
Abstract
Pain is a protective perceptual response shaped by contextual, psychological, and sensory inputs that suggest danger to the body. Sensory cues suggesting that a body part is moving toward a painful position may credibly signal the threat and thereby modulate pain. In this experiment, we used virtual reality to investigate whether manipulating visual proprioceptive cues could alter movement-evoked pain in 24 people with neck pain. We hypothesized that pain would occur at a lesser degree of head rotation when visual feedback overstated true rotation and at a greater degree of rotation when visual feedback understated true rotation. Our hypothesis was clearly supported: When vision overstated the amount of rotation, pain occurred at 7% less rotation than under conditions of accurate visual feedback, and when vision understated rotation, pain occurred at 6% greater rotation than under conditions of accurate visual feedback. We concluded that visual-proprioceptive information modulated the threshold for movement-evoked pain, which suggests that stimuli that become associated with pain can themselves trigger pain.
Keywords
Over the past three decades, the multidimensional nature of pain and nociception has been elucidated by research revealing that many factors from sensory domains (Moseley & Arntz, 2007), cognitive domains (Brooks & Tracey, 2005; Tracey, 2010; Wiech, Ploner, & Tracey, 2008), and emotional domains (Brooks & Tracey, 2005; Wiech & Tracey, 2009) modulate pain. Nonnociceptive information can both modulate and evoke pain (Acerra & Moseley, 2005; Arntz & Claassens, 2004; Derbyshire, Whalley, Stenger, & Oakley, 2004), which suggests that pain is evoked by information that exceeds a certain threshold but need not be from the nociceptive domain. The sensory cues capable of contributing to pain have not been well explored, although some authors suggest that any mix of information that leads the brain to conclude that the body is in danger may evoke pain (Arntz & Claassens, 2004; Merskey & Bogduk, 1994; Moseley, 2003; Price, 1999).
That suggestion is not surprising given what is known about other perceptual domains, in which sensory elements are combined into meaningful wholes. When perceiving a table, for example, people are aware of the unified whole rather than the individual colors, edges, and shapes (Goldstein, 2010; Hochstein & Ahissar, 2002; Weiten, 2007). Research investigating the principles underlying this sensory integration is plentiful, particularly with respect to vision (Wagemans et al., 2012), but such principles have received scant attention in the study of pain.
Pain serves a protective function, so it is natural to think that non-nociceptive sensory information might help to determine whether pain is an appropriate perceptual response. For example, nociceptive input from a small laceration may evoke pain only after visual information is added. In experiments, non-nociceptive cues contingently paired with nociceptive input (through classical conditioning) modulate the pain evoked by subsequent nociceptive stimulation (Atlas, Bolger, Lindquist, & Wager, 2010; Keltner, Furst, Fan, Rick, & Fields, 2006; Koyama, McHaffie, Laurienti, & Coghill, 2005). It is assumed that, after such pairing, the non-nociceptive cues become signals of body-related threat and thus join the suite of information used by the brain to determine whether pain is an appropriate protective perceptual response. Experimental evidence shows that the pain evoked by a nociceptive stimulus is affected by the meaning of both the nociceptive stimulus and other non-nociceptive stimuli that are presented at the same time (Arntz & Claassens, 2004; Moseley & Arntz, 2007).
Very few studies have investigated the relationship between non-nociceptive information and clinical pain. One study showed that people with complex regional pain syndrome experienced pain when what they saw in a mirror suggested that they were being touched, despite the absence of actual touch (Acerra & Moseley, 2005). However, in a group of people with neuropathic hand pain not caused by complex regional pain syndrome, such a procedure did not evoke pain (Krämer, Seddigh, Moseley, & Birklein, 2008). One might therefore suggest that non-nociceptive cues are important only in certain conditions. But if pain is information evoked—as we contend—then a threshold of relevant information (nociceptive, non-nociceptive, or both) must be reached before pain is evoked. This idea is untested, but if relevant non-nociceptive input did affect the amount of additional input required to evoke pain, such an effect would constitute, in essence, a change in pain threshold. Proprioceptive information about specific movements and body positions might have such an influence, especially when a vulnerable body part needs to be protected. For people with a neck injury, for example, specific proprioceptive information might predict nociceptive stimulation and thus contribute to defensive responses, including pain.
In the present study, we tested the hypothesis that proprioceptive information might contribute to pain. We examined whether altering visual-proprioceptive feedback during neck rotation would affect the position at which people suffering from long-standing pain experienced its onset (i.e., the threshold of movement-evoked pain). We hypothesized that pain would occur earlier when visual-proprioceptive information overstated real-world rotation and later when visual-proprioceptive information understated real-world rotation.
Method
Participants
Twenty-four individuals (6 men, 18 women; mean age = 45 years, SD = 15) volunteered to participate in this study. Sample size was determined a priori to enable detection of a small- to medium-sized effect (η p 2 = .04) with 80% power, conservatively assuming a 60% correlation among repeated measures. The average duration of complaint was 11 years (SD = 11; range: 2 months to 45 years), and the participants’ pain conditions resulted primarily from posture, tension, or repeated strain (n = 9); whiplash (n = 7); degeneration (n = 5); trauma (n = 2); and scoliosis (n = 1). Participants were mildly to moderately disabled (Neck Pain Disability Index score = 29%, SD = 13%). Participants were recruited through local physiotherapy clinics and were reimbursed AU$20 for their participation. Participants were excluded if they had pain-free neck rotation, were unable to tolerate repeated rotation to the position at which they experienced the onset of pain, had severely impaired vision, or were under the age of 18. Participants were also excluded if a physiotherapist had identified significant neurological impairments, such as sensory or motor deficits and easily provoked, constant, or progressive upper-limb dysesthesia. The protocol was approved by the Human Research Ethics Committee of the University of South Australia (Protocol Number 31537).
Stimulus material and apparatus
A virtual-reality (VR) technique known as redirected walking modulates visual-proprioceptive feedback by tracking real-world movement and then feeding this back into the virtual environment in an understated or overstated form. Rotation gain (the factor by which real rotation is translated to virtual rotation) can be manipulated such that virtual and physical rotation differ. This process creates the illusion of either more or less movement than is actually happening. Within certain limits, participants remain unaware of the manipulation (Steinicke, Bruder, Jerald, Frenz, & Lappe, 2008). We used a VR head-mounted display (HMD) designed for immersive VR environments (Oculus Rift; Oculus VR, Irvine, CA). The HMD shown in Figure 1 displayed a virtual world and recorded head movement using internal gyroscopes. Customized software was used to map each of six scenes to the virtual template and apply the selected rotation gains. The six scenes included four outdoor scenes (a park, a mountain, a countryside, and church grounds) and two indoor scenes (a dining room and a living room).

Virtual reality equipment. Participants sat in supportive chairs that prevented trunk movement (i.e., a seat belt around their shoulders) and wore headphones and head-mounted displays. A 360° (cylindrical) virtual template accommodated six virtual scenes.
Experimental design
We used a within-subjects, randomized, double-blinded, repeated-measures design. The distance from center position to the left or right position at which participants experienced the onset of pain (i.e., the pain-free range of motion) was quantified in three conditions. Virtual rotation was (a) 20% less than actual physical rotation (rotation gain = 0.8), (b) equal to actual physical rotation (rotation gain = 1), or (c) 20% greater than actual physical rotation (rotation gain = 1.2). The order of the three conditions was counterbalanced across participants, which created six possible orders in which conditions could be presented. Four participants were randomly assigned to each order. Participants were blinded to the experimental manipulation and study purpose, and the files relating to the three VR conditions were coded, thereby blinding the experimenter to the order of conditions. The measurement of neck rotation was automated, and data were extracted only after collection was completed.
Measurement
Pain-free range of motion for axial neck rotation was measured in degrees. Because participants stopped at the onset of pain in each trial, the distance from center position to the left or right position at which participants experienced the onset of pain was defined as the peak rotation for each trial. This measure was extracted from each automated trial output by a blinded assessor.
Protocol
To prevent changes in postural alignment and to isolate neck movement, we asked participants to sit in a supportive chair, and their torsos were fixed in place by a seat belt at the level of the shoulders (see Fig. 1). A laser pointer was affixed to the HMD; the dot it produced was used to mark the starting position on the wall as a physical reference point for zeroing of the gyroscopes between measures and conditions. Participants wore headphones that emitted white noise to counter any incidental noise that might inform head orientation and disrupt the illusion.
For each of the three conditions, participants were asked to rotate their heads slowly to the left and to stop at the onset of pain. They then returned their heads to the center, at which point the next trial was loaded, and the task was repeated on the right side. Each condition consisted of six measures of left rotation and six measures of right rotation. After each condition, participants were asked to rate the average pain intensity experienced for each rotation direction. This allowed for subsequent assessment of any overall differences in pain intensity between conditions. Pain was rated on an 11-point numerical scale (0 = no pain, 10 = the worst imaginable pain). To minimize the possibility that subjects would become aware of the different rotation settings and thus directly compare them, we used a 3-min interval between conditions. In addition, the six different VR scenes acted as a distraction from the actual study purpose and reduced the risk of participants’ anchoring their rotation to a previous visual cue within a VR scene. To assess blinding, at the end of the experiment we asked participants whether they noticed anything different between the three conditions.
Manipulation Check 1: setting boundaries for altered visual-proprioceptive feedback
To blind participants to this manipulation, we based the upper and lower limits of the rotation gain on the results of a pilot study. During this pilot study, an independent cohort of 9 healthy participants (7 men, 2 women; mean age = 32 years, SD = 12) were presented with a range of rotation-gain settings and asked to rotate their heads. They were to indicate when a difference between real and virtual rotation occurred by rating the observed rotation as slower than, equal to, or faster than the true physical rotation. We aimed to determine the rotation gain at which participants were more likely to judge the virtual and real rotation to be equal than they were to judge them to be different. As shown in Figure 2, the rotation gains that corresponded to these points were 0.72 and 1.18. As a result, our experimental gain settings were chosen to fall between 0.8 and 1.2, and additional controls were implemented to ensure that participants in the main study remained blinded.
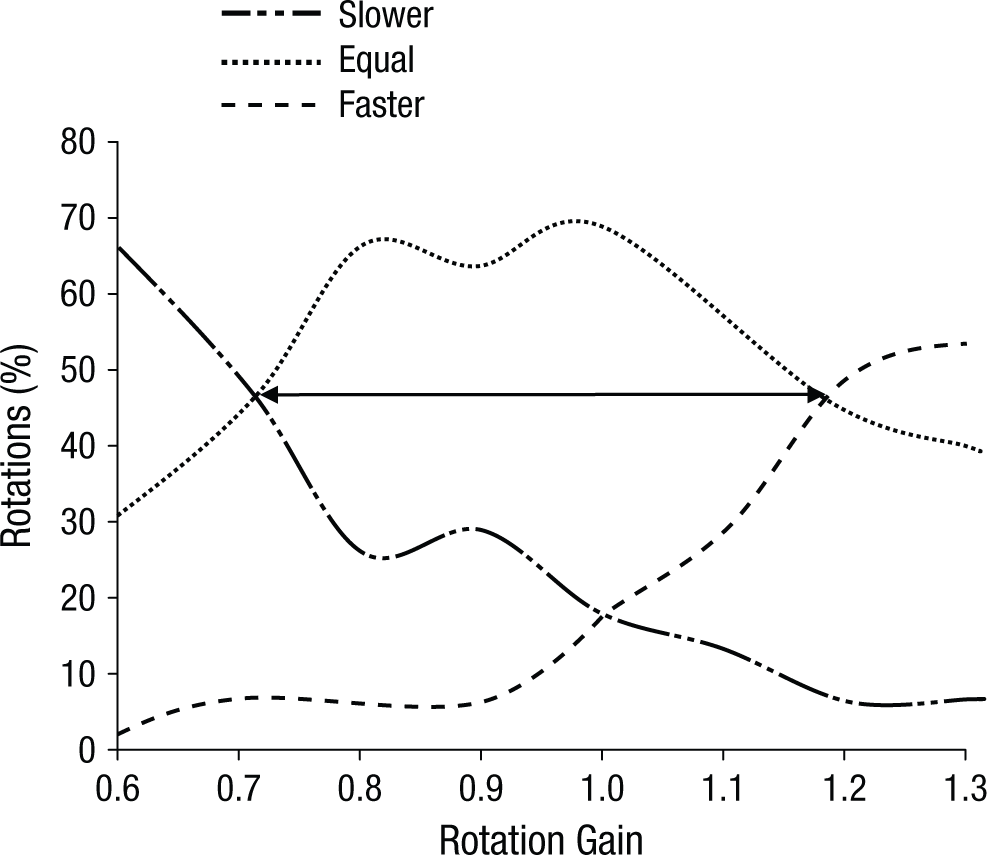
Results from the pilot study. The graph shows the percentage of rotations rated as slower than, equal to, or faster than the true physical rotation as a function of rotation gain. The double-headed arrow indicates the range of rotation-gain settings at which virtual movement speeds were more often perceived as equal to, rather than faster or slower than, real-world movement speeds.
Manipulation Check 2: reliability and validity of measurement
The reliability of the gyroscopic measurement of rotation was tested by attaching the HMD to a mechanical (goniometric) arm and testing its ability to repeatedly and accurately measure three set angles (20°, 40°, and 60°). Initial observation of repeated measures revealed that the measurement was highly precise over a small number of trials, but error gradually increased as the number of trials increased. Therefore, a protocol was developed that required the virtual compass to be zeroed every five trials. To further prevent accumulation of error, we refreshed the program between conditions. This protocol enabled rotation as measured by the HMD to be correlated with actual rotation, r = .994, and produced a high degree of precision in the HDM’s measurements of rotations of 20° (M = 19.8°, SD = 0.8), 40° (M = 39.6°, SD = 0.3), and 60° (M = 59.1°, SD = 2.5).
Data extraction and statistical-analysis overview
To test our main hypothesis (i.e., that visual information that overstates or understates true rotation can affect movement-evoked pain), we compared pain-free range of motion across the three conditions. We used repeated measures analysis of variance (ANOVA) with Bonferroni-corrected pairwise comparisons. To account for between-subject differences in range of motion, we transformed data for each participant to a proportion of the average range of motion demonstrated in the neutral condition. Alpha was set at p < .05, and we used Cohen’s guidelines (Cohen, 1988) to interpret the effect sizes η p 2 (.01 = small, .059 = medium, and .138 = large) and Cohen’s d (0.2 = small, 0.5 = medium, and 0.8 = large). As a manipulation check, each participant’s average movement-evoked pain for each condition was normalized to a proportion of their average across conditions. Normalized pain ratings were then compared among conditions using repeated measures ANOVA.
Results
Primary outcome: pain-free range of motion
The repeated measures ANOVA revealed a large overall effect of visual-proprioceptive feedback (condition) on pain-free range of motion F(2, 94) = 18.9, p < .001, η p 2 = 0.29. All pairwise comparisons were significant (ps < .01). As shown in Figure 3, when vision understated true rotation, pain-free range of motion was increased, and this was a medium-sized effect, p = .006, d = 0.67; when vision overstated true rotation, pain-free range of motion was decreased, and this was a large effect, p = .001, d = 0.80. Specifically, during visual feedback that understated true rotation, pain-free range of motion was increased by 6% (95% confidence interval, or CI = [2%, 11%]); during visual feedback that overstated true rotation, pain-free range of motion decreased by 7% (95% CI = [3%, 11%]). Therefore, our results show an overall effect of the manipulation of 13%.
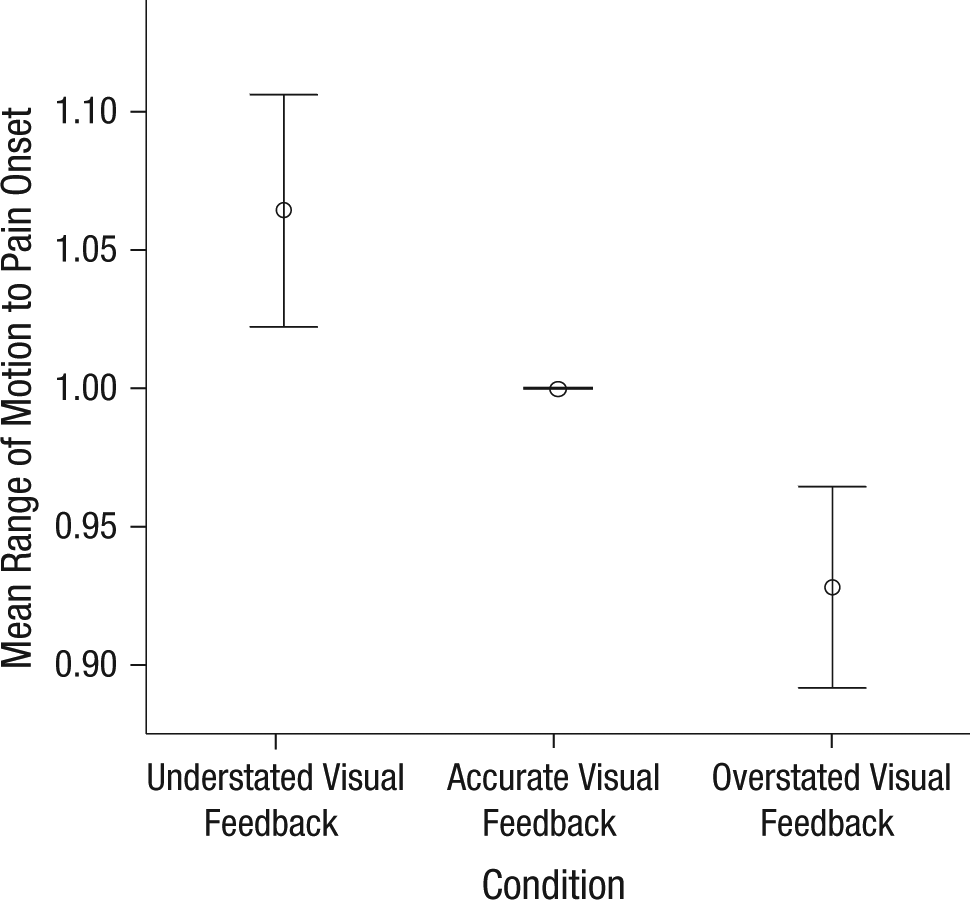
Mean range of motion to first onset of pain for the conditions of understated, accurate, or overstated visual feedback. Range of motion is presented as a proportion of the mean range of rotation for the accurate visual feedback condition. Error bars indicate 95% confidence intervals.
Pain intensity across conditions
The repeated measures ANOVA revealed no difference in pain intensity between conditions (p = .6). That is, the difference in rotation could not be explained by participants’ actually moving beyond or stopping short of their pain threshold, which strengthens our confidence in the main finding. In fact, Figure 4 hints at more pain in the condition in which visual feedback suggested more movement, further reinforcing the finding that the visual suggestion of more movement increases sensitivity.
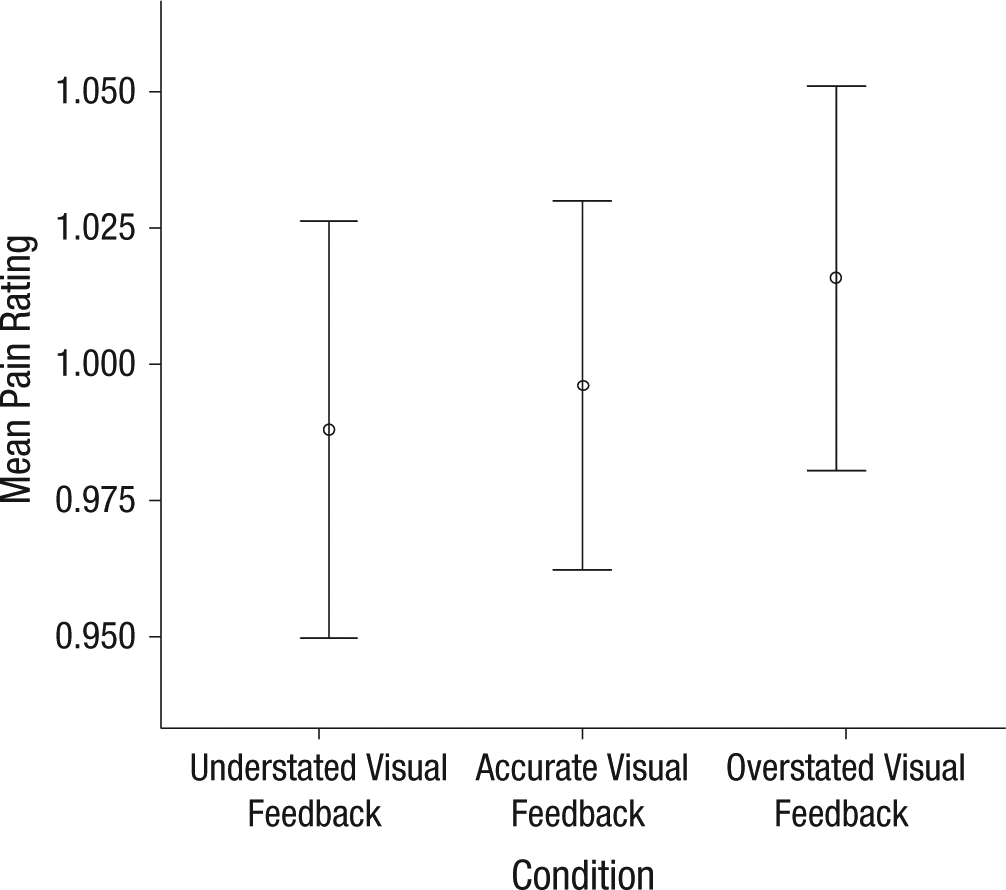
Mean pain ratings (normalized data) for the conditions of understated, accurate, or overstated visual feedback. Error bars indicate 95% confidence intervals.
Discussion
We examined how altering visual-proprioceptive feedback during neck rotation affected pain-free range of motion (the threshold of movement-evoked pain) in sufferers of long-standing neck pain. Using the information-based view of pain, we hypothesized that pain would occur with less head rotation when visual-proprioceptive information overstated true rotation (i.e., there would be a reduced pain threshold compared with that during accurate visual feedback), and that pain would occur with more head rotation when visual-proprioceptive information understated true rotation (i.e., there would be an increased pain threshold compared with that during accurate visual feedback). The hypothesis was clearly supported. The finding that visual-proprioceptive cues may contribute to a pain-evoking sensory suite is particularly relevant because chronic pain is most commonly provoked by particular movements and body positions.
Our results appear to be consistent with the view of pain as the perceptual result of the brain’s inference that body tissue is threatened (Arntz & Claassens, 2004; Merskey & Bogduk, 1994; Moseley, 2003; Price, 1999) and with related evidence of the relationship between experienced pain intensity and cues that imply threat to tissue (Arntz & Claassens, 2004; Atlas & Wager, 2012; Moseley & Arntz, 2007; Wiech et al., 2010). For example, a noxious cold stimulus evokes more pain if it is accompanied by a red light than if it is accompanied by a blue light (Moseley & Arntz, 2007). A noxious laser stimulus evokes more pain (and different cortical activation) if it is delivered to an area of skin thought by the participant to be thinner than normal than if it is delivered to skin thought to be normal (Wiech et al., 2010). Such examples offer compelling evidence that pain can be modulated if there is credible evidence of tissue danger, even if that evidence is not from the nociceptive domain.
The current results offer a new direction, however, because they show a shift in pain threshold rather than an increase in pain. This is important because most examples of amplification of pain can be explained by enhanced sensitivity within the nociceptive system (which is most notably termed central sensitization). We contend that our results (i.e., of a reduced pain-free range of motion compared with that during accurate visual feedback) are very unlikely to be due to central sensitization, which would be more likely to manifest in the opposite result—a greater pain increase in response to greater magnitude of movement. We contend that the most obvious explanation for the current results is associative learning (see also Moseley & Vlaeyen, 2014). That is, neck rotation involves a suite of motor, visual, and proprioceptive processes that, for the person with neck pain, becomes associated with nociceptive input, such that the nonnociceptive aspects of the sensorial suite are sufficient to trigger pain with or without the nociceptive component.
The effect clearly relies on visual input triggering some sort of threat mechanism, but this does not exclude the possibility that non-cortical mechanisms are involved. The most obvious candidate is the descending modulatory system, whereby brainstem nuclei (and other structures) exert both inhibitory and facilitatory influences over dorsal horn neurons (see Woolf & Slater, 2006, for review). According to modern models of pain, however, the evaluative processes that subserve descending modulation are grounded in those that subserve the production of pain itself. That is, pain can be considered to reflect the perceived need to protect body tissue; descending modulation can be considered a correction of spinal nociceptor activity (i.e., the danger message) that brings such activity in line with the brain’s evaluation of true danger (see Butler & Moseley, 2013; Fields, Basbaum, & Heinricher, 2006; Moseley, 2007). This understanding is analogous to that applied to motor control, whereby motor commands are corrected according to somatosensory and visual feedback (Sperry, 1950; von Holst, 1950). Such models are also relevant here, because it is also possible that the bogus visual feedback modulates proprioceptive sensitivity (Gandevia, Refshauge, & Collins, 2002), which in turn may modulate nociceptive input.
The pain-advancing effect of overstating rotation was greater than the pain-delaying effect of understating rotation, which indicates that visual feedback had a greater ability to reduce the pain threshold than to increase it. This finding fits the idea that perceptual error that delays the onset of pain can be costly and is consistent with inferential perceptual models (i.e., Bayesian) that include a cost function (Feldman, in press; Tabor, Catley, Gandevia, Thacker, & Moseley, 2013).
Although the current work was experimental in nature, it raises intriguing potential clinical implications. First, our results clearly suggest a reevaluation of how we interpret simple clinical tests such as movement-evoked pain. Such tests are widely held to reflect sensitivity of tissues and nociceptive pathways; that is, repeatable and stable thresholds for movement-evoked pain are considered to indicate a primary nociceptive driver of pain—the presence of tissue pathology (Jones & Rivett, 2004). However, our results suggest that this is a naive perspective. Therefore, it is not unreasonable to suggest that a VR setup such as that used here might play a role in the assessment of pain, to identify and quantify the role of nonnociceptive cues.
Second, if cues signaling danger amplify or indeed trigger pain, then these cues present a novel target for therapy. One way to extinguish the effect of such cues on pain might be to experientially dissociate them from pain. For example, the redirected-walking techniques used in this study can provide the experience of large movements, albeit illusory, but limit real-world movement and pain. This idea might also be relevant to the use of mirrors to generate visual illusory movements; mirror therapy is commonly used for conditions involving chronic limb pain, including phantom limb pain and complex regional pain syndrome (Bowering et al., 2012; Daly & Bialocerkowski, 2009; Ezendam, Bongers, & Jannink, 2009; Moseley, 2004b, 2006), but the idea of altering feedback as a method of disentangling non-nociceptive movement-related cues appears to have not been considered in the studies cited here.
The relationship observed here between potentially threatening information and movement-evoked pain might also provide insight as to why cognitive and behavioral interventions (e.g., education and exposure) that target fear related to perceived threat, including pain, also positively alter the relationship between movement and pain (Moseley, 2004a; Vlaeyen, de Jong, Geilen, Heuts, & van Breukelen, 2002). Although education, for example, may aim to convince patients that their pain is not a direct correlate of tissue stress, demonstrating this with real-time evidence that their pain depends on visually encoded movement rather than actual movement may have therapeutic power.
Further research using the current framework might exploit more immersive and multisensory VR, which may enable the delivery of more convincing and multimodal illusory evidence of danger to the body. Further studies could also investigate how visual-proprioceptive and other cues might acquire the ability to modulate/mediate pain (i.e., through associative learning) as well as investigate why the effect might persist or overgeneralize, and how it might be extinguished. Disentangling pain from nociception is a challenge that has been identified in experimental and cognitive psychology research (Moseley, Gallace, & Spence, 2012), but the methodology used here creates a basis from which this challenge might be addressed.
Conclusion
In people with neck pain, when visual-proprioceptive feedback overstates true neck rotation, pain-free range of motion is reduced compared with that during accurate visual feedback. When visual-proprioceptive feedback understates true neck rotation, pain-free range of motion is increased compared with that during accurate visual feedback. We conclude, then, that visual-proprioceptive information modulates pain thresholds during head rotation in people with neck pain. This has broad implications for our view of pain as an information-evoked response and supports further investigation of nonnociceptive contributions to long-standing pain. Furthermore, the methodology outlined here presents a new method for theoretical and experimental interrogation of pain and raises the possibility of novel assessment and therapeutic applications.
Footnotes
Acknowledgements
For assistance with recruitment, we thank Julie Peacock, Jonathon Schubert, William Kuang, and Ellie Magarey (Marion Physiotherapy); Di Wilson, Peter Roberts, Eva Boesch, and Toby Moen (Roberts Physiotherapy); and Jason Collins and Glen Kocher (Northcare Physiotherapy). We thank Anthony Ikiosoglous for acting as a blind assessor for data extraction.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by National Health and Medical Research Council (NMHRC) Principal Research Fellowship 1061279 (to G. L. Moseley), NHMRC Grant 1047317 (to G. L. Moseley), Fonds Wetenschappelijk Onderzoek–Vlaanderen Grant 12E3714N (to A. Meulders), and European Pain Federation-EFIC Grünenthal Research Grant E-G-G 169518451 (to A. Meulders).
Open Practices
All data have been made publicly available via the Harvard Dataverse Network and can be accessed at http://thedata.harvard.edu/dvn/dv/PainData. The complete Open Practices Disclosure for this article can be found at http://pss.sagepub.com/content/by/supplemental-data. This article has received the badge for Open Data. More information about the Open Practices badges can be found at https://osf.io/tvyxz/wiki/view/ and ![]() .
.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.