
Abstract
Associative activation is commonly assumed to rely on associative strength, such that if A is strongly associated with B, B is activated whenever A is activated. We challenged this assumption by examining whether the activation of associations is state dependent. In three experiments, subjects performed a free-association task while the level of a simultaneous load was manipulated in various ways. In all three experiments subjects in the low-load conditions provided significantly more diverse and original associations compared with subjects in the high-load conditions, who exhibited high consensus. In an additional experiment, we found increased semantic priming of immediate associations under high load and of remote associations under low load. Taken together, these findings imply that activation of associations is an exploratory process by default, but is narrowed to exploiting the more immediate associations under conditions of high load. We propose a potential mechanism for processing associations in exploration and in exploitation modes, and suggest clinical implications.
Keywords
Individuals’ state of mind and modes of behavior are proposed to be continuously affected by an interesting tension in the brain between exploration and exploitation. At the exploratory end of this continuum, people attend the environment with a wider scope, are more learning oriented, and are attuned more to sensory input than to familiar knowledge in memory (Hills et al., 2015). At the exploitatory end, they rely more on what they already know and on their expectations, are less open to novelty and surprises, and gravitate more to the details than to the “big picture” (Schwartenbeck, FitzGerald, Dolan, & Friston, 2013). Time, context, goals, and level of interest have an effect on where an individual is positioned on the exploration-exploitation spectrum (see Cohen, McClure, & Yu, 2007, for a review).
One interesting question is whether the tension between exploration and exploitation is maintained when individuals attend their inner mental space, rather than the physical world around them. If this is the case, then the availability of mental resources may be a critical factor that directly influences where an individual is on this continuum. When people are stressed, and when they are simply intensely focused, they become less likely to notice peripheral information (Booth & Sharma, 2009; Simons & Chabris, 1999). We hypothesized that limiting subjects’ mental resources would similarly bias how exploratory or exploitatory their mental processes would be. Therefore, in a series of studies, we examined the effect of load on associative activation in a free-association task. We used associative activation as a proxy for how exploratory subjects’ thinking was, assuming that when subjects were biased toward exploration, their associations would be more unique and broader in scope, whereas when they were in a more exploitatory state, their associations would be narrower and more consensual.
The level of consensus achieved in free-association settings has previously been used to model relatedness or associative strength (e.g., Nelson, McEvoy, & Dennis, 2000; Nelson, McEvoy, & Schreiber, 2004). In models of spreading activation, this factor of relatedness traditionally served as the key predictor for associative processing. According to such models, when an individual encounters A, B has the highest chances of being triggered if B has the strongest association with A (Anderson, 1983; Collins & Loftus, 1975; Neely, 1977).
The debate on whether associative activation is automatic or not is ongoing, and recent findings obtained using priming paradigms suggest that spreading activation of associations depends on executive resources (Heyman, Van Rensbergen, Storms, Hutchison, & De Deyne, 2015; Hutchison, Heap, Neely, & Thomas, 2014). In any case, however, activation of associations is considered to follow the central prediction of spreading-activation models, that the level of activation depends on the strength of association. The current study shows that this prediction is not entirely valid.
We asked subjects to report rapidly the first association that came to mind as they viewed each of a series of target words. Simultaneously, we manipulated subjects’ load. Our hypothesis was that if activation of associations is affected by mental mode (i.e., location on the exploration-exploitation continuum) and not determined solely by inherent associative strength, then subjects’ reported associations would differ between high- and low-load conditions. Specifically, we expected that in high-load conditions, which are presumably conducive to a more exploitatory state of mind, associations would rely primarily on associative strength, and would be significantly less diverse than in low-load conditions, which presumably afford a more exploratory state. All the studies reported here were approved by the University of Bar-Ilan Brain Research Ethics Committee.
Experiment 1a: The Effect of Working Memory Load on Associative Activation
In our first experiment, we examined our main hypothesis regarding the possible effect of cognitive load on associative activation. Cognitive load was manipulated via a working memory (WM) task, given that WM and cognitive load are often taken as equivalent (Sweller, 1988).
Method
Subjects
Twenty subjects (13 females; mean age = 22.75 years, SD = 2.55) participated in this study in return for course credits. 1
Tasks
In the free-association task, a word-association task (Zeelenberg, Pecher, Shiffrin, & Raaijmakers, 2003), subjects were asked to respond, as quickly as possible, to each of a series of target words with the first association that came to mind. They were assured that there were no correct answers and that they would not be asked about their associations later. One hundred target words were used as stimuli in this task. Fifty words were taken from a list developed by Jung (1910), who used the method of free association in clinical settings, and 50 words, controlled for frequency and imaginability, were taken (with some adaptations) from a study by Jefferies, Patterson, Jones, and Lambon Ralph (2009). Subjects typed their responses using a computer keyboard, and their reaction time (RT) was measured as the time from the onset of the target word to the first key press of the answer.
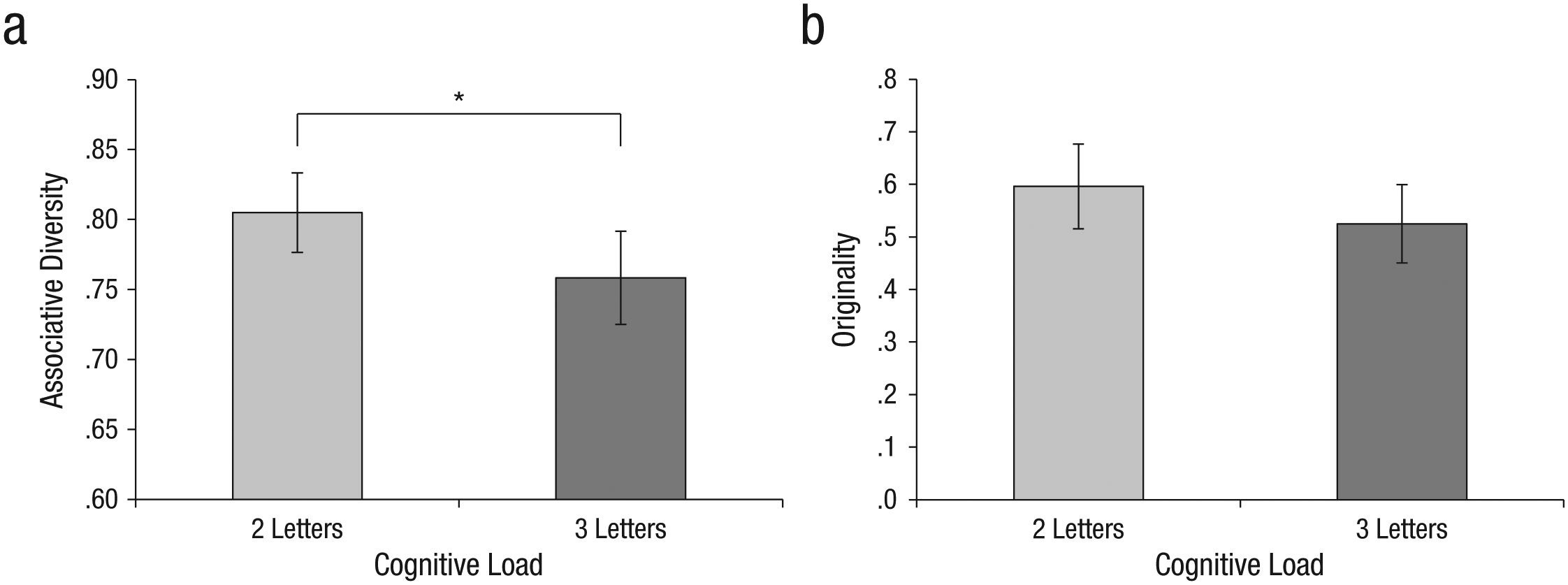
Our WM task was a commonly used adaptation of the digit-span task. Subjects were asked to hold in memory strings of digits, and were later requested to repeat them explicitly. This task has been shown to occupy phonological-repetition resources, a key component of WM (Baddeley, 2000). In the current experiment, subjects in the low-load condition were asked to remember short strings (two digits), and those in the high-load condition were asked to remember long strings (six digits). The digits were randomly chosen and were presented visually. A different string was presented for each block of free-association trials.
Procedure
After signing consent forms and receiving brief instructions, subjects were randomly assigned to an experimental condition. Each block of trials began with a 10-s presentation of the digit string to hold in memory. Subjects were then shown 10 target words, in random order, and were instructed to type the first word that came to mind for each, as quickly as possible. Time for each trial was not limited, and only after an answer was given did the following target word appear. At the end of each block, subjects repeated the digits held in memory. After 10 blocks, the experiment ended, and subjects were debriefed and thanked.
Results
Accuracy on the WM task differed significantly between the conditions. Subjects under low load were more accurate (M = .96, SD = .08) than subjects under high load (M = .78, SD = .23), t(11) = 2.28, p < .04, Cohen’s d = 1.37. This result confirmed that the cognitive-load manipulation was effective.
Analysis of RT in the free-association task (including only blocks for which the WM response was correct) showed no significant RT differences between the low-load condition (M = 3.16 s, SD = 1.2) and the high-load condition (M = 2.91 s, SD = 1.13), p > .25.
Because of the open-ended nature of the free-association task (i.e., numerous answers were possible for each target word), we tested for a difference between the conditions by analyzing the variance in answers across subjects. Our indices of variance were the proportion of trials on which nondominant answers were given and the proportion of trials on which the dominant answer was given. Independent-samples t tests revealed significant differences between conditions. The dominant answers for target words were repeated on a greater proportion of trials in the high-load condition (M = .34, SD = .14) than in the low-load condition (M = .27, SD = .1), t(175) = −3.64, p < 3 × 10−4, Cohen’s d = 0.55. In addition, nondominant answers were given less often under high load (M = .67, SD = .16) than under low load (M = .75, SD = .13), t(189) = 4.01, p < 8 × 10−5, Cohen’s d = 0.58. Thus, there was a significant reduction in variance of associations under high load.
To create a single measure of diversity, we calculated the overall associative diversity in each condition as the proportion of maximal entropy in associations:
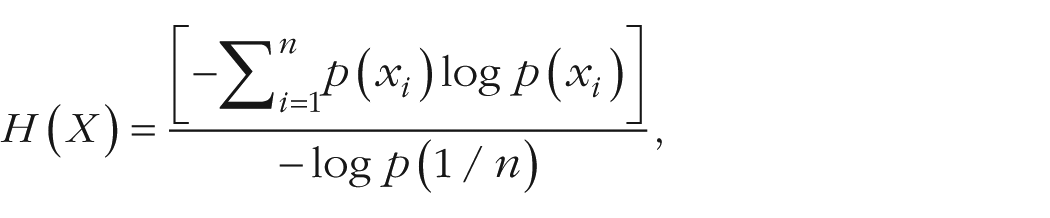
where x is the association given to a target word. For each condition, we calculated the entropy in the distribution of associations given to each target word and divided this score by the maximal entropy possible for the number of answers provided. This measure of entropy relies on the probability of each association being provided and is therefore sensitive both to the number of times an answer is provided and the overall number of different associations given.
The level of associative diversity was higher for answers given under low load (M = .82, SD = .11) compared with answers given under high load (M = .74, SD = .15), and this effect was significant, t(176) = 4.25, p < 3 × 10−6, Cohen’s d = 0.64 (Fig. 1a). This entropy difference in free associations implies that subjects were less “informative,” and more predictable, under high load than under low load.

The effect of working memory load on associative activation in Experiment 1a. The graphs show (a) the proportion of associative diversity (measured as maximal entropy) and (b) mean originality scores (unique responses) in the low-load and high-load conditions. Error bars indicate 95% confidence intervals, and asterisks indicate significant differences between conditions (*p < .05, ***p < .001).
Finally, to confirm that the effect of high load on associations was not caused by a few highly associative or highly uninterested subjects, we compared originality in responses between the conditions. Each subject’s score for originality was calculated as the proportion of unique answers given. A response was scored as 1 if it was the only such response given among all subjects and as 0 if it was repeated by at least 1 other subject, and the proportion of unique answers was calculated. This analysis showed that originality decreased as load increased. Overall, originality scores were higher in the low-load condition (M = .60, SD = .08) than in the high-load condition (M = .50, SD = .10), and this difference was significant, t(17) = 2.35, p < .031, Cohen’s d = 1.1 (Fig. 1b).
This first experiment showed that taxing cognitive resources (WM) directly reduces diversity in associative responses. To test whether this effect is reliable, we conducted a similar experiment, with different subjects, while slightly changing the load levels in the WM task.
Experiment 1b: Replicating the Effect of WM Load on Associative Activation
Method
Subjects
Twenty subjects (15 females; mean age = 22.05 years, SD = 2.25) took part in this experiment in exchange for course credits. Informed consent was obtained from all subjects.
Procedure
The experimental procedure was identical to that in Experiment 1a except that subjects in the low- and high-load conditions were instructed to remember strings of four and seven digits, respectively (instead of two and six digits), while completing the free-association task.
Results
As in Experiment 1a, subjects under low load were significantly more accurate in the WM task (M = .87, SD = .13) than subjects under high load (M = .66, SD = .17), t(16) = 2.98, p < .008, Cohen’s d = 1.79. Thus, the load manipulation was effective.
Analysis of RTs in the free-association task (including only blocks for which the WM response was correct) showed no significant difference between the conditions (low load: M = 2.93 s, SD = 0.76; high load: M = 2.35 s, SD = 0.54), p < .06.
Analysis of answers in the free-association task showed that the dominant answers were provided more frequently across subjects in the high-load condition (M = .33, SD = .14) than in the low-load condition (M = .28, SD = .13), t(196) = −2.84, p < .005, Cohen’s d = 0.4. In addition, nondominant associations were given less often under high load (M = .68, SD = .15) than under low load (M = .76, SD = .15), t(197) = 3.24, p < .001, Cohen’s d = 0.46. The level of diversity in associations, as measured by the proportion of maximal entropy, was significantly higher in the low-load condition (M = .82, SD = .14) than in the high-load condition (M = .76, SD = .14), t(197) = 2.97, p < .003, Cohen’s d = 0.42 (Fig. 2a).
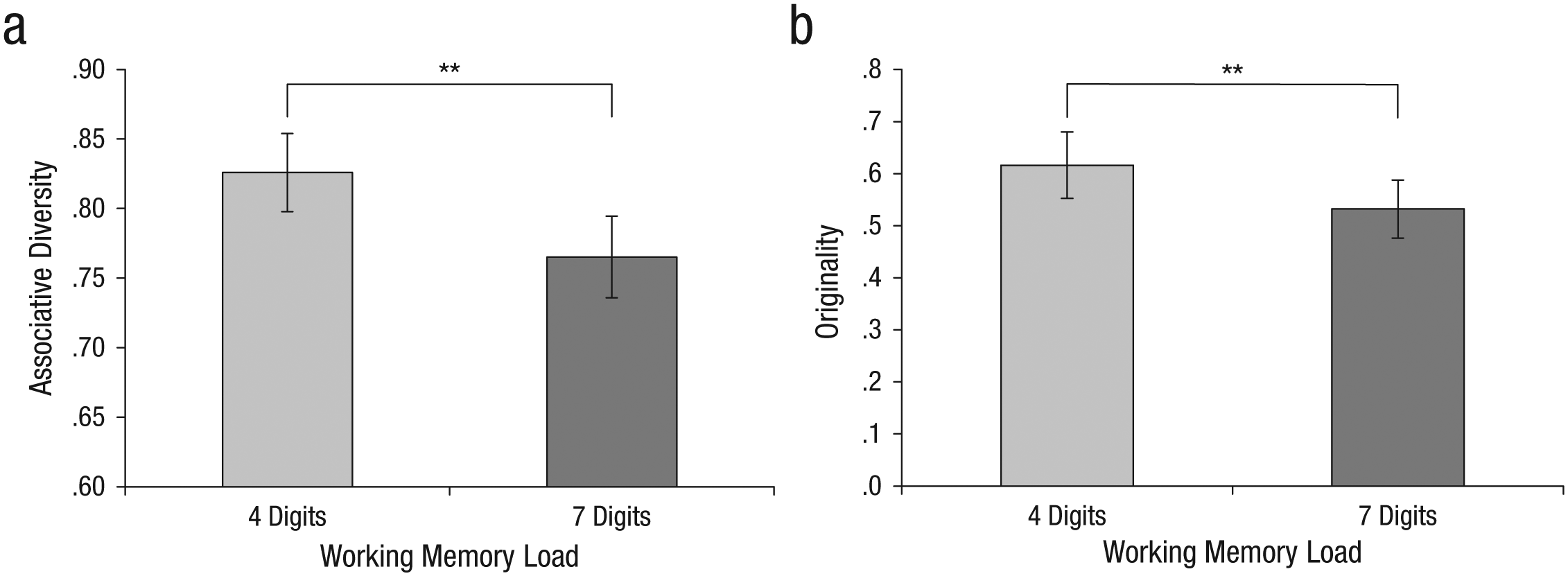
The effect of working memory load on associative activation in Experiment 1b. The graphs show (a) the proportion of associative diversity (measured by maximal entropy) and (b) mean originality scores (unique responses) in the low-load and high-load conditions. Error bars indicate 95% confidence intervals, and asterisks indicate significant differences between conditions (**p < .01).
The analysis of originality scores showed that subjects were significantly more original under low load (M = .61, SD = .08) than under high load (M = .53, SD = .07), t(17) = 2.24, p < .01, Cohen’s d = 1.08 (Fig. 2b).
Experiments 1a and 1b demonstrate how the availability of WM resources affects the breadth of associations, possibly reflecting the distinction between the exploration and exploitation modes. These results suggest a tendency toward exploration by default, and a tendency toward exploitation when resources are low. One question that can be raised is whether the effect of load on associative activation is caused specifically by WM load. Therefore, in the next experiment, we manipulated cognitive load without taxing WM resources.
Experiment 2: Non-WM Cognitive Load Also Decreases Diversity in Associative Activation
Method
Subjects
Twenty-one subjects (12 females; mean age = 24.35 years, SD = 3.88) took part in this experiment in exchange for either course credits or a monetary payment. Informed consent was obtained from all subjects.
Tasks
The free-association task was the same as in the previous experiments. In addition, subjects completed an alphabetization task, in which they were asked to indicate the correct alphabetical order of the first letters of each target word. We manipulated the difficulty level of this task. In the low-load condition, subjects were asked to press the keys “1” and then “2” if the first letter of the word preceded the second letter in the alphabet, and to press the keys “2” and then “1” if the first letter followed the second. In the high-load condition, subjects were asked to indicate the correct alphabetical order of the first three letters of the target word in the same way, using the “1,” “2,” and “3” keys (Fig. 3). Because the stimuli were the target words from the free-association task, they were identical in the two conditions; the only difference between the conditions was the added computational requirement in the high-load condition.

Illustration of the alphabetization task in Experiment 2. Subjects were asked to indicate the alphabetical order of the first letters of each target word. In the low-load condition, subjects indicated the order of the first two letters, using the “1” and “2” keys. For example, the correct response to the word white was to press “2” and then “1” (w comes after h in the alphabet). In the high-load condition, subjects indicated the order of the first three letters of the same target words, using the “1,” “2,” and “3” keys. Thus, the correct response to the word white in the high-load condition was to press “3,” then “1,” and then “2” (of the three letters w is third in the alphabet, h is first, and i is second).
Procedure
After the subjects were given the instructions for the experiment, they were randomly assigned to either the high- or the low-load experimental condition. As in the previous experiments, the target words were presented serially, in random order. On each trial, subjects were asked to provide an association and to indicate the alphabetical order of the word’s first two (low-load condition) or three (high-load condition) letters. The order in which the two tasks were to be performed changed randomly across trials, and each target word was presented until both tasks were completed. Given this task-switching setting, subjects could perform each task separately and not lack WM resources while they retrieved associations for the target words. As in the previous experiments, the procedure consisted of 10 blocks of 10 trials each. After completing the experiment, subjects were debriefed and thanked.
Results
Of the 21 students, 1 was excluded for being an outlier (RT on the free-association task > 2 SD above the mean). Therefore, in the analyses, each condition included 10 subjects.
Analysis of performance in the alphabetization task suggests that the manipulation was successful. Although the difference was not significant, subjects in the high-load condition were less accurate (M = .94, SD = .04) than those in the low-load condition (M = .97, SD = .01), p < .06. Nonetheless, subjects under high load were significantly slower (M = 3.5 s, SD = 0.57) than those under low load (M = 2.23 s, SD = 0.74), t(16) = −4.3, p < 4 × 10−4, Cohen’s d = 2.15.
As in the previous experiments, analysis of RT in the free-association task showed no significant differences between the high-load condition (M = 2.56 s, SD = 0.68) and the low-load condition (M = 2.15 s, SD = 0.29), p < .10.
The main analysis revealed differences between the conditions in the measures of associative diversity. The dominant associations were given on a greater proportion of trials in the high-load condition (M = .34, SD = .15) than in the low-load condition (M = .30, SD = .14), though this difference was not significant, t(196) = −1.62, p < .1. Nonetheless, the proportion of nondominant associations provided for each target word significantly decreased as load increased (high load: M = .68, SD = .01; low load: M = .73, SD = .01), t(196) = 2.26, p < .02, Cohen’s d = 0.32. The proportion of maximal entropy was significantly higher for answers given under low load (M = .80, SD = .14) compared with answers given under high load (M = .75, SD = .16), t(193) = 2.11, p < .03, Cohen’s d = 0.30 (Fig. 4a).
The effect of non–working memory cognitive load on associative activation in Experiment 2. The graphs show (a) the proportion of associative diversity (measured by maximal entropy) and (b) mean originality scores (unique responses) in the low-load and high-load conditions. Error bars indicate 95% confidence intervals, and the asterisk indicates a significant difference between conditions (*p < .5).
Analysis of the originality scores showed that subjects under low load were more original (M = .59, SD = .11) than subjects under high load (M = .52, SD = .10), but this effect did not reach significance, p < .16 (Fig. 4b).
These results help generalize our findings in showing that associative activation relies on available resources and is state dependent. To examine whether the “bottleneck” for mental exploration is exclusively cognitive in nature, or whether other types of load similarly constrain the breadth of associative activation, in the next experiment we manipulated perceptual load.
Experiment 3: Perceptual Load Also Decreases Diversity in Associative Activation
Method
Subjects
Twenty-four students (19 females; mean age = 21.95 years, SD = 2.41) took part in this experiment in exchange for course credits or a monetary payment. Informed consent was obtained from all subjects.
Tasks
The free-association task was the same as in the previous experiments. Subjects also performed a task that was intended to limit their available perceptual resources. Subjects in the low-load condition were asked to attend to one feature (color) and respond by key press to each red letter they observed. Subjects in the high-load condition were asked to attend to a conjunction of features (color and shape) and respond by key press to each green L they observed (Fig. 5). In both conditions, 10% of the letter stimuli were targets. Stimuli for the two conditions were drawn from the same distribution, and only the instructions differed between the conditions.
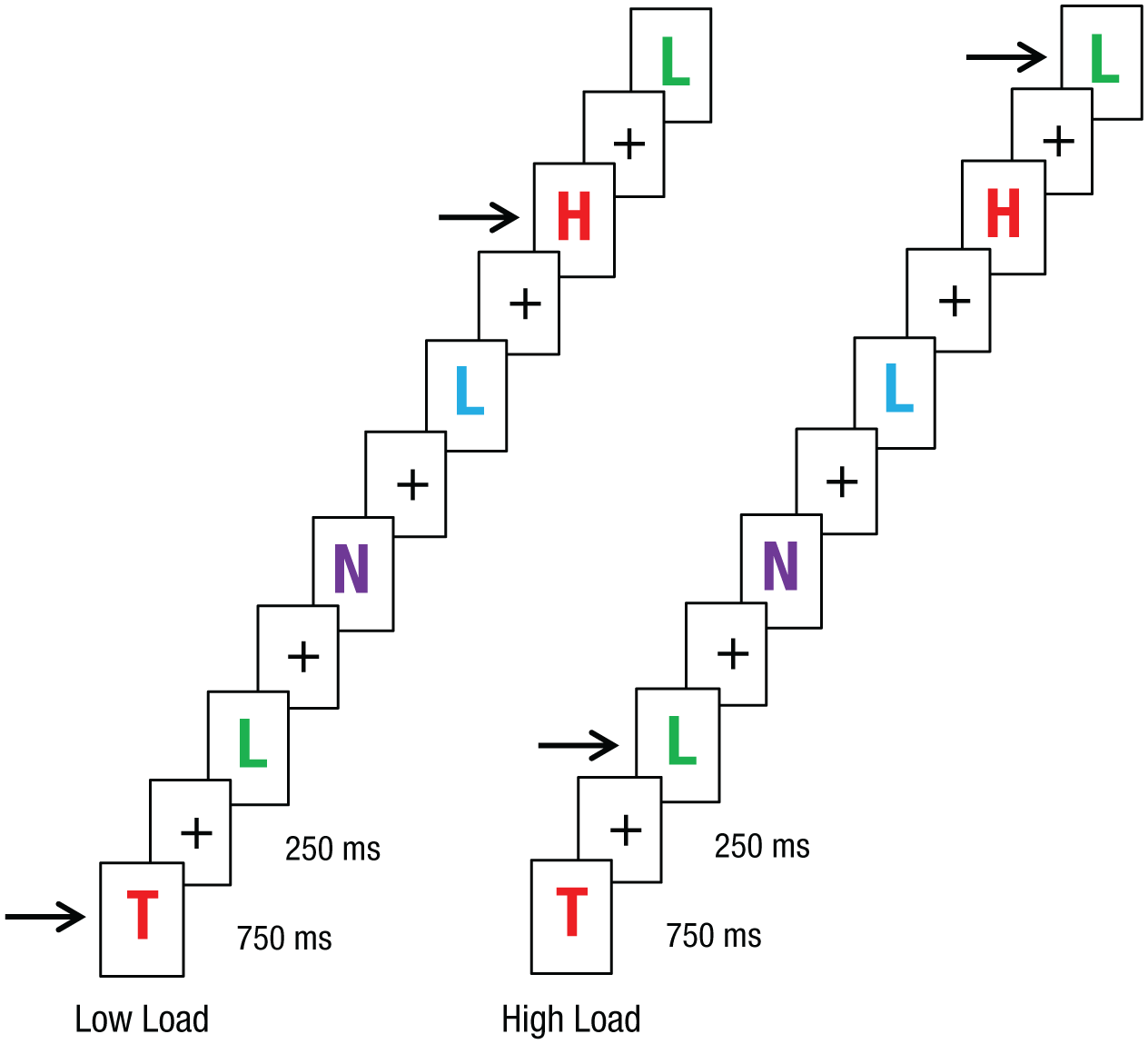
Illustration of the perceptual-load task in Experiment 3. Colored letters appeared on the screen in random order; each was presented for 750 ms and followed by a 250-ms presentation of a fixation cross. In the low-load (single-feature) condition, subjects responded by key press to each red letter they observed. In the high-load (feature-conjunction) condition, they responded to each green L they observed. Target letters are indicated by the arrows. Note that for simplicity of illustration, the displays for the free-association task are not shown here, although the stimuli for the two tasks were on-screen simultaneously.
Procedure
After signing consent forms, subjects were randomly assigned to either the high- or the low-load condition. The experiment began with brief instructions, which explained that subjects would be asked to respond to each target word with the first association that came to mind, while monitoring the screen and responding each time a target letter stimulus appeared. The target words were presented in random order, and each remained on-screen until an association was given. While subjects attended to the target words, random colored letters appeared on the screen, in a location between the target words and response box for the free-association task. Each letter appeared for 750 ms and was followed by a 250-ms presentation of a fixation cross; the timing of these stimuli was independent from the timing of the free-association task. Subjects were asked to respond by key press whenever they noticed a target letter. In both conditions, the experiment consisted of 10 blocks of 10 trials each. After completing the experiment, subjects were debriefed and thanked.
Results
Of the 24 students, 1 was excluded from analysis for being an RT outlier in the free-association task (> 2 SD from the mean). Therefore, 12 and 11 subjects in the low-load and high-load conditions, respectively, were included in the analyses.
Analysis of performance on the perceptual-load task revealed a significant difference between the conditions. Subjects in the high-load condition committed a higher proportion of false alarms (M = 0.01, SD = 0.01) than subjects in the low-load condition did (M = 0.001, SD = 0.002), t(11) = −2.23, p < .04. Sensitivity, as measured by d′, was higher in the low-load condition (M = 2.55) than in the high-load condition (M = 1.80). These results indicate that the experimental conditions indeed differed in their perceptual-load levels.
As in the previous experiments, RT in the free-association task did not differ significantly between the high- and low-load conditions (high load: M = 2.40 s, SD = 0.57; low load: M = 2.58 s, SD = 0.61), p > .25.
Analysis of the associations subjects provided also supported the results from the previous experiments. The dominant associations were provided more often in the high-load condition (M = .31, SD = .01) than in the low-load condition (M = .26, SD = .01), t(186) = −2.91, p < .004, Cohen’s d = 0.42. In addition, nondominant associations given for each target word decreased in frequency as load increased (high load: M = .69, SD = .01; low load: M = .75, SD = .01), t(195) = 3.01, p < .003, Cohen’s d = 0.43. The analysis of the proportion of maximal entropy revealed that this measure was significantly higher for answers given under low load (M = .83, SD = .11) than for answers given under high load (M = .77, SD = .15), t(184) = 3.11, p < .002, Cohen’s d = 0.45 (Fig. 6a).
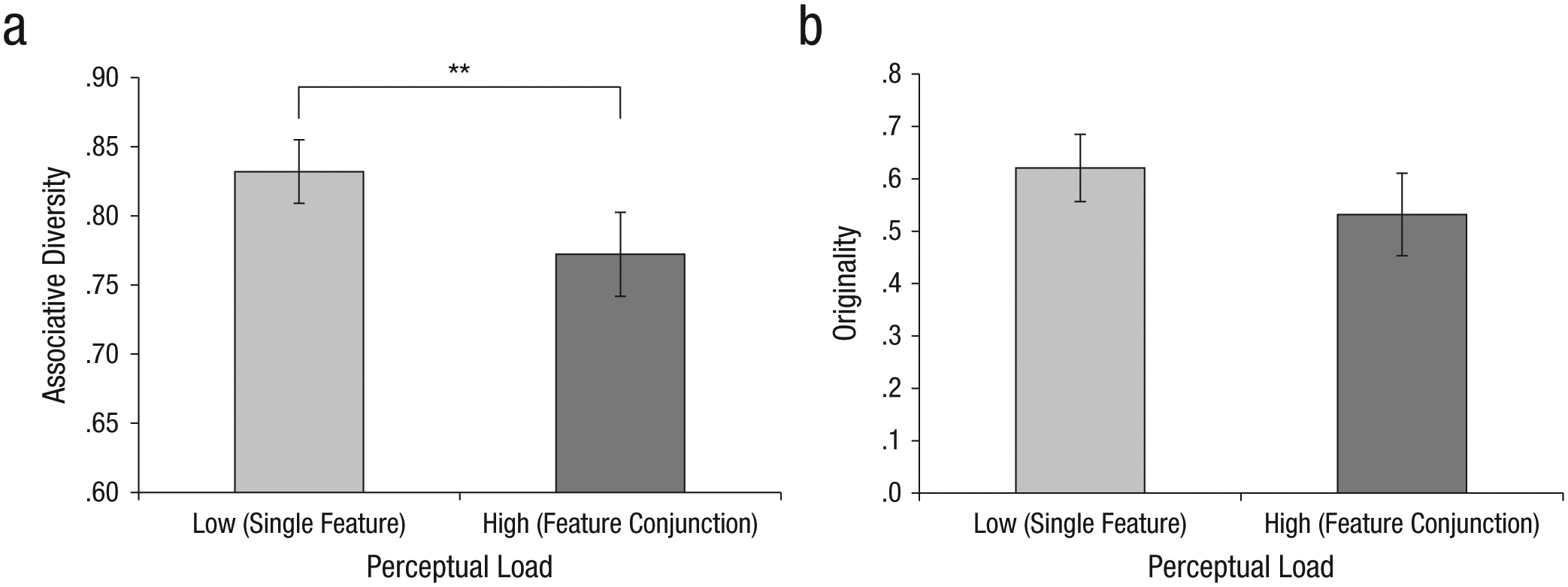
The effect of perceptual load on associative activation in Experiment 3. The graphs show (a) the proportion of associative diversity (measured by maximal entropy) and (b) mean originality scores (unique responses) in the low-load and high-load conditions. Error bars indicate 95% confidence intervals, and asterisks indicate significant differences between conditions (**p < .01).
Finally, although the difference was not significant, subjects’ originality scores were higher in the low-load condition (M = .62, SD = .10) than in the high-load condition (M = .53, SD = .11), p < .06 (Fig. 6b).
Taken together, our findings demonstrate that associative activation cannot be accounted for by associative strength alone, and that the breadth of associative activation is determined directly by cognitive and perceptual load. One important question that should be addressed before drawing conclusions is whether the subjects in the low-load conditions indeed activated remote associations as readily as the subjects in the high-load conditions activated consensual associations. They might instead have first activated the immediate, strongest associations—just as the subjects in the high-load conditions did—but achieved greater diversity in their answers because they then took additional time to search for more “interesting,” remote responses. To answer this question, we report a detailed RT analysis and a follow-up priming experiment that also revealed more about the mechanisms underlying associative activation under different loads (and modes).
RT Analysis Across Experiments
Although we did not find significant effects of RT on associative diversity in any of the experiments, we wanted to examine more closely whether greater diversity was associated with longer RTs across experiments. To do so, we imposed post hoc RT windows on the data, binning the answers according to whether they were provided within 2 s or within 3 s of the onset of the target word (for shorter windows, there were too few trials to be analyzed). In Experiments 1a, 1b, and 3, although a one-way multivariate analysis of variance (MANOVA) showed a significant effect of load on diversity, there were neither significant effects of response window nor significant interactions between response window and load (see Table 1 and Fig. 7). In Experiment 2, we did find a significant effect of response window on diversity. However, in this experiment, which involved task switching, longer RTs resulted in lower associative diversity. We provide a possible explanation for this intriguing result in the General Discussion. In summary, this post hoc response-window analysis supports the conclusion that increased load reduced associative diversity in the free-association task independently of changes in RT.
Summarized Effects of Load, Response Window, and Their Interaction on Associative Diversity in Experiments 1a, 1b, 2, and 3
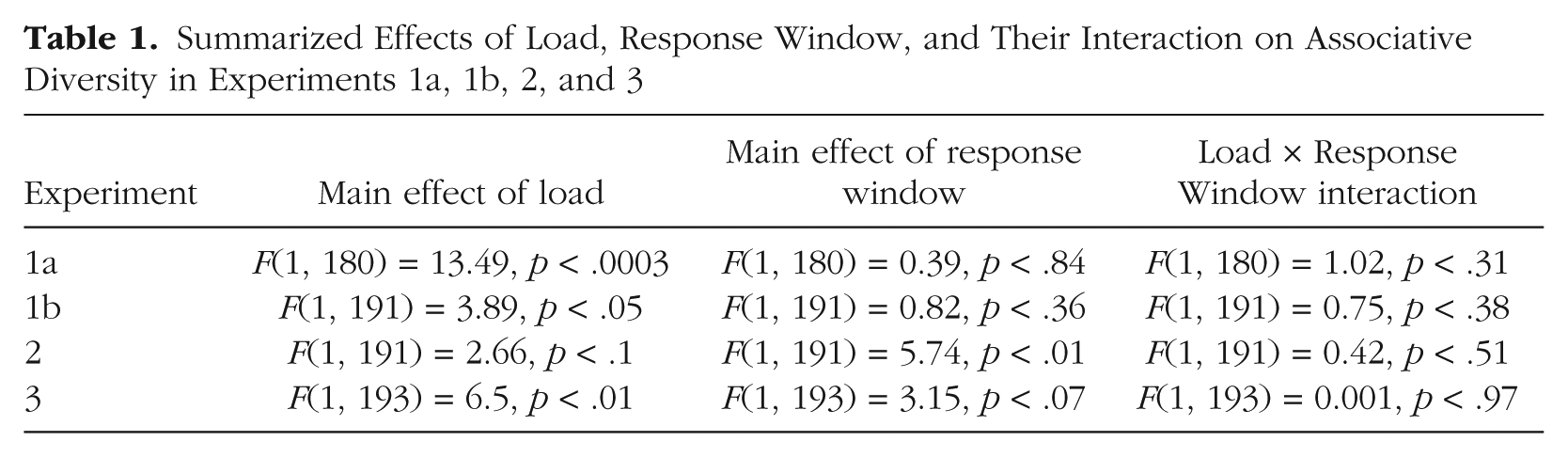
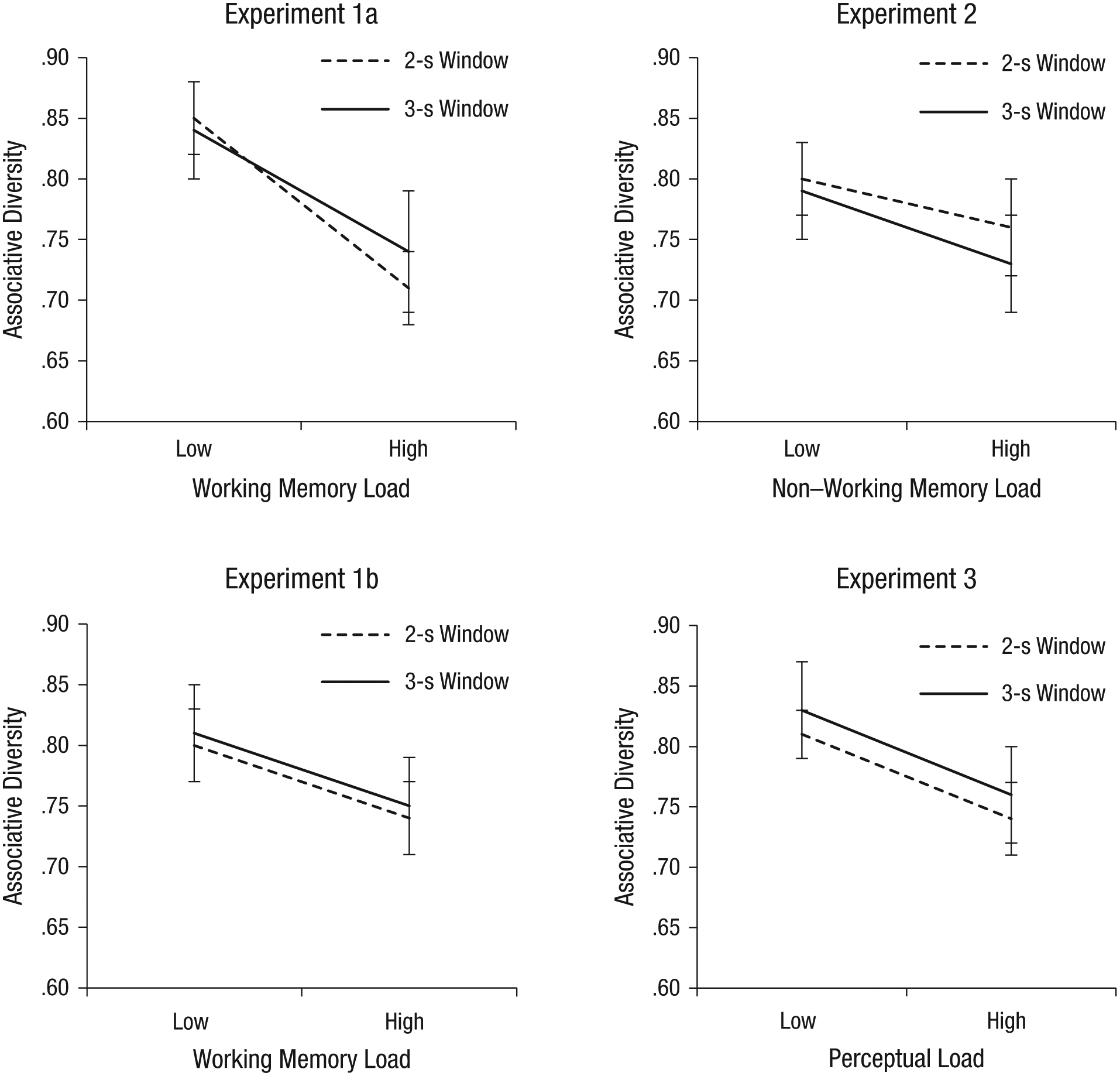
Associative diversity as a function of load and response window in Experiments 1a, 1b, 2, and 3. For each experiment, the graph shows the proportion of associative diversity (measured by maximal entropy) in the low- and high-load conditions, separately for responses provided within 2 s and responses provided within 3 s of the onset of the target word. Error bars indicate 95% confidence intervals.
Interim Discussion
We hypothesized that the availability of resources would affect how exploratory or exploitatory people were in performing a free-association task, a task assumed to reflect and rely on associative activation. This hypothesis challenges the common assumption that responses in such a task are governed by associative strength, and suggests instead that the activation of associations might be dictated by the availability of resources. Across our manipulations, increased load led to reduced variance of associations and reduced originality in subjects’ responses. Such dependence on load presumably promotes a more exploratory mode when resources are available and a more exploitatory mode when resources are scarce.
Our findings thus far cannot distinguish between two possible underlying mechanisms for the observed effects (see Fig. 8). On the one hand, it is possible that under conditions of low load, the more immediate associations were not activated at all or, alternatively, were inhibited in favor of exploratory activations of remote associations. But on the other hand, it is possible that the immediate associations were activated in parallel to the remote ones, and that the load changed the threshold for consciousness, rather than the strength of activation. In other words, we cannot rule out the possibility that under low load, immediate and remote associations were activated simultaneously, but only the remote associations were provided by our subjects (even if the equivalent RTs in the high- and low-load conditions make this possibility seem less likely).
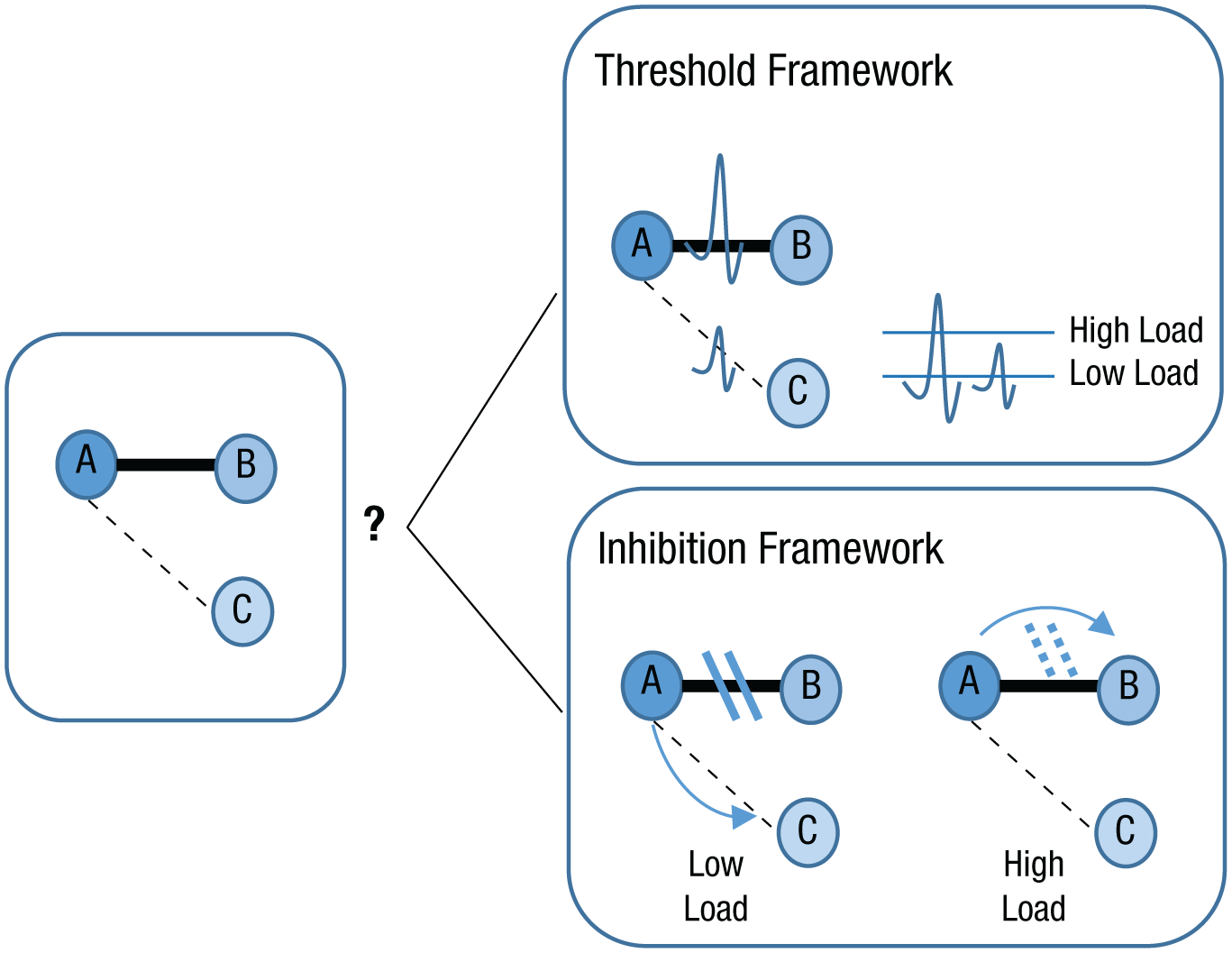
Proposed mechanisms for exploration and exploitation in associative activation. In this example, B is an immediate associate of A (indicated by thick lines), and C is a remote associate of A (indicated by dashed lines). In the illustration of the threshold framework, the waveforms indicate the relative levels of activation for the two associates of A. According to this framework, the threshold of activation needed for associations to reach conscious awareness (illustrated at the far right) is low when load is low (i.e., exploration mode) but high when load is high (i.e., exploitation mode). Thus, an immediate associate, such as B, reaches conscious awareness in both modes, but a remote associate, such as C, reaches conscious awareness only during exploration mode. According to the inhibition framework, when load is low, available resources strongly inhibit (solid parallel lines) immediate associates, and activation flows in the direction of remote associates. Thus, in exploration mode, remote associates reach conscious awareness. However, when load is high, inhibition is less efficient (dotted parallel lines) because of limited resources, so activation continues to flow toward immediate associates. Thus, in exploitation mode, only immediate associates reach conscious awareness, as predicted by the classic models of spreading activation.
As an initial test of these two alternatives, we designed a semantic-priming experiment in which both remote and immediate associations were primed. We reasoned that reduced activation of immediate associations in the low- compared with the high-load condition, as measured by priming, would support the account based on inhibition (or nonactivation) of immediate associations, whereas equivalent priming of immediate associations in the two load conditions would support the parallel-activation account.
Experiment 4: Semantic Priming of Immediate and Remote Associations Under Low and High WM Load
Method
Subjects
Fifty subjects (37 females; mean age = 22.81 years, SD = 2.42) participated in this study in exchange for a monetary payment.
Tasks
As in the first experiment, subjects performed an adapted digit-span task; in this case, they were asked to remember two digits in the low-load condition and five digits in the high-load condition. Subjects also completed a lexical decision task aimed at measuring semantic priming. On each trial of this task, a prime word was presented and followed by a probe stimulus, either a word or a pseudoword. Subjects were asked to report as quickly as possible whether the probe was a word. The primes were 288 words taken from the Hebrew word-association norms for college students collected by Rubinsten, Anaki, Henik, Drori, and Faran (2005). All the words and pseudowords were controlled for length. Words were also controlled for imaginability and frequency. Half of the probes were words, and half were pseudowords. On 25% of the trials with word probes, the prime word and probe were immediate associates (average association strength = .53; on another 25% of the trials with word probes, the prime and probe were remote associates (average association strength = .22); and on 50% of the trials with word probes, the prime and probe were unrelated.
Procedure
After signing consent forms and receiving brief instructions, subjects were randomly assigned to an experimental condition. The experiment began with a block of practice and continued with 29 experimental blocks. Each block began with a 3-s presentation of a digit string. Ten trials (or eight trials, in the last block) of the lexical decision task followed. In each trial, a prime word was presented for 250 ms, followed by a 100-ms mask and then a 250-ms presentation of the probe. As in the study by Heyman et al. (2015), subjects were instructed to read each prime silently and then to report as quickly and accurately as possible whether the probe was a real word or not. At the end of each lexical decision block, subjects were asked to type the digit string they had been asked to remember. After the 29 blocks were completed, the experiment ended, and subjects were debriefed and thanked
Results
Of the 50 subjects who took part in the study, 1 was excluded for being an outlier (RT > 2 SD above the mean). Therefore, the analyses included data from 24 subjects in the high-load condition and 25 subjects in the low-load condition.
Analysis showed that accuracy on the cognitive-load task differed significantly between the conditions. Subjects under low load were more accurate (M = .87, SD = .13) than those under high load (M = .73, SD = .16). Thus, the load manipulation was effective, t(43.89) = −3.31, p < .002. The two conditions did not differ significantly in accuracy on the lexical decision task (low load: M = .94, SD = .07; high load: M = .93, SD = .04), t(40.7) = −0.31, p > .25, or in RT (low load: M = 728.8 ms, SD = 105.36; high load: M = 724.62 ms, SD = 113.01), t(46.42) = −0.13, p > .25.
Next, to examine whether either immediate or remote associates were indeed primed, we conducted paired-samples t tests on RTs in each condition. In the high-load condition, as expected, immediate associates were significantly primed, t(23) = 8.6, p < 10−6, whereas remote associates were not, t(23) = 1.25, p < .22. In the low-load condition, significant priming was found both for immediate associates, t(24) = 6.69, p < 6 × 10−6, and remote associates, t(24) = 6.59, p < 8 × 10−7.
The main analysis was a one-way MANOVA, which tested whether the load manipulation had an influence on priming effects (i.e., shorter RTs for responses to probes that were remote or immediate associates of the primes compared with probes that were unrelated to the primes). Results showed a significant main effect for association strength, supporting the distinction between processing of remote and immediate associations. Immediate associates were primed to a greater extent than remote associates (M = 61.52 ms, SD = 42.58, and M = 25.76 ms, SD = 38.41, respectively), and this effect was significant, F(47, 1) = 33.17, p < .001, η p 2 = .41.
More important, the analysis showed a significant interaction between load and association strength, F(47, 1) = 25.65, p < .001, η p 2 = .35. Furthermore, pairwise comparisons (Bonferroni corrected for multiple tests) revealed a double dissociation between the effects of load on priming of immediate and remote associates: The semantic priming of immediate associates that was found under low load (M = 45.11 ms, SD = 7.89) was heightened under high load (M = 78.62 ms, SD = 8.06), and this difference was significant, F(47, 1) = 8.81, p < .005, η p 2 = .15 (Fig. 9a). In contrast, the semantic priming of remote associates that was found under low load (M = 40.71 ms, SD = 7.11) was diminished under high load (M = 10.19 ms, SD = 7.25). This difference was significant as well, F(47, 1) = 9.02, p < .004, η p 2 = .16 (Fig. 9b). The finding that subjects under low load showed any priming at all for the immediate associates is a reflection of the fact that not all subjects were equally original under low load; indeed, some provided consensual responses under low load.
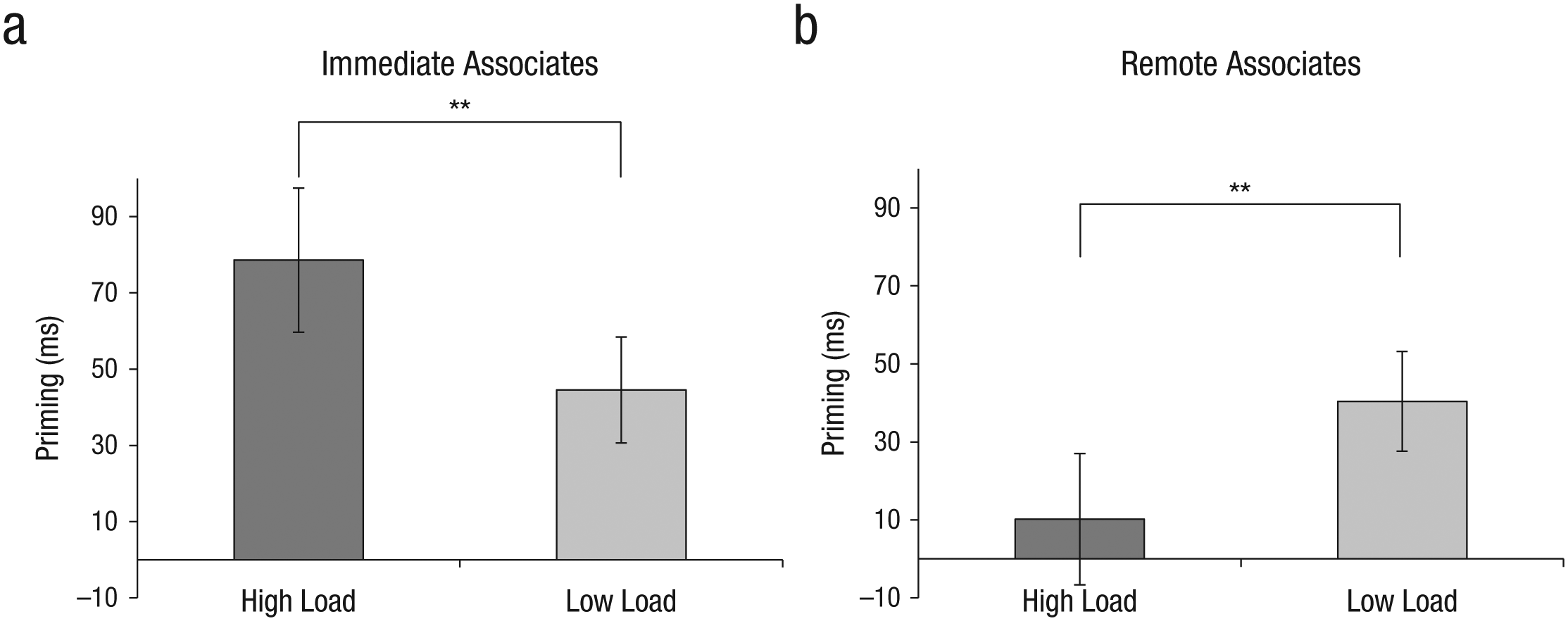
Semantic priming of (a) immediate and (b) remote associates in the high- and low-load conditions of Experiment 4. Priming was calculated as the difference in reaction time between responses to probes that were related to the prime and responses to probes that were unrelated to the prime. Error bars indicate 95% confidence intervals, and asterisks indicate significant differences between conditions (**p < .01).
The significant increase in priming of the remote associates along with the significant decrease in priming of the immediate associates in the low-load condition implies that when cognitive resources are available, activation of immediate and consensual associations is inhibited, or these associations are not even activated, in favor of activation of remote and unique associations.
General Discussion
The aim of this study was to examine whether associative thinking patterns can be more exploratory or more exploitatory depending on the availability of resources. We found that, just like perception, inner mental processes are indeed state dependent, and furthermore, that associative activation is less automatic than previously believed. Specifically, our findings in the first three experiments show that increased load leads to a reduction in associative variance, and to greater reliance on immediate associations in a free-association task. Further analysis and a fourth experiment indicate that the diversity and originality in associative activation under low load is not due to longer search for interesting responses, but rather is due to low load favoring the activation of remote associations. Our results indicated that in task-switching settings, longer RTs correlated with reduced diversity. We reason that under such task demands, delayed responses are indicative of higher load, rather than of longer search for interesting responses.
Taken together, these studies suggest that the main prediction of the spreading-activation framework—that activation is related to the strength of association—is more accurate when resources for exploration are depleted than when they are fully available. Our findings support the notion that exploration is the default mode: The brain has a basic tendency to go beyond the nearest associations and activate unique ones instead when resources are available.
An intriguing question raised by our results is why subjects in the low-load conditions tended to favor less consensual, more explorative associations even though the immediate, strongest associations would have been easier and faster to retrieve? What was the incentive to explore when there was no load? The originality observed in the low-load conditions is particularly remarkable given that subjects received no reward for creativity.
We suggest that this tendency toward exploration may arise from the involvement of reward-related mechanisms. It has been shown that attention to novelty in the perceptual environment is supported by brain areas related to reward (Bunzeck, Dayan, Dolan, & Düzel, 2010; Wittmann, Bunzeck, Dolan, & Düzel, 2007), and that the involvement of learning circuits (Bunzeck & Düzel, 2006) and reward circuits (Guitart-Masip, Bunzeck, Stephan, Dolan, & Düzel, 2010) is enhanced in the context of coding novelty. This line of research suggests that the quest for new information is rewarding in itself (Biederman & Vessel, 2006). We suspect that with respect to reward, attending to unique mental stimuli, such as remote associations, is not different from attending to novelty in the external environment, and that this reward value may drive the activation of diverse associations when possible.
We have suggested two possible mechanisms to account for our findings that reduced load allows subjects to go beyond consensual associations and rapidly access more diverse ones, without having to take more time to answer. According to one account, load does not affect associative activation per se, but rather changes the threshold of activation needed for conscious access. Under low load, presumably an exploratory state, all associations are activated in parallel, and a low threshold of activation allows remote associations to compete for access. However, under high load, a more exploitatory state, all associations are activated according to their relative strength, and only immediate activations cross the threshold. Indeed, some evidence has demonstrated that load modulates conscious perception of both visual (Carmel, Saker, Rees, & Lavie, 2007) and auditory (Macdonald & Lavie, 2011) sensory stimuli. However, our fourth experiment does not support this account because priming of immediate associates was significantly reduced in the low-load compared with the high-load condition.
An alternative account is that exploration of remote associations is enabled through inhibition of immediate associations. Under high load, resources for such inhibition are limited, and the strongest associations are uncontrollably triggered, by the rules of spreading activation. Such exploration via inhibition allows a balance between exploration and exploitation, between learning when learning is possible and reliance on stored information when resources are low. This account is supported by the differential priming found in our fourth experiment: Increased processing of remote associations was accompanied by decreased processing of immediate associations.
Our results suggest that exploration is the default mode. This idea is supported by the literature on mind wandering, which shows that when no other goal is to be achieved, people engage in spontaneous thoughts, as if exploring their internal world of memories and simulations (Andrews-Hanna, Reidler, Huang, & Buckner, 2010; Bar, Aminoff, Mason, & Fenske, 2007; Schooler et al., 2011). Here, we have shown that when there are no taxing external demands, the first conscious association can be the product of inner exploration, and need not simply be the strongest association triggered, as was claimed previously.
The findings reported here also have potential clinical implications, particularly regarding anxiety and depression. One of the dominant behaviors experienced in mood disorders is rumination, that is, dwelling on the same thought repeatedly. We suggest that ruminations are akin to cognitive load, and thus result in a perpetually exploitatory state. Hence, depression narrows people’s associations (Bar, 2009). Indeed, Isen, Johnson, Mertz, and Robinson (1985) found that induced positive affect leads to more diverse associations. We suggest that different moods serve as contexts for explorative and exploitative behaviors. Moreover, if exploration in mental processes involves the reward system, this involvement might be down-regulated in mood disorders that are accompanied by ruminative thinking. Providing conditions for engagement in remote associations may have therapeutic results in such cases. This hypothesis is in line with previous results showing that mere engagement in remote associations improves mood (Mason & Bar, 2012).
Conclusions
The experiments reported here support two main proposals. One is that the activation of associations is state dependent. Activation can be selectively drawn to distinct and interesting associations in an explorative manner, or can be determined by associative strength in an exploitative manner. Second, the availability of brain resources, whether cognitive or other kinds of resources, is a key factor in biasing which associations are activated. We have reviewed experimental findings regarding exploitation and. exploration in processes of attention to the external, perceptual environment, and our experiments demonstrate that the balance between exploitation and exploration is maintained with regard to internal mental processes as well. Further research is required to shed light on the mechanisms motivating and mediating this balance. The findings presented here have theoretical implications as well as possible implications in clinical settings.
Footnotes
Acknowledgements
We are grateful to Einav Sudai for helpful discussions and comments.
Action Editor
Edward S. Awh served as action editor for this article.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
The research was supported by The Israeli Center of Research Excellence in Cognition, Grant 51/11.