
Abstract
We used a visual illusion called motion repulsion as a model system for investigating competition between two mental representations. Subjects were asked to remember two random-dot-motion displays presented in sequence and then to report the motion directions for each. Remembered motion directions were shifted away from the actual motion directions, an effect similar to the motion repulsion observed during perception. More important, the item retrieved second showed greater repulsion than the item retrieved first. This suggests that earlier retrieval exerted greater inhibition on the other item being held in short-term memory. This retrieval-induced motion repulsion could be explained neither by reduced cognitive resources for maintaining short-term memory nor by continued inhibition between short-term memory representations. These results indicate that retrieval of memory representations inhibits other representations in short-term memory. We discuss mechanisms of retrieval-induced inhibition and their implications for the structure of memory.
Competition between representations is inevitable because of the limited capacity of human cognition. In perception, the outcomes of such competition are so compelling that people often fail to process task-relevant information when presented with multiple distractors (Cavanagh & Alvarez, 2005). Sometimes, even visually salient objects are rendered invisible (Kim & Blake, 2005). Representations also compete in memory. For example, proactive interference occurs when memory representations that were relevant in previous trials modify representations that are relevant in the present (Brown, 1958; Huang & Sekuler, 2010b; Peterson & Peterson, 1959). Memory retrieval also influences other items in short-term or long-term memory (Roediger, 1974; Tulving & Arbuckle, 1966), sometimes leading to memory loss (Anderson, 2003; Anderson, Bjork, & Bjork, 1994; Anderson & Spellman, 1995; Kuhl, Dudukovic, Kahn, & Wagner, 2007).
The literature regarding retrieval-induced forgetting in long-term memory suggests that items held in memory can be modified during retrieval as a result of inhibitory mechanisms (Anderson, 2003; Levy & Anderson, 2002). In the current study, we examined how retrieved representations from short-term memory interact with representations still held in short-term memory (Kuhl, Rissman, Chun, & Wagner, 2011; Levy & Anderson, 2002). This question is important for a number of reasons. First, memory retrieval must be considered when theories of short-term or working memory are being constructed, because executive control in working memory 1 needs to access and retrieve mental representations for further manipulation (Baddeley, 1992). Alternatively, short-term memories in different states are distinguished in relation to attention, and a retrieved representation is thought to be within the focus of attention (Cowan, 1988; Jonides et al., 2008; Larocque, Lewis-Peacock, & Postle, 2014; McElree, 1996; Oberauer, 2002). Second, researchers rely heavily on memory retrieval for identifying representations in short-term memory. However, if the memory retrieval itself modifies other representations still in short-term memory, measurements may be compromised when multiple retrievals are needed. Finally, the literature on retrieval-induced forgetting indicates that memory retrieval induces inhibition of competing representations (Anderson, 2003; Kuhl et al., 2007; Levy & Anderson, 2002), but it remains unknown whether inhibition is a common feature in the retrieval of both short-term and long-term memories.
In the present study, we used a visual illusion called motion repulsion as a model system to test whether retrieved representation from short-term memory inhibited competing representations held in short-term memory. We examined repulsive interactions that are typically found for motion direction. When two sets of random dots moving in different directions are superimposed, perceived differences in their motion directions are exaggerated compared with the actual motion differences, as if there were repulsion between the two motion directions (Marshak & Sekuler, 1979). Motion repulsion is an ideal model for studying competition-induced inhibition because it is thought to occur because of mutual inhibitions of the two pools of neurons representing the different motion directions (Hiris & Blake, 1996; Mather & Moulden, 1980). In addition, the repulsion is stronger when the two motion directions are approximately 45° apart. Using this characteristic feature, we can make inferences on how mental representations interact with one another.
Experiment 1
We examined whether memory retrieval of motion direction induces repulsion of the motion direction in a competing memory representation. Figure 1a illustrates the experimental procedure we used. Subjects were asked to remember two sets of random-dot-motion stimuli presented in sequence; one of the stimuli used red dots and the other used green dots. They were then asked to perform a well-established recall task in which they reported remembered motion directions by adjusting a clock hand. The color of the clock hand (i.e., red or green) matched the color of the motion stimulus for which the subjects were to recall the motion direction (Blake, Cepeda, & Hiris, 1997; Zokaei, Gorgoraptis, Bahrami, Bays, & Husain, 2011). In forward-recall trials, subjects were asked to report the motion directions of the first and second stimuli, in that order. In backward-recall trials, that order was reversed. The forward- and backward-recall trials were given in random order, and thus the subjects were unable to predict the recall order while encoding the stimuli.
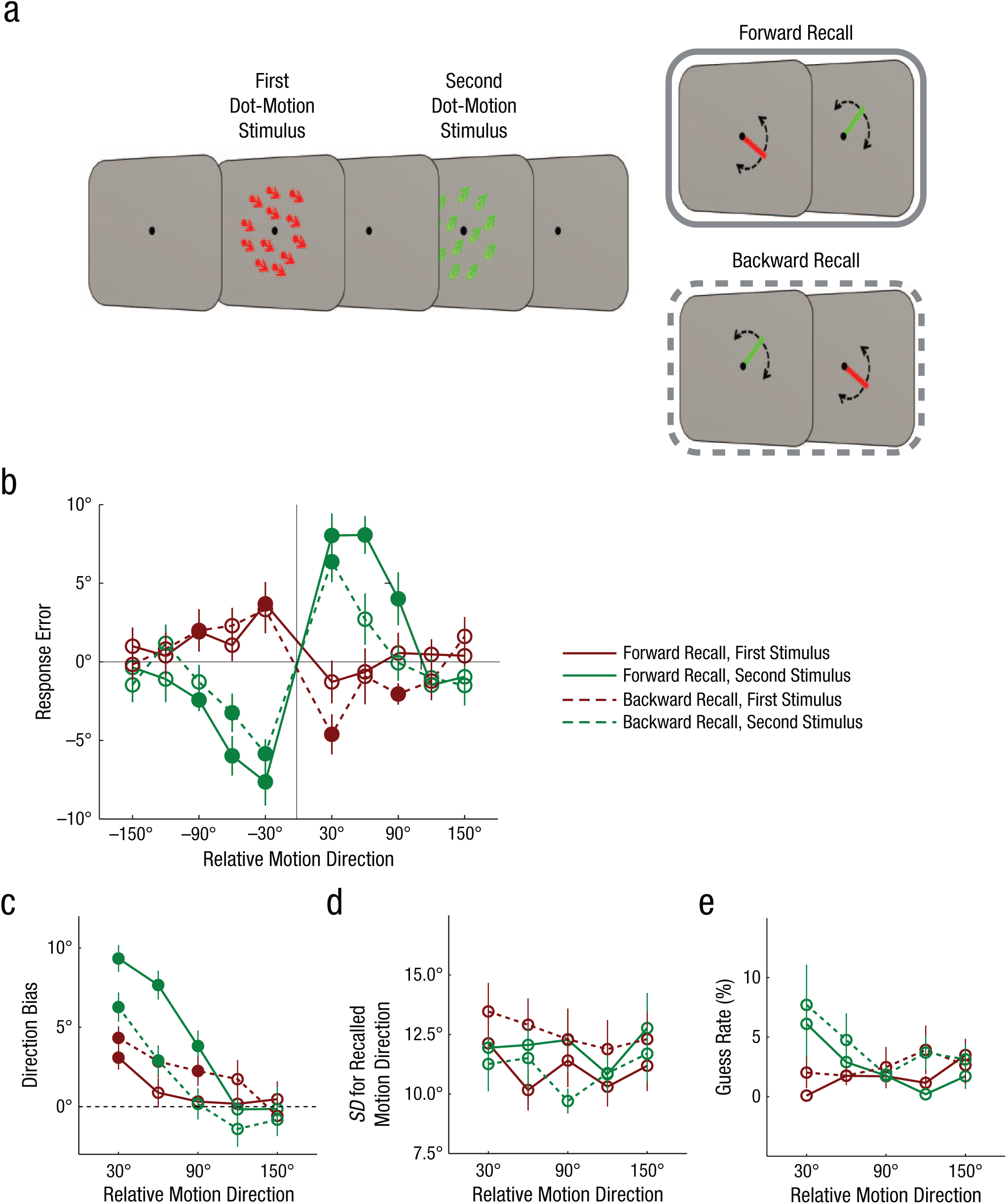
Procedure and results for Experiment 1. On each trial, two random-dot-motion stimuli were presented in sequence (a). Subjects then performed either a forward-recall task (reporting the motion direction of the first dot-motion stimulus and then the second) or a backward-recall task (reporting the motion direction of the second dot-motion stimulus and then the first). In the recall task, the color of the clock hand shown indicated the color of the motion stimulus to be recalled. Subjects moved the clock hand to indicate the motion direction of the stimuli. The graphs show (b) average response error for recalled direction, (c) direction bias, (d) standard deviation (SD) for recalled motion direction, and (e) guess rate as a function of the relative motion direction of the first and second dot-motion stimuli. Error bars represent ±1 SE. In (b) and (c), the filled circles indicate data points that are significantly different from 0. In (c), positive values indicate repulsive bias from the item in memory. In (c), (d), and (e), the results are collapsed across clockwise and counterclockwise relative motion directions (e.g., data from 30° clockwise and 30° counterclockwise).
These forward- and backward-recall trials served the purpose of the experiment as follows. Subjects had to hold the second motion stimulus in memory while retrieving the first motion stimulus. If retrieval of the first stimulus inhibits the competing representation (i.e., of the second stimulus), the repulsion magnitude for the second stimulus should be greater when it is retrieved second than when it is retrieved first. The opposite would be true for the backward-recall trials, in which the first stimulus was retrieved later than the second stimulus: The repulsion magnitude of the first stimulus should be greater than when it was retrieved before the second stimulus.
Method
We chose approximately 12 subjects on the basis of a previous study investigating short-term memory of motion stimuli. Specifically, Zokaei et al. (2011) obtained data for 11 subjects in their Experiment 1, but we added 1 more subject to counterbalance the stimulus color across subjects. We collected data for 13 subjects (including those of author J. Choi) but included data for only 12 subjects (5 men and 8 women; mean age = 26.6 years, age range = 22–27 years) in the analysis. Data for 1 subject were excluded because there were too many inaccurate trials (> 30%), as indicated by the feedback (described later in this section). All subjects provided informed consent, and the study was approved by the Sungkyunkwan University institutional review board. All subjects had normal or corrected-to-normal visual acuity.
Stimuli were presented on a CRT monitor (resolution = 1024 × 768 pixels; size = 31 × 24 cm; refresh rate = 85 Hz; mean luminance = 70.69 cd/m2; Samsung, Seoul, South Korea) placed 70 cm in front of the observers in a dimly illuminated room. We used Psychophysics Toolbox (Brainard, 1997; Pelli, 1997) running on a Mac Mini (Apple, Cupertino, CA) for all stimulus presentations and to control the experiment.
The fixation point was a dot surrounded by a circle subtending 0.3° of visual angle, presented in the center of the screen. The random-dot-motion stimulus consisted of approximately 78 dots within a circular window 5° in diameter. Each dot (0.05° in diameter) was replotted at a random location every 100 ms, and all dots moved coherently at 3.5° per second. The dots were either red (x = 0.63, y = 0.33, Y = 15.7 cd/m2) or green (x = 0.29, y = 0.60, Y = 34.9 cd/m2) on a gray background (48 cd/m2; the values given in parentheses are International Commission on Illumination, or CIE, color-space coordinates).
The trial sequence consisted of two motion stimuli and two recall tasks (Fig. 1a). Subjects were required to remember the direction in which the dots were moving for each stimulus. Each trial began with a 600-ms fixation period followed by the 500-ms presentation of the first motion stimulus (red). After a 1-s interval during which only the fixation point was visible, the second motion stimulus (green) was presented for 500 ms. The relative motion direction of the first and second stimuli varied from −150° to 150°, in steps of 30° (omitting 0°, which would constitute no change in motion direction). The colors of the motion stimuli were counterbalanced across sessions and observers.
The first recall task began 1 s after the offset of the second stimulus. To report the remembered motion direction, subjects adjusted a clock hand (2.5° in length from the fixation point) by moving a mouse. The clock hand was red for forward-recall trials and green for backward-recall trials. The initial direction of the clock hand was randomly chosen for each recall task. Once the subjects finished the adjustment, they clicked the mouse button to enter the response. Then, 1 s later, they performed the recall task for the other stimulus. If a response indicated that the remembered motion direction differed by more than 30° from the actual direction, the actual motion direction was presented for 500 ms as feedback. Each condition consisted of 1 of 10 relative motion directions and 1 of 2 recall orders (forward and backward), for a total of 20 conditions. Twenty trials of each condition were administered; thus, there were 400 trials in total. This number of trials was determined on the basis of a previous study in which 22 trials were used for each condition (Kang, Hong, Blake, & Woodman, 2011).
Response errors were measured as the difference between the reported motion direction and the true motion direction. We applied a standard mixture-model analysis with bias parameter. The model posits that there are two types of response errors: (a) response errors associated with memory representation, which are normally distributed near the true motion direction, and (b) response errors associated guessing, which are uniformly distributed over all directions (Zhang & Luck, 2008). We estimated three parameters—direction bias, standard deviation of recalled motion direction (as a proxy for memory precision), and guess rate—by using the MemToolbox (Suchow, Brady, Fougnie, & Alvarez, 2013).
Before the experiment, all subjects completed one practice session of a cued-recall task in which they were required to remember two random-dot-motion stimuli presented in sequence and to recall the direction of one of the motion stimuli, cued by the color of the clock hand.
Results
We found that memory retrieval of an item induced greater motion repulsion of the other item held in short-term memory. Figure 1b shows the remembered directions of the first (red) and second (green) motion stimuli as a function of relative motion direction. We created this figure by averaging differences between subjects’ responses and the true motion direction; we excluded trials in which the difference was more than 30° in either direction because the subjects had been provided with feedback for those responses during the experiment.
Motion repulsion occurred between the two stimuli such that the remembered motion directions of the first stimulus (red) and the second stimulus (green) were shifted away from their true directions, especially when the relative motion directions were approximately 45° apart. The characteristics of such repulsion mirror those found in a previous study, in which the repulsive interaction between the short-term memory representation and the perceptual representation was greater for the second stimulus than for the first one (Kang et al., 2011). More important, the repulsion magnitude was greater for the second stimulus in the forward-recall trials (i.e., the green solid line shows a greater shift than the green dashed line; Fig. 1b). In contrast, in the backward-recall trials, the repulsion was greater for the first stimulus (i.e., the red dashed line shows a greater shift than the red solid line; Fig. 1b).
For systematic analyses, we performed a mixture-model analysis that provided the following three measures: direction bias, precision of the memory representation, and subjects’ guess rate (Zhang & Luck, 2008). Specifically, the mixture model consists of two distributions with unequal weights; the remembered motion direction and its precision are represented by a normal distribution, and the subjects’ guess is represented by a uniform distribution. This approach is suitable especially because the direction bias and precision measures can be estimated without placing arbitrary criteria on subjects’ guesses (e.g., response errors larger than 30° in either direction as in Fig. 1b).
We first collapsed the responses for clockwise and counterclockwise relative motion directions (e.g., data from the 30° and −30° conditions were placed into the same model) to increase the number of trials. To accomplish this, we assigned positive values to errors shifted away from the item in the memory and negative values to errors shifted toward the item in the memory. A standard mixture model with bias parameter was then applied using MemToolbox (Suchow et al., 2013).
Figure 1c shows the magnitude of direction bias in memory representations. A three-way analysis of variance (ANOVA) with factors of relative motion direction, recall order (forward vs. backward), and stimulus order (first vs. second) yielded significant effects demonstrating characteristic motion repulsion between the two motion stimuli: The main effect of relative motion direction was significant, F(1, 11) = 38.65, p < .001, as was the Relative Motion Direction × Stimulus Order interaction, F(1, 11) = 16.47, p = .002. More important, recall order interacted significantly with stimulus order, F(1, 11) = 21.18, p < .001, which indicates that greater motion repulsion, particularly for relative motion directions close to 45° apart, was induced by memory retrieval compared with no memory retrieval. This result is consistent with the hypothesis that memory retrieval affects representations held in short-term memory.
The results for standard deviation of recalled motion direction, a measure of memory precision, also indicated that the retrieved memory representation interacted with the representation being held in short-term memory (Fig. 1d). A three-way ANOVA with factors of relative motion direction, recall order, and stimulus order yielded a significant interaction between retrieval order and stimulus order, F(1, 11) = 8.782, p = .013; variability in the reported motion direction of the same motion stimulus was higher when it was retrieved later. Other factors and interactions did not have significant effects, F(1, 11)s < 0.776, ps > .397. In contrast, guess rates were low in all conditions (M = 2.74%, SD = 3.5), and no factor was significant, F(1, 11) < 3.882, p > .076. These results are similar to those for repulsion magnitude (Fig. 1c): They indicate that the quality of memory representation was worse when retrieved later than when retrieved earlier, but the guess rate was not a sensitive measure to reveal retrieval-induced inhibition because memory demand was low (Fig. 1e).
We addressed two concerns regarding these results. First, we ruled out a concern that reporting the nontarget stimulus (e.g., reporting the motion direction of the second stimulus when given the cue to report the motion direction of the first stimulus) can induce direction bias (Bays, Catalao, & Husain, 2009). This concern was ruled out because the guess rate was too low to produce meaningful direction bias in the remembered motion directions, and, more important, if subjects had swapped the responses for the two motion stimuli, the direction bias would have been the opposite of what we found. Second, although the results for direction bias and standard deviation of recalled motion direction showed similar patterns, it is difficult to conclude that these two measures reflect the same aspects of memory representation. The next two experiments indicate that direction bias and standard deviation of recalled motion direction reflect different aspects of memory representation.
Experiment 2
The results of Experiment 1 are inconclusive regarding whether retrieval-induced motion repulsion was caused by reduced cognitive resources during retrieval or by the retrieved representation of the motion stimulus. The results can be explained by reduced cognitive resources (e.g., attention) for maintaining short-term memory during memory retrieval. For example, when subjects perform the recall task, they have to process cue information to determine which stimulus to retrieve and adjust the clock hand for the remembered motion direction. Although the task is simple, cognitive resources have to be directed away from maintaining items in memory during memory retrieval. Thus, the items in memory can be more susceptible to further memory distortion regardless of retrieved representation.
This hypothesis is reasonable considering the results for standard deviation of recalled motion in Experiment 1. Previous studies have shown that quality of memory representation is compromised when a subject’s attention is diverted to other items (Huang & Sekuler, 2010a; Williams, Pouget, Boucher, & Woodman, 2013). We found consistently that standard deviation of recalled motion direction for the same motion stimulus was higher when that stimulus was retrieved later (Fig. 1d). Taken together, these results indicate that if the reduced cognitive resources are responsible for the retrieval-induced repulsion, the standard deviation of recalled motion direction should be positively correlated with the direction bias.
To test this alternative hypothesis, we asked subjects to perform a memory task in which they were required to remember three stimuli in sequence: the directions of the two motion stimuli and the color of an object. They were then asked to perform two recall tasks concerning those three items (Fig. 2a). In particular, we created a memory-matching task for a colored square. In this task, six different colored squares, including one in the target color, were randomly presented over an imaginary circle while subjects rotated a white clock hand to point at the square that matched the remembered color. We designed this memory-matching task to recruit cognitive processes similar to those recruited by the memory-recall task for the motion stimulus (e.g., cue processing and rotating the clock hand) while remaining sufficiently easy for subjects to remember all three items.

Procedure and results for Experiment 2. On each trial, a colored square and two random-dot-motion stimuli were presented in sequence (a). Subjects then performed a recall task in which they reported the color of the square and the direction of motion of the first dot-motion stimulus, the color of the square and the direction of motion of the second dot-motion stimulus, or (as in the Experiment 1 forward-recall task) the directions of motion of the first and second dot-motion stimuli. To report the color of the square, subjects moved a white clock hand. In the motion-stimuli-recall tasks, the color of the clock hand shown indicated the color of the motion stimulus to be recalled. Subjects moved the clock hand to indicate the motion direction of the stimulus. The graphs show (b) direction bias, (c) standard deviation (SD) of recalled motion direction, and (d) guess rate as a function of the relative motion directions of the second dot-motion stimulus in two different conditions (colored square + second motion stimulus, and first motion stimulus + second motion stimulus). Error bars represent ±1 SE. In (b), the filled circles indicate data points that are significantly different from 0, and positive values indicate repulsive bias from the item in memory. In (b), (c), and (d), results are collapsed across clockwise and counterclockwise relative motion directions (e.g., data from 30° clockwise and 30° counterclockwise).
We hypothesized that if cognitive processes involved in memory retrieval produced greater repulsion because of decreased cognitive resources for memory maintenance in Experiment 1, the magnitude of motion repulsion of the second motion stimulus should be comparable whether it is retrieved after a colored square or after the first motion stimulus. However, if greater motion repulsion is induced by inhibition of the retrieved motion stimulus, the magnitude of direction bias of the second motion stimulus should be greater only when it is retrieved after the retrieval of the first motion stimulus. We also investigated whether the magnitude of motion repulsion was correlated with the variability of memory representation, as mentioned earlier, which provided an opportunity for testing the hypothesis, considering that the amount of cognitive resources cannot be equated between the memory-matching and motion-recall tasks.
Method
Fourteen subjects (3 men and 11 women; mean age = 23.9 years, age range = 18–27 years) provided informed consent, and the study was approved by the Sungkyunkwan University institutional review board. All observers had normal or corrected-to-normal visual acuity.
Subjects were asked to perform a memory task in which a colored square and two random-dot-motion stimuli were presented in sequence. They were then asked to complete two recall tasks. The square was one of six colors (red, green, blue, yellow, magenta, and cyan). In the memory-matching task, squares of all six colors were presented over an imaginary circle in random order, and subjects were required to rotate a white clock hand to point at the square that matched the remembered color (Fig. 2a). We considered responses to be incorrect if the direction of the clock hand differed from the center of the probe item by more than 30° in either direction, because each colored square was 60° apart from the two closest squares. The motion stimuli and the motion-direction-recall task were similar to those used in the forward-recall trials in Experiment 1.
Subjects were required to report two items among the three stimuli. On one third of the trials, subjects were asked to report the color of the square and then the direction of the first motion stimulus (which was not of interest in this experiment but was included so that subjects would have an equal probability of recalling each stimulus). On another third of the trials, subjects were asked to report the color of the square and the direction of the second motion stimulus. In the remaining third of the trials, subjects were asked to report the motion directions of the first and second motion stimuli. The three trial types were presented in pseudorandom order.
Results
The colored square was correctly recalled in 96.1% (SD = 2.4) of trials. This indicates that cognitive resources were directed toward retrieving the colored square while holding the directions of the other two motion stimuli in short-term memory.
For the memory representation of the second motion stimulus, we obtained direction bias, standard deviation of recalled motion direction, and guess rate from the mixture-model analysis. Figure 2b shows direction bias (a measure of repulsion magnitude) when the motion direction of the second motion stimulus was reported after memory retrieval of the colored square and when it was reported after memory retrieval of the first motion stimulus. The responses for clockwise and counterclockwise relative motion directions (e.g., data from 30° and −30°) were collapsed (as in Fig. 1c). Motion repulsion was robustly reproduced. The repulsion magnitude of the second stimulus was greater when it was retrieved after the direction of the first motion stimulus than when it was retrieved after the colored square. A two-way ANOVA with factors of relative motion direction and prior retrieved item (colored square vs. first motion stimulus) yielded significant effects: The main effect of relative motion direction was significant, F(1, 13) = 31.63, p < .001, as was the main effect of prior retrieved item, F(1, 13) = 19.86, p < .001. The Relative Motion Direction × Prior Retrieved Item interaction was significant as well, F(1, 13) = 10.14, p = .007. This result cannot be explained by reduced cognitive resources for maintaining short-term memory representations.
The results for standard deviation of recalled motion direction and guess rate showed that memory retrieval of the colored square did affect the memory representation of the direction of the second motion stimulus, which was disrupted more when it was retrieved after the colored square than after the direction of the first motion stimulus. For standard deviation of recalled motion direction, a two-way ANOVA with factors of relative motion direction and prior retrieved item (colored square vs. first motion stimulus) yielded significant effects: The main effect of relative motion direction was significant, F(1, 13) = 8.129, p = .013, as was the main effect of prior retrieved item, F(1, 13) = 14.62, p = .002. The Relative Motion Direction × Prior Retrieved Item interaction was significant as well, F(1, 13) = 6.81, p = .021 (Fig. 2c). For guess rate, a two-way ANOVA using the same factors yielded a significant main effect of prior retrieved item, F(1, 13) = 19.00, p < .001 (Fig. 2d).
Several accounts can be offered for the higher standard deviation of recalled motion direction and guess rate of the second motion stimulus after retrieval of the colored square than after the retrieval of the first motion stimulus. The memory-matching task could be more disruptive than the motion-recall task because subjects had to process six items and the clock hand in the memory-matching task, whereas they had to process only the clock hand in recalling motion direction (Magnussen & Greenlee, 1999). Retrieving memory representations of the two different categories might be more demanding than retrieving representations of the same category, as shown by results when subjects switch attentional sets (Visser, Bischof, & Di Lollo, 1999), or perceptual differences between color and motion stimuli might result in nonspecific inhibition of direction, as found when similarity between memory items is manipulated in retrieval-induced forgetting tasks (Anderson, Green, & McCulloch, 2000). Distinguishing among these alternatives is beyond the scope of the present study. However, these results provide even stronger evidence that the retrieved motion stimulus produced greater motion repulsion, rather than reduced cognitive resources for maintaining short-term memory. This is because the hypothesized cognitive resources should result in similar patterns of results for direction bias and standard deviation of recalled motion direction, but we found that they had opposite patterns of results.
Experiment 3
It is still possible that what appears to be retrieval-induced motion repulsion actually reflects continued motion repulsion within short-term memory, which could have occurred regardless of memory retrieval. Specifically, in the forward-recall trials of Experiment 1, the second stimulus should be held in short-term memory for a greater period of time than the first stimulus while subjects were performing the recall task. The opposite is true for the backward-recall trials; thus, the retention interval may be a determining factor for the retrieval-induced motion repulsion. This hypothesis is reasonable because the perceptual repulsion of the superimposed motion stimuli increases with the duration of presentation (Rauber & Treue, 1999). We tested this hypothesis by asking subjects to perform only a single recall task for one of the two motion stimuli while using various retention intervals (0.5–6 s) before the recall task.
Method
Fourteen subjects (4 men and 10 women; mean age = 24.8 years, age range = 18–24 years), including author J. Choi, provided informed consent, and the study was approved by the Sungkyunkwan University institutional review board. All subjects had normal or corrected-to-normal visual acuity.
Figure 3a illustrates the stimulus sequence used in Experiment 3, which was very similar to Experiment 1 with the following exceptions. First, subjects were asked to remember the directions of two random-dot-motion stimuli and then to perform a memory-recall task for only one of the two stimuli, cued by the color of the clock hand on the response screen. The first and the second motion stimuli were probed on equal numbers of trials. Second, we used various retention intervals (0.5, 2, 4, and 6 s). The retention interval was measured from the offset of the second stimulus to the onset of the memory cue (i.e., the clock hand). Third, the relative motion directions were varied at ±20°, ±40°, ±60°, ±100°, and ±150° to characterize in detail the motion repulsion near 45° in either direction. Finally, subjects repeated the task during four 50-min sessions, once per day for 4 days. This schedule resulted in 12 trials for each condition, for a total of 480 trials.

Procedure and results for Experiment 3. On each trial, two random-dot-motion stimuli were presented in sequence (a). After a variable retention interval, subjects were asked to recall one of the two motion directions. The color of the clock hand shown indicated the color of the motion stimulus to be recalled. Subjects moved the clock hand to indicate the motion direction of the stimulus. The graphs show (b) direction bias, (c) standard deviation (SD) of recalled motion direction, and (d) guess rate as a function of the relative motion directions of the first (red) and second (green) dot-motion stimuli for each of the four retention intervals. Error bars represent ±1 SE. In (b), the filled circles indicate data points that are significantly different from 0 and positive values indicate repulsive bias from the item in memory. In (b), (c), and (d), results are collapsed across clockwise and counterclockwise relative motion directions (e.g., data from 30° clockwise and 30° counterclockwise).
However, we were concerned with statistical power, so we repeated Experiment 1 before performing Experiment 3 to determine the strength of the retrieval-induced repulsion. We did this because the number of trials in Experiment 3 was reduced by 60% compared with Experiments 1 and 2 (20 trials per condition in each), but if we performed 20 trials for each condition, the experiment would have required 3 more days to complete for each subject, which would have been impractical. The replication result indicated that the protocol we used with 12 trials for each condition did not undermine statistical power and enabled us to manipulate the retention interval up to 6 s if at least 14 subjects were recruited. The results of the replication of Experiment 1 are described in the Supplemental Material available online.
Results
Figure 3b shows the magnitude of direction bias (red for the first stimulus and green for the second stimulus) as a function of retention interval for each relative motion direction. As in Experiment 1, motion repulsion occurred between the first motion stimulus (red) and the second motion stimulus (green). The repulsion magnitude was greater for the second stimulus than for the first. However, repulsion magnitude was unaffected by retention interval. A three-way ANOVA with factors of relative motion direction, retention interval, and stimulus order yielded significant effects confirming the defining characteristics of motion repulsion between the first and the second motion stimuli: The main effect of relative motion direction was significant, F(1, 13) = 28.30, p < .001, as was stimulus order, F(1, 13) = 17.41, p = .001, and the Relative Motion Direction × Stimulus Order interaction, F(1, 13) = 12.68, p = .003. However, direction bias did not change with retention interval at all, such that the main effect of retention interval was not significant, F(1, 13) = 0.968, p = .343, and retention interval did not interact significantly with any other factors, F(1, 13) < 0.902, p > .36.
In contrast to direction bias, the standard deviation of recalled motion direction increased with retention interval (Fig. 3c); that is, regardless of relative motion directions, longer retention intervals were associated with greater disruption of the quality of memory representations. A three-way ANOVA with factors of relative motion direction, retention interval, and stimulus order yielded significant main effects of retention interval, F(1, 13) = 31.67, p < .001, and relative motion direction, F(1, 13) = 15.39, p = .002. Stimulus order and interactions of the factors did not have significant effects, F(1, 13) < 1.75, p > .209. The general increase in standard deviation of recalled motion direction in memory representation as a function of retention interval is consistent with results from previous studies showing degradation of short-term memory representation over time (Fougnie & Alvarez, 2011; Magnussen & Greenlee, 1999; but see Zhang & Luck, 2009).
Guess rates (Fig. 3d) were fairly low for all conditions (M = 2.17%, SD = 1.6), but retention interval had a significant effect on guess rate, F(1, 13) = 5.199, p = .040, and the interaction of relative motion direction and stimulus order had a significant effect on guess rate, F(1, 13) = 5.232, p = .039. However, it is difficult to conclude that changes in the retention interval produced systematic changes in the guess rate, because pairwise t tests with Holm’s adjustment resulted in significant differences only for guess rates for retention intervals between 0.5 s and 4 s (p = .019).
Taken together, the findings indicate that retrieval-induced inhibition remains a viable account for the direction-bias result in Experiment 1. On the other hand, continued interaction between two short-term memory representations can explain the result for standard deviation of recalled motion direction in Experiment 1 (Fig. 1d) such that a longer retention interval during the first memory retrieval is responsible for the decreasing quality of memory representation being held in short-term memory.
Discussion
We have shown that memory retrieval of a motion stimulus produces greater motion repulsion of another item held in visual short-term memory. The retrieval-induced motion repulsion cannot be explained by either reduced cognitive resources for maintaining short-term memory or by continued interactions between short-term memory representations. Considering that mutual inhibition in motion direction produces characteristics of motion repulsion as a function of relative motion direction, our results indicate that memory retrieval of a motion stimulus from short-term memory inhibits competing representations still held in short-term memory.
How does then the memory retrieval of motion stimulus induce greater motion repulsion? The literature on retrieval-induced forgetting posits that executive control inhibits competing representations while retrieving items from long-term memory (Anderson, 2003; Levy & Anderson, 2002), similar to inhibiting competing responses (Logan, 1994). The proposed mechanism for retrieval-induced forgetting also provides a parsimonious account for the retrieval-induced motion repulsion: Executive control accesses memory presentation in short-term memory (Baddeley, 1992), and mutual inhibition between two motion stimuli is responsible for motion repulsion (Hiris & Blake, 1996; Mather & Moulden, 1980).
Our paradigm is distinguished from that of previous studies in that it provides evidence for retrieval-induced inhibition based on direction-specific motion repulsion. Until now, the retrieval-practice paradigm has been used to demonstrate that memory retrieval can lead to memory loss via inhibition in long-term memory (Anderson et al., 1994). Subjects study lists of category-exemplar pairs and then perform retrieval practice on half of the exemplars from a subset of categories. When subjects are tested on all of the category-exemplar pairs, their performance for the practiced exemplars is superior to that for the unpracticed exemplars. However, subjects forget more unpracticed exemplars from the practiced categories than they forget unpracticed exemplars from the unpracticed categories. This failure in memory for the unpracticed items is considered evidence for retrieval-induced inhibition, such that both the practiced and unpracticed exemplars compete for the same category cue during the retrieval practice, and the executive control process inhibits competing unpracticed representations when retrieving practiced exemplars (Anderson, 2003; Levy & Anderson, 2002). However, we demonstrated retrieval-induced inhibition with only two items in short-term memory by adopting motion repulsion, which is thought to occur because of mutual inhibition of competing motion stimuli.
In addition, the present study contributes to the growing literature showing similarities between short- and long-term memories (Jonides et al., 2008; Ranganath & Blumenfeld, 2005). Mechanisms of memory disruption in short-term memory have been discussed mainly in the contexts of decay over time and interference between representations (for review, see Jonides et al., 2008). The present study expands the scope of memory disruption in short-term memory by including retrieval-induced inhibition. This suggests that inhibition is a common feature of memory retrieval for both short- and long-term memory, a notion that is consistent with findings from a recent neuroimaging study showing that similar neural processes are involved in short- and long-term memory retrieval (Nee & Jonides, 2008).
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2013S1A5A8025812).
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.