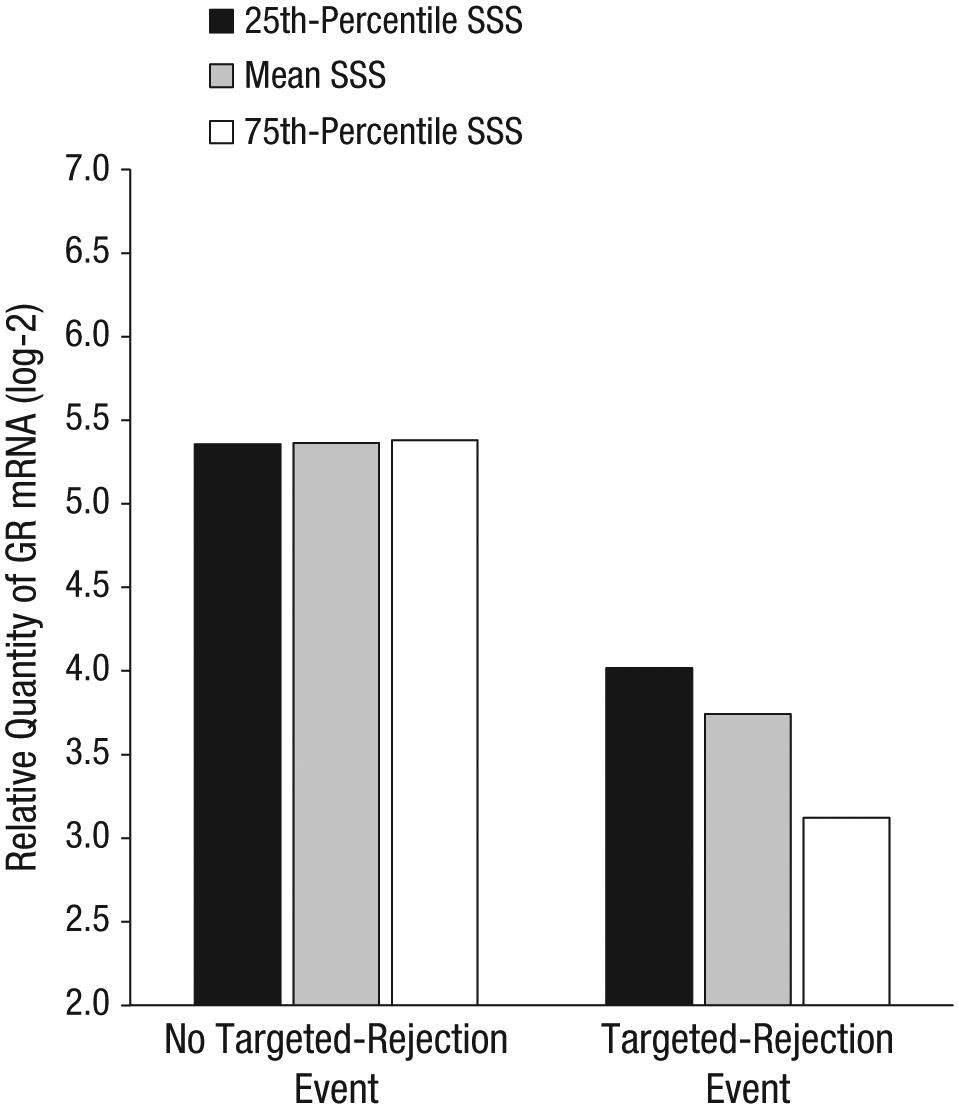
Predicted relative expression of glucocorticoid receptor (GR) messenger RNA (mRNA) after occurrence or nonoccurrence of a recent targeted-rejection major life event for participants at three levels of subjective social status (SSS). Individual bars represent the point estimates (not observed means) for relative expression of GR mRNA based on multilevel-model regression coefficients (n = 121).
We conducted parallel analyses to examine whether these patterns held for other similarly severe stressors. However, we found no significant associations between expression of GR mRNA and within-person changes in the occurrence of noninterpersonal (b = 0.82, SE = 0.44, p = .07, d = 0.36) or interpersonal (b = 0.21, SE = 0.53, p = .70, d = 0.01) major life events. Furthermore, individual differences in subjective social status did not moderate associations between expression of GR mRNA and noninterpersonal (b = −0.15, SE = 0.14, p = .31) or interpersonal (b = −0.15, SE = 0.11, p = .15) major life events. These findings are consistent with the notion that associations between major life events and expression of GR mRNA are specific to targeted rejection.
Major life events, subjective social status, and β2-AR mRNA
We also found a significant association between within-person changes in the occurrence of targeted-rejection events and relative expression of β2-AR mRNA (see Fig. 2). As predicted, participants’ β2-AR mRNA expression was significantly lower when they had experienced targeted rejection within the preceding 6 months than when they had not experienced targeted rejection (b = −3.20, SE = 1.14, p = .006, d = −0.51). Furthermore, individual differences in subjective social status moderated the strength of the association between targeted rejection and β2-AR mRNA (b = −0.59, SE = 0.30, p = .05). Simple-slopes analyses revealed that after a recent targeted-rejection major life event, β2-AR mRNA expression declined more in higher-status individuals (b = −4.01, SE = 1.15, p = .001, d = −0.64) than in lower-status individuals (b = −2.84, SE = 1.19, p = .02, d = −0.45). This pattern of findings remained mostly intact in a separate model that adjusted for variations in the distribution of circulating leukocytes (b = −2.46, SE = 1.26, p = .05, for the association; b = −0.55, SE = 0.30, p = .07, for the interaction).

Predicted relative expression of β2-adrenergic receptor (β2-AR) messenger RNA (mRNA) after occurrence or nonoccurrence of a recent targeted-rejection major life event for participants at three levels of subjective social status (SSS). Individual bars represent the point estimates (not observed means) for relative expression of β2-AR mRNA based on multilevel-model regression coefficients (n = 121).
Do other comparably severe stressful life events show similar associations with changes in expression of β2-AR mRNA? We found no associations between expression of β2-AR mRNA and within-person changes in the occurrence of noninterpersonal (b = 1.03, SE = 0.91, p = .20, d = 0.28) or interpersonal (b = −0.16, SE = 0.96, p = .87, d = −0.05) major life events. Likewise, subjective social status did not moderate the strength of the association between β2-AR mRNA expression and noninterpersonal (b = −0.41, SE = 0.27, p = .13) or interpersonal (b = −0.11, SE = 0.17, p = .52) major life events. These findings are consistent with the notion that associations between major life events and expression of β2-AR mRNA are specific to targeted rejection.
Major life events, subjective social status, and asthma symptoms
There were no within-person associations between the occurrence of targeted-rejection events and asthma symptoms, which were measured during the 2-week period after each study visit (b = 0.07, SE = 0.10, p = .48, d = 0.11). However, there was a significant cross-level interaction with subjective social status (see Fig. 3; b = 0.18, SE = 0.04, p < .001). Simple-slopes analysis revealed that individuals with lower social status did not report targeted-rejection-related changes in asthma symptoms (b = −0.04, SE = 0.10, p = .72, d = −0.05). However, participants with higher social status who had experienced a recent targeted-rejection major life event reported significantly more asthma symptoms during the 2-week period after their study visit (b = 0.32, SE = 0.12, p = .008, d = 0.47) than participants who had not experienced such an event.
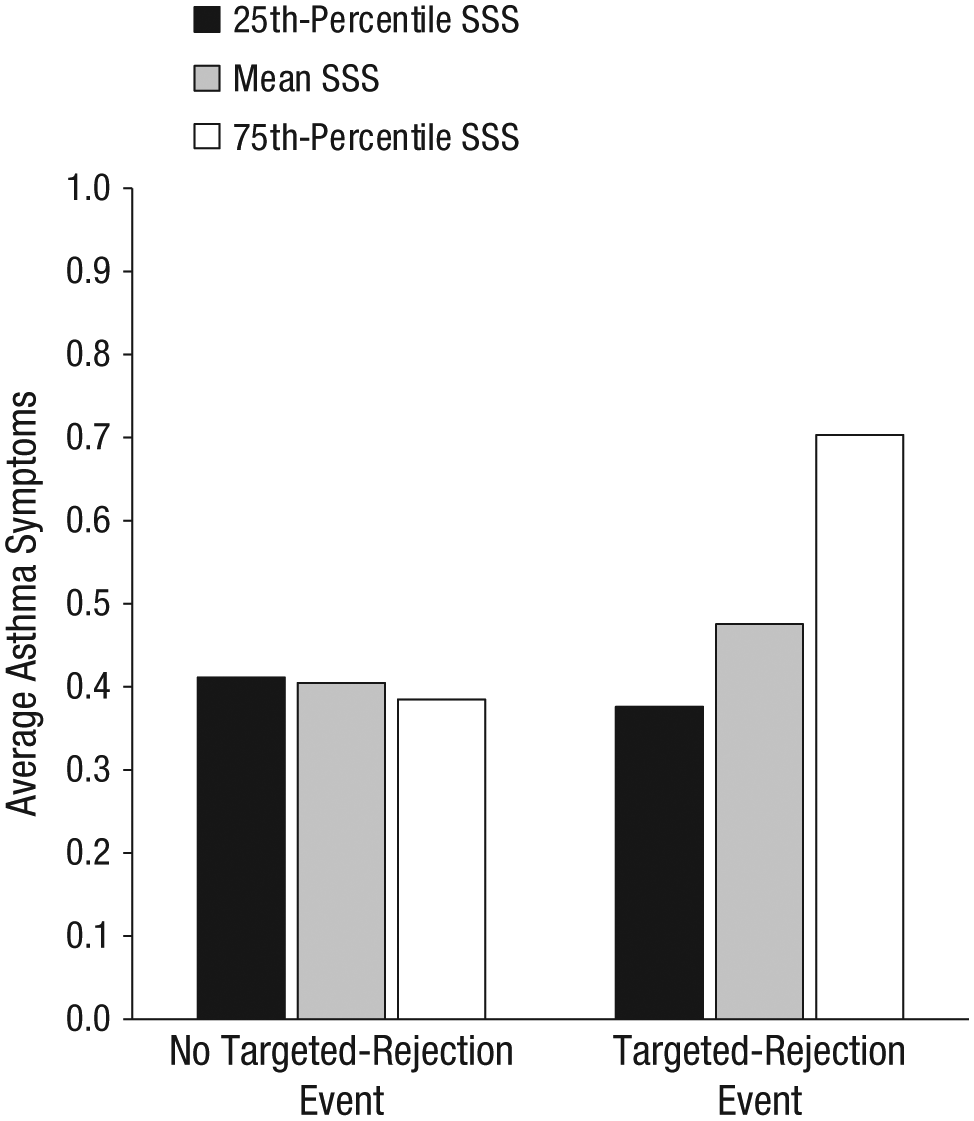
Average asthma-symptom scores after occurrence or nonoccurrence of a recent targeted-rejection major life event for participants at three levels of subjective social status (SSS). Individual bars represent the point estimates (not observed means) for asthma symptoms based on the multilevel-model regression coefficients (n = 121).
Results were consistent with the gene-expression findings described earlier for both interpersonal and noninterpersonal major life events: There were no associations between changes in asthma symptoms and the occurrence of noninterpersonal (b = −0.07, SE = 0.11, p = .51, d = −0.19) or interpersonal (b = 0.06, SE = 0.07, p = .36, d = 0.17) major life events. Likewise, subjective social status did not moderate associations between asthma symptoms and the occurrence of noninterpersonal (b = 0.06, SE = 0.03, p = .08) or interpersonal (b = −0.02, SE = 0.01, p = .14) major life events.
Mediation analyses
Next, we examined whether changes in the relative expression of GR and β2-AR mRNA mediated the association between targeted rejection and self-reported asthma symptoms. For a test of mediation to be warranted, there should have been associations between targeted rejection and both mRNA expression and asthma symptoms, and changes in mRNA expression should have predicted changes in asthma symptoms. However, we found no evidence that changes in asthma symptoms were predicted by within-person changes in expression of GR mRNA (b = 0.01, SE = 0.02, p = .76) or β2-AR mRNA (b = −0.01, SE = 0.01, p = .41). We found weak evidence that within-person changes in expression of GR mRNA predicted changes in asthma symptoms as a function of subjective social status (b = 0.02, SE = 0.01, p = .05) and no evidence that changes in expression of β2-AR mRNA predicted changes in asthma symptoms as a function of social status (b = 0.01, SE = 0.01, p = .13). Therefore, there was no evidence that changes in relative expression of GR and β2-AR mRNA mediated the association between targeted rejection and self-reported asthma symptoms.
Subjective social status and life-event exposure and severity
Finally, we examined whether the occurrence or severity of major life events that participants experienced varied as a function of their subjective social status. We used multilevel modeling to compute the odds of experiencing a major life event during the study as a function of individual differences in subjective social status. We found no evidence that subjective social status (entered into models as a continuous predictor variable) predicted differential exposure to noninterpersonal (odds ratio, OR = 0.99, b = −0.01, SE = 0.01, p = .21), interpersonal (OR = 0.99, b = −0.01, SE = 0.01, p = .10), or targeted-rejection (OR = 1.00, b = −0.004, SE = 0.01, p = .76) major life events. Next, we examined whether subjective social status predicted differences in the severity of the major life events experienced by participants. Again, we found no evidence for such an association for noninterpersonal (b = 0.002, SE = 0.003, p = .65), interpersonal (b = −0.003, SE = 0.003, p = .41), or targeted-rejection (b = 0.003, SE = 0.004, p = .44) major life events. Taken together, these findings suggest that the observed outcomes were not due simply to social-status-related differences in participants’ exposure to major life events or the severity of these events.
Discussion
In a 2-year multiwave study, we found consistent within-person associations between targeted rejection and the expression of signaling molecules that regulate airway inflammation and obstruction, in addition to associations between targeted rejection and asthma-symptom severity. These data are among the first to provide evidence that supports the specificity hypothesis in the context of major life events (Kemeny, 2003; Mason, 1971; Weiner, 1992); this hypothesis posits that different stressors elicit relatively distinct biological responses, with corresponding consequences for health. A key feature of targeted rejection is social-evaluative threat (Slavich et al., 2010), which is relatively unique among major life events in its capacity to engage neural pain and threat networks that amplify hypothalamic-pituitary-adrenal axis, sympathetic nervous system, and inflammatory activity (Dickerson & Kemeny, 2004; Kemeny, 2009). As we noted earlier, increased activation of these systems after social rejection or exclusion would have been adaptive in the ancestral context. But our data suggest that for modern patients with asthma, targeted rejection may undermine health, whereas other similarly severe stressors do not. In future research, it will be important to replicate these patterns and further delineate the neural, endocrine, and immune pathways that are engaged by targeted rejection and underlie its effects.
We also found that subjective social status moderated targeted rejection’s association with asthma outcomes. Higher-status youths exhibited the greatest declines in gene expression and the largest increases in symptom severity after targeted rejection. These findings replicate previous research on social status as a moderator (Gruenewald et al., 2006; Murphy et al., 2013) and relate this work to a clinical health outcome. Our findings also converge with research results showing that social instability (Manuck et al., 1995) and stature loss (e.g., Shively & Clarkson, 1994) have more pronounced health consequences for high-status than for low-status nonhuman primates.
This study is not without limitations. First, its design was correlational. Although data were collected prospectively across five waves and the statistical models adjusted for several possible confounding factors, definitive causative inferences cannot be made. At the same time, most analyses were based on within-person comparisons, and their results cannot logically be confounded by individual difference factors that typically threaten interpretations in correlational research. Regarding the moderation analyses, social status is unlikely to be distributed at random, and several factors (e.g., neuroticism, excessive reassurance seeking, narcissism, and rejection sensitivity) could potentially explain differential responses to targeted rejection in high- versus low-status individuals.
Second, although targeted rejection was associated with changes in signaling-molecule expression and everyday symptom reports, we found no evidence for mediation. This is not surprising given that many regulatory processes lie between gene expression and symptom presentation. Third, although levels of GR and β2-AR mRNA expression were derived from peripheral leukocytes, asthma is exacerbated by perturbations in the cells that make up the airways. Moreover, changes in the peripheral expression of the genes that code for these receptors do not necessarily guarantee corresponding changes in the airways. Additional research in which cells are harvested from the airways is thus needed to advance understanding of these processes in relevant tissue. Fourth, we focused only on youths with asthma who reported experiencing major life events. It will be important for future research to examine whether these findings generalize to other, less severe forms of rejection, and whether subjective social status also moderates associations between exposure to less severe rejection and physiological outcomes in a similar way. It will also be important to study whether these findings can be replicated across different chronic illnesses and developmental periods.
Another limitation concerns the relatively small number of targeted-rejection events in the sample. There are several reasons why this may be of concern. First, with such a low base rate, power to detect associations is low. Because we did find strong associations, this is not an issue for the current study. Second, fewer life events could lead to less precise estimates. However, this also does not seem to have been a problem for our study, because we found that targeted rejection was consistently associated with both biological and symptom outcomes, which themselves were weakly correlated. Furthermore, the standard errors associated with the multilevel-model parameter estimates were not large. We suspect that any problems with precision were offset by the high interrater reliability that we achieved using a well-validated, semistructured interview and strict coding guidelines. Finally, there could be concern that the findings are unreliable or spurious. Although this cannot be ruled out, targeted rejection showed consistent associations across multiple outcomes, and such relations were not present for other life events of similar severity. Furthermore, these gene-expression findings replicate those of another study (Murphy et al., 2013) in which an independent sample of adolescents reported similar rates of targeted rejection. Therefore, although it would be informative to study more targeted-rejection life events, thousands of interviews and concomitant assessments of asthma symptoms and gene expression would be needed to obtain a much larger sample of targeted-rejection events. Such a study would be extremely costly and is likely to be unfeasible.
Notwithstanding these limitations, this study offers new theoretical insights. Specifically, we found some of the first real-life evidence to support the stressor-specificity hypothesis in the context of health outcomes. These results challenge the uniformity assumption that has dominated theory and research on stress for much of the past century. In addition, the findings highlight the moderating role of subjective social status, extending from nonhumans to humans the observation that social instability takes a disproportionate toll on the health of high-ranking individuals. More broadly, these findings deepen knowledge of how social adversity influences health (Slavich & Cole, 2013).
Footnotes
Acknowledgements
We thank Katherine Ehrlich for her helpful feedback and insights during the early stages of this project. We also thank Shreya Agarwal, Nova Hou, Victoria Larsen, and Julia Wolloch for their assistance with preparing data for this manuscript.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by National Heart, Lung, and Blood Institute Grants R01-HL073975 and R01-HL108723 (to E. Chen), by Eunice Kennedy Shriver National Institute of Child Health and Human Development Grant R01-HD058502 (to G. E. Miller), and by a Society in Science – Branco Weiss Fellowship (to G. M. Slavich).
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.