
Abstract
For decades, there has been controversy about whether forgetting is caused by decay over time or by interference from irrelevant information. We suggest that forgetting occurs because of decay or interference, depending on the memory representation. Recollection-based memories, supported by the hippocampus, are represented in orthogonal patterns and are therefore relatively resistant to interference from one another. Decay should be a major source of their forgetting. By contrast, familiarity-based memories, supported by extrahippocampal structures, are not represented in orthogonal patterns and are therefore sensitive to interference. In a study in which we manipulated the postencoding task-interference level and the length of the delay between study and testing, we provide direct evidence in support of our representation theory of forgetting. Recollection and familiarity were measured using the remember/know procedure. We show that the causes of forgetting depend on the nature of the underlying memory representation, which places the century-old puzzle of forgetting in a coherent framework.
What causes forgetting? This question has challenged scholars since the beginning of memory research (Ebbinghaus, 1885/1913). Early students of memory presumed that memories decay and fade with time, but this idea was challenged by evidence pointing to the deleterious effects of interference on memory (McGeoch, 1932; Underwood, 1957). This evidence suggested that memories are made less accessible because of interference from similar information acquired before or after their formation. The interference account was supported by an abundance of empirical evidence, but the decay account continued to retain adherents (for a review, see Wixted, 2004). Thus, the cognitive literature currently lacks a coherent unifying account for the causes of forgetting (Wixted, 2004). 1 In a recent review (Sadeh, Ozubko, Winocur, & Moscovitch, 2014), however, we reported evidence for a representation theory of forgetting, which posits that the primary cause of forgetting—interference or decay—depends on the nature of the initial memory representation.
Our representation theory of forgetting stems from recent neuropsychological investigations of the hippocampus. The hippocampus is a pivotal mnemonic structure that, through its different subfields, plays important roles in linking memories that share similar properties (e.g., similar contexts), as well as in distinguishing between similar memories (McKenzie et al., 2014; Zeithamova, Dominick, & Preston, 2012). The latter role, accomplished by pattern separation, has been shown to be mediated specifically by the dentate gyrus and cornu ammonis 1 (CA1) subfields of the hippocampus (Yassa & Stark, 2011). Pattern separation refers to the unique ability to reduce interference among similar memories 2 (Kuhl, Shah, DuBrow, & Wagner, 2010) by forming orthogonal, nonoverlapping representations of them (Hardt, Nader, & Nadel, 2013; Leutgeb, Leutgeb, Moser, & Moser, 2007; O’Reilly & Norman, 2002). Although interference among hippocampus-dependent memories may be reduced in this way (compared with extrahippocampal memories), it has been suggested that hippocampally represented memories are sensitive to decay (Hardt et al., 2013). At the cellular level, decay may result from remodeling of hippocampal circuits as a result of neurogenesis (Frankland, Köhler, & Josselyn, 2013) or from the loss of synaptic connections by intracellular processes (Nader, Schafe, & LeDoux, 2000). By contrast, other types of memory that rely on extrahippocampal structures, such as the perirhinal cortex, would be more susceptible to interference, which would be the major source of forgetting for these types of memory.
Two approaches have guided current thinking regarding the functional distinction between hippocampal and extrahippocampal memories. The first approach maintains that the hippocampus supports a mnemonic process termed recollection, whereas extrahippocampal structures, especially the perirhinal cortex, support a process termed familiarity (Diana, Yonelinas, & Ranganath, 2007). Recollection is a mnemonic process that involves reinstatement of memory traces within the context in which they were formed. Familiarity is a mnemonic process that manifests itself in the feeling that a studied item has been experienced, but without reinstating the original context (e.g., the feeling that a certain person is well known, even though no additional episodic details about that person can be recalled).
According to the second approach, the hippocampus is essential to representing particularly strong memories, whereas extrahippocampal memories, associated in particular with the perirhinal cortex, are sensitive to differences between weaker memory traces. Despite many differences between these two views, they share the notion that the hippocampus represents memories in a distinct manner from the perirhinal cortex (or for that matter any other extrahippocampal structure). Hence, we predicted that different memory representations, whether distinguished by the former approach or by the latter, would undergo different patterns of forgetting. The aim of the current study was not to distinguish between these two approaches, but rather to provide evidence that the causes of forgetting vary according to the nature of the underlying memory representation.
In our previous review of the effects of delay and interference on forgetting, we reported evidence for our representation theory of forgetting (Sadeh et al., 2014). The present literature, however, includes only studies that manipulated either interference or decay. Such studies cannot clearly tease apart the effects of decay or interference. Thus, decay manipulations typically involve several retention intervals of various lengths, and these most likely include a certain degree of interference from external stimuli or from internal thoughts. Likewise, high-interference tasks necessarily involve a decay component, which varies according to the duration of the tasks and the study-test interval. Clearly, there is a need for a systematic, within-study comparison of the manner in which interference and decay exert their effects on different memory representations.
In the current study, we examined the effects of decay and interference on forgetting of different memory representations by comparing memory performance in an immediate test with performance in a later test. To obtain estimates of recollection and familiarity, we used the remember/know paradigm (Tulving, 1985) in both initial and final tests. We used confidence ratings to examine differences between stronger and weaker memories. Forgetting levels were compared across two levels of interference while equating the duration of the two interference tasks. In addition, forgetting was examined across two levels of decay (short and long retention intervals) with little interference in each interval.
Method
The study consisted of three phases: (a) continuous recognition, (b) retention, and (c) final recognition test. To examine the effects of interference and delay on forgetting, we manipulated the nature of the retention phase between subjects, forming three groups. The first group (long-delay/high-interference) was presented with an interference task for approximately 15 min. The second group (long-delay/low-interference) was given a task with little interference, and this task also took approximately 15 min to complete. The third group (short-delay/low-interference) was given the same task as the long-delay/low-interference group, but for only 2 min. During the final recognition test, we measured the degree to which memory had declined. The contributions of recollection and familiarity in the continuous recognition phase and in the final recognition phase were estimated using subjective ratings (the remember/know paradigm; Tulving, 1985). In addition, confidence ratings were collected in both tests.
Participants
Participants were 272 3 students from the University of Toronto who completed the experiment in exchange for $10 or course credit. Of these, 77, 104, and 91 participants were assigned to the long-delay/high-interference, long-delay/low-interference, and short-delay/low-interference groups, respectively.
Materials
Six hundred sixty words were selected from the free-association norms of Nelson, McEvoy, and Schreiber (2004). Words were between 5 and 10 letters long and had a mean Kucera-Francis word frequency (Kucera & Francis, 1967) of 84.26 (SD = 137.13). For each participant, 100 words were randomly selected; each word would be seen twice, the second time 8 to 12 trials after the first presentation. Because the order of the words and their repetition lag was fully randomized, some filler words (i.e., words that were presented only once) were needed to fill in some trials between repetitions. Approximately 20 filler words were randomly selected for each participant.
For the music tasks, recordings of 160 classical music pieces for guitar, piano, violin, and trumpet were downloaded from http://www.free-scores.com/index_uk.php3. There were 40 recordings for each instrument. A 2-s clip was randomly extracted from each recording.
Procedure
The procedure for all three phases is illustrated in Figure 1. Phase 1 of the experiment was a continuous recognition task. A single word appeared in the center of the screen on each trial. After 1 s, a confidence scale ranging from 6 (sure old) to 1 (sure new) appeared below the word. Participants were instructed to indicate whether each word was old (i.e., it had appeared earlier in the experiment) or new by selecting the appropriate confidence rating. After they made a confidence rating, the confidence scale disappeared, but the word remained on-screen, and participants were prompted to indicate whether the word was reexperienced, 4 familiar, or neither; the last of these indicated that it was judged as a new word. A 0.5-s interstimulus interval preceded the next trial.
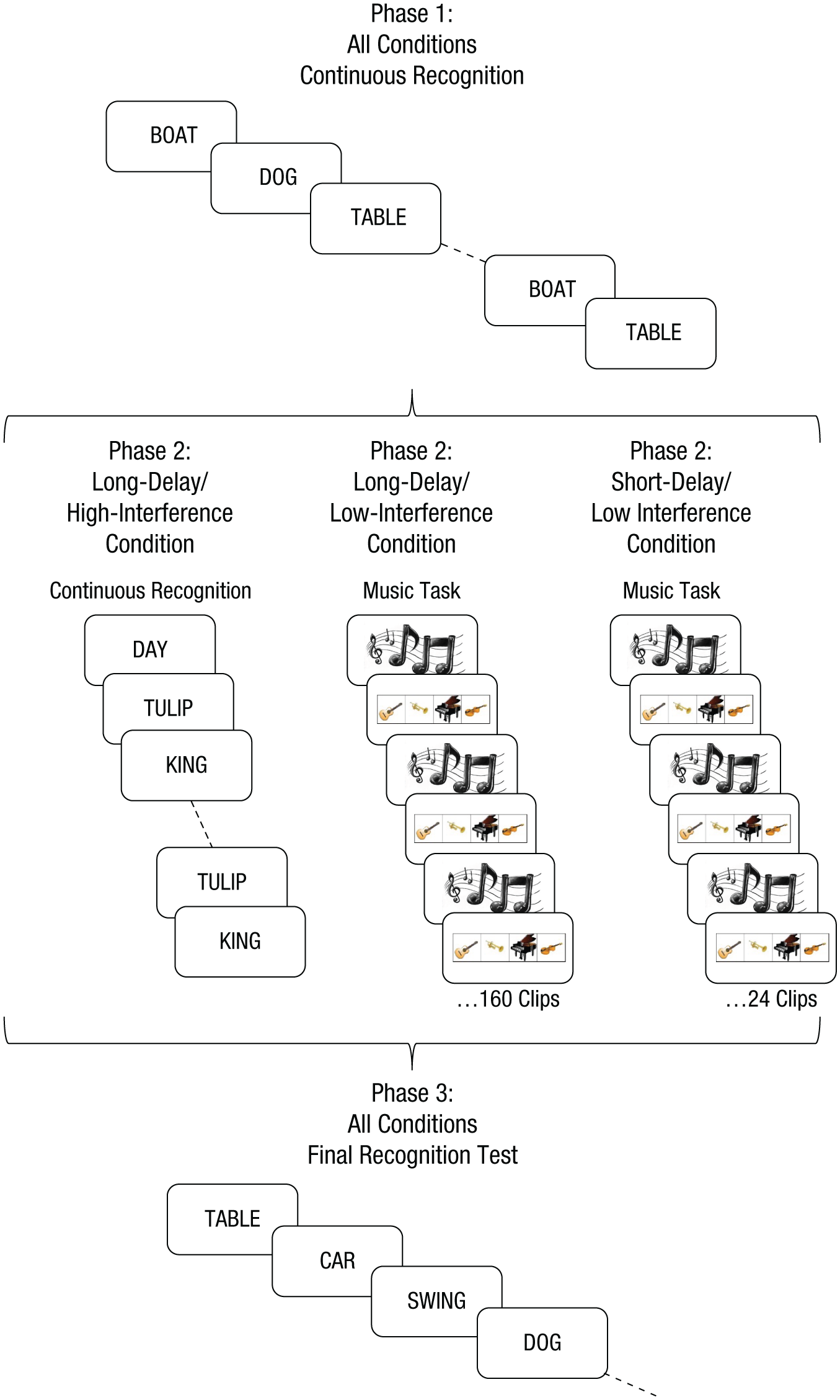
Illustration of the three phases of the experiment for the three conditions. During Phase 1, in all conditions, single words appeared sequentially in the center of the screen. Participants rated how confident they were that each word had already appeared in the experiment and gave a subjective rating of the experience the word evoked (”re-experience,” “familiar,” or “neither”; see Method section). Phase 2 differed across conditions. In the long-delay/low-interference and short-delay/low-interference conditions, short music clips (160 clips and 24 clips, respectively) were presented. An image of musical notes appeared on the screen while each clip played (2 s). After each music clip, images of four possible musical instruments (i.e., guitar, trumpet, piano, violin) appeared on the screen, and participants decided which instrument they had heard in that clip. Phase 2 of the long-delay/high-interference condition was identical to Phase 1, except that a new set of words was presented. The contrast between the short-delay/low-interference and long-delay/low-interference conditions provides a measure of memory decay, whereas the comparison between the long-delay/low-interference and long-delay/high-interference conditions provides a measure of interference. Phase 3 was a final recognition test for words that had appeared in Phase 1. As in Phase 1, participants gave confidence ratings and responded to each word with “re-experience,” “familiar,” or “neither.”
During Phase 2 (the retention phase), participants in the long-delay/high-interference condition were presented with an additional continuous recognition task that was very similar to that used in the first phase, except that a second set of words was used, none of which were used in Phase 1 (see Materials). Participants in the long-delay/low-interference condition were presented with 160 2-s music clips; after each clip, participants identified which instrument—guitar, piano, violin or trumpet—was playing (see Fig. 1). There was no time limit for participants’ responses. A 1-s inter-stimulus interval preceded presentation of the next music clip. The short-delay/low-interference condition was identical in all aspects to the long-delay/low-interference condition, except that only 24 music clips were presented. Phase 2 for the short-delay/low-interference condition lasted approximately 5 min (including instructions); Phase 2 for the long-delay/low-interference and long-delay/high-interference conditions lasted approximately 15 min (including instructions).
Phase 3 (the final recognition phase) included all 100 words that had been repeated in the initial continuous recognition phase, as well as 100 new words that were randomly selected from the word pool. Participants were informed that this phase of the experiment was a memory test for the words that they had learned in the first phase of the experiment and were reminded of the task instructions (i.e., old/new confidence ratings and reexperience, familiar, or neither ratings). The presentation order of old and new words was randomized. Each trial in the final recognition phase operated identically to those in the continuous recognition phase: Words were shown for 1 s, followed by the old/new confidence rating, and then a subjective rating regarding recollection and familiarity, with an interstimulus interval of 0.5 s.
After this phase, the experiment ended and participants were thanked, debriefed, and paid. The entire experiment took no longer than 1 hr for the long-delay/high-interference and long-delay/low-interference groups, and approximately 45 min for the short-delay/low-interference group.
Data analysis
We hypothesized that the post-encoding processes that influence forgetting are dependent on the type of representation that supported memory initially (i.e., before these representations were subjected to alterations by post-encoding processes). Therefore, in all analyses, the percentages of recollected and familiar words (see note 4) were derived from hits in the continuous recognition test (see the Discussion section for the possible ramifications of this approach).
Memory accuracy was indexed by corrected recognition: the probability of detecting studied items, adjusted for the false alarm (FA) rate (i.e., p(hit) − p(FA)). We used a modified version of the measure of discriminability (following Duarte, Graham, & Henson, 2010). For recollected words, we multiplied the percentage of recollection responses by the percentage of hits minus the percentage of false alarms— p(R) × [p(hit) − p(FA)] (see Table 1). This calculation was performed once for corrected recognition in the continuous recognition test and once for corrected recognition in the final test (but for both tests, percentages of recollected words were taken from the continuous recognition test). For illustration of this calculation for the final test, consider a hypothetical participant who judged 60% of the correctly recognized items in the continuous recognition test as recollected, correctly detected 80% of the old items in the final test, and falsely recognized 15% of the new items in the final test. For that participant, the measure of discriminability for the recollection estimate would be
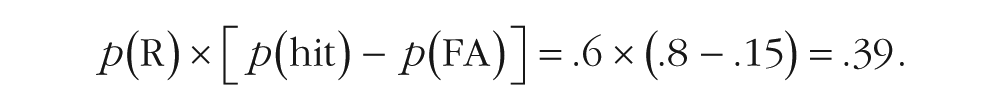
Performance in the Continuous Recognition Test (Phase 1)
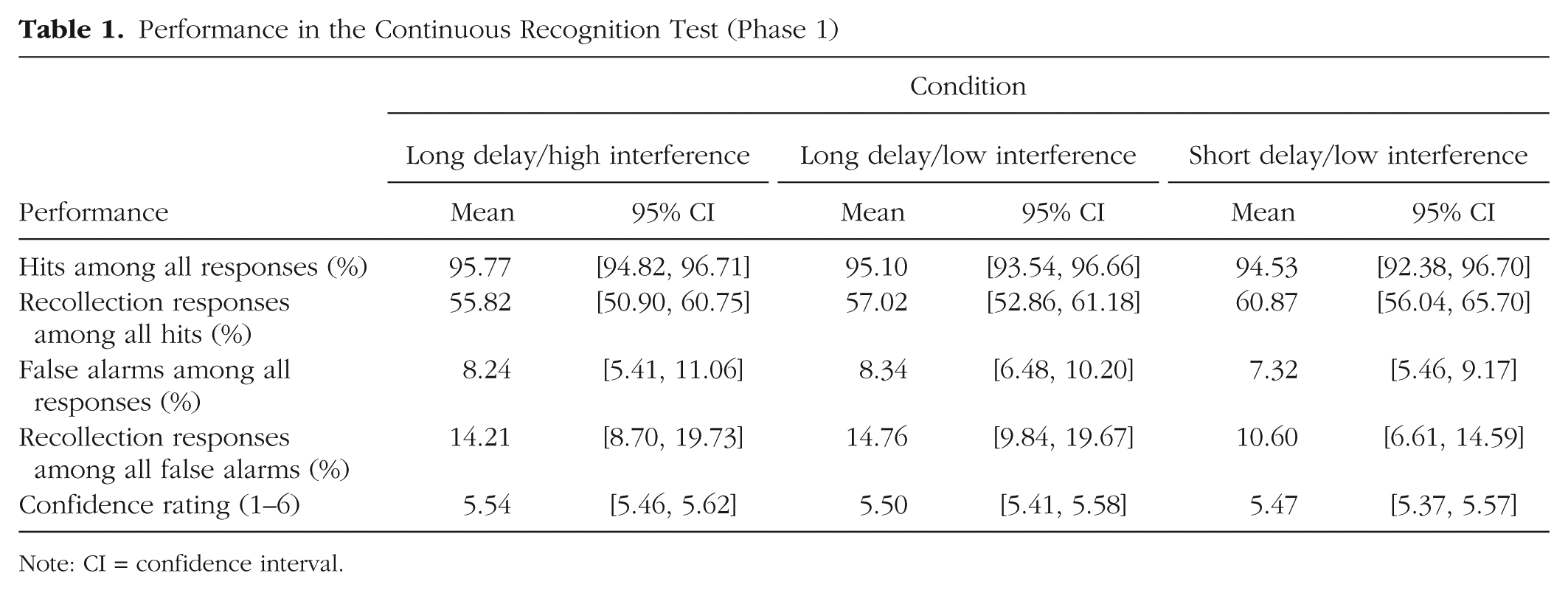
Note: CI = confidence interval.
Furthermore, we accounted for the independence between recollection and familiarity (according to certain models, e.g., Yonelinas & Jacoby, 1995, but not others, e.g., Ingram, Mickes, & Wixted, 2011; for a recent review advocating the independence measure, see Migo, Mayes, & Montaldi, 2012). In addition to its theoretical importance, independence allows measures of recollection and familiarity estimates to be used as factors in the same analysis of variance (ANOVA). We thus calculated hits and false alarms, respectively, as follows:
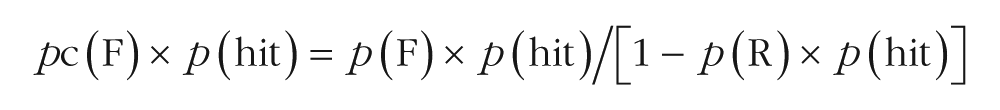
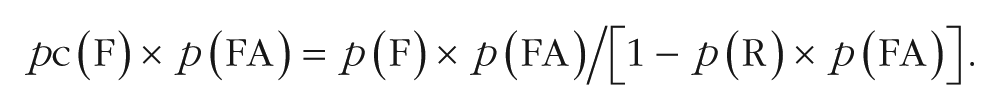
In these equations, pc(F) is the percentage of familiar-word hits in the continuous recognition test, corrected for independence (i.e., the familiarity estimate), and p(F) is the percentage of familiar-word hits in the continuous recognition test, uncorrected for independence. The corrected recognition score for the familiarity estimate was thus pc(F) × p(hit) − pc(F) × p(FA).
For each participant, these computations resulted in one measure of memory performance weighted by his or her estimate of recollection in the continuous recognition test, and one weighted by his or her estimate of familiarity. To reiterate, we performed the same calculations for the continuous recognition test, but for that test, recollection and familiarity estimates and hit and false alarm rates were derived from the same phase. We henceforth refer to these measures of corrected recognition weighted by estimations of recollection and familiarity as weighted corrected recognition (termed probability estimates by others; see Duarte, Graham, & Henson, 2010).
Results
Continuous recognition phase
The results of the first phase of the experiment, the continuous recognition test, are summarized in Table 1. As expected, no significant differences were found among the three conditions (long-delay/high-interference, long-delay/low-interference and short-delay/low-interference) with regard to any of the measures, all F(2, 269)s < 1, ps > .250. The high percentage of hits in the continuous recognition test is in line with previously reported results (see Experiment 1 in Hintzman, 2010).
Differences in memory performance between the continuous recognition and final tests
To examine the effects of decay and interference on memory, we compared the difference in memory performance between the continuous recognition test and the final test across the three experimental groups. Thus, forgetting is indexed by the difference in weighted corrected recognition between the continuous recognition test and the final test. Decay was operationalized as the comparison between the long-delay/low-interference condition and the short-delay/low-interference condition, given that the nature of the interfering task was held constant in these two conditions and only the duration of the interfering task differed. Interference was operationalized as the comparison between the long-delay/high-interference condition and the long-delay/low-interference condition, given that the length of the interfering task was equivalent in these two conditions and only the nature of the interfering materials differed. Memory performance was indexed by corrected recognition weighted by recollection and familiarity estimates, as detailed in the Method section. Following recent recommendations (Bakker & Wicherts, 2014; Leys, Ley, Klein, Bernard, & Licata, 2013; Wilcox, 2009), we used the median absolute deviation (MAD) as a robust method for outlier detection. The criterion was set a priori as any value that was more than 3 MAD from the median. Using this criterion, we found no outliers with regard to the percentage of recollected words in the continuous recognition test. Twelve outliers were found with regard to corrected recognition in the continuous recognition test: three in the short-delay/low-interference condition, six in the long-delay/low-interference condition, and three in the long-delay/high-interference condition. The data of the corresponding participants were removed from all subsequent analyses.
Figure 2 presents the difference in weighted corrected recognition between the two tests for the recollection estimates and the familiarity estimates across all conditions. To test the significance of this pattern of results, we submitted the data to a 2 × 2 × 3 ANOVA with response (recollection estimate, familiarity estimate) and test (continuous recognition, final recognition) as within-subjects factors and condition (short-delay/low-interference, long-delay/low-interference, long-delay/high-interference) as a between-subjects factor. The results are presented in Table 2. Most important, the ANOVA statistically confirmed that the patterns of forgetting for recollection and familiarity differed among the three conditions, given that the three-way interaction of response, test, and condition was significant.
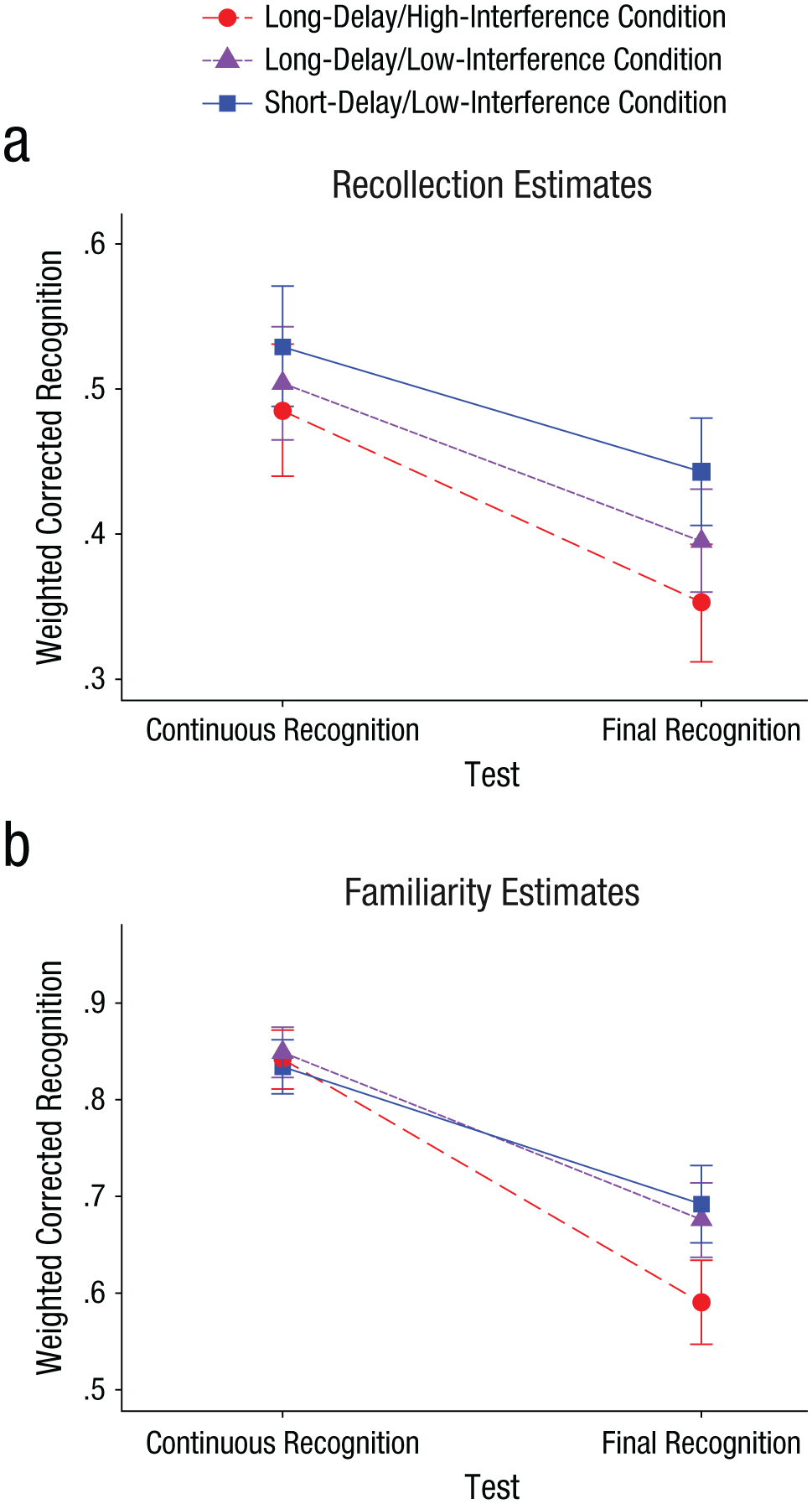
Corrected recognition as a function of test, weighted by (a) recollection estimates and (b) familiarity estimates, presented separately by condition. Error bars represent 95% confidence intervals around the means.
Analysis of Variance Results: Effects of Condition, Test, and Response on Weighted Corrected Recognition
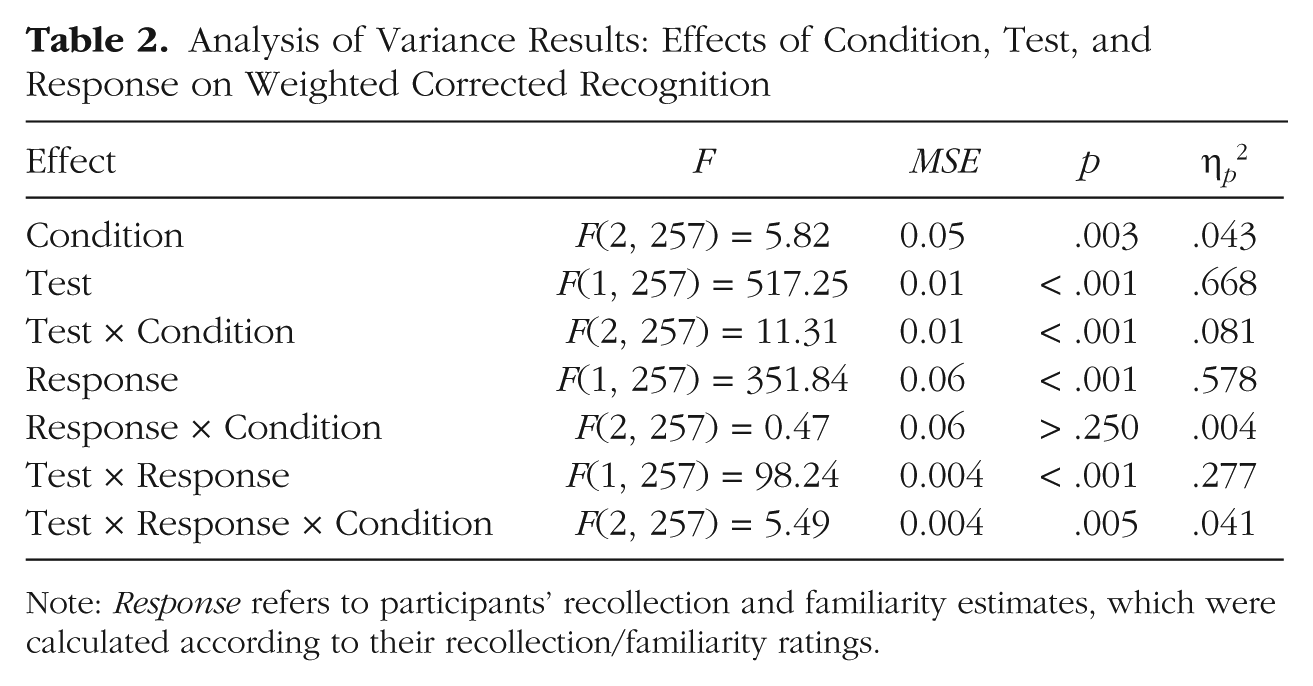
Note: Response refers to participants’ recollection and familiarity estimates, which were calculated according to their recollection/familiarity ratings.
Solid statistical support for our hypothesized pattern of results would require the following three outcomes: (a) differential effects of decay and interference on the recollection and familiarity estimates, (b) a decay effect on the recollection estimates, and (c) an interference effect on the familiarity estimates. The prediction of differential effects of decay and interference was confirmed by the significant three-way interaction between condition, test, and response, as elaborated earlier. To confirm the other two predictions, we conducted two separate 2 × 2 ANOVAs with test as a within-subjects factor and condition as a between-subjects factor. The first ANOVA included only corrected recognition weighted by the recollection estimates as the dependent variable and only the short-delay/low-interference and long-delay/low-interference conditions: thus, it focused specifically on a decay effect on recollection. The second ANOVA included only corrected recognition weighted by the familiarity estimates as the dependent variable and only the long-delay/low-interference and long-delay/high-interference conditions, thus focusing specifically on an interference effect on familiarity. Because our hypothesis entailed a union of all three outcomes, we computed a combined p value akin to that obtained using the Fisher technique (Fisher, 1973; see also Slotnick & Schacter, 2004). 5 This computation resulted in a statistically significant result (combined p = .008), confirming that the pattern of results was in line with our a priori hypothesis.
An additional important prediction of our theory is that memories dependent on the hippocampus will be less sensitive to interference than memories dependent on extrahippocampal structures. To statistically test this prediction, we ran an additional ANOVA on the data in which we included only the long-delay/low-interference and long-delay/high-interference conditions, thus focusing on the effects of interference. A significant three-way interaction between condition, response (recollection estimate, familiarity estimate) and test (continuous recognition, final recognition) was found, statistically confirming our prediction that recollection would be less sensitive to interference than familiarity, F(1, 170) = 8.01, MSE = 0.03, p = .005, η p 2 = .045 (see the pattern in Fig. 2).
We ran an analysis in parallel to the 2 × 2 × 3 ANOVA; in the new analysis, participants’ responses in the continuous recognition test were divided according to their confidence ratings rather than according to whether they had reported recollecting or being familiar with the word. We calculated the proportion of words given a confidence rating of 6 (high confidence) and the proportion of words given a confidence rating of 4 or 5 (low confidence) out of the total number of hits in the continuous recognition test (words given ratings of 1–3 were considered to be “new” responses). Examination of the results (Fig. 3 and Table 3) revealed that the overall pattern of the data was similar to that when they were analyzed according to the recollection/familiarity division.
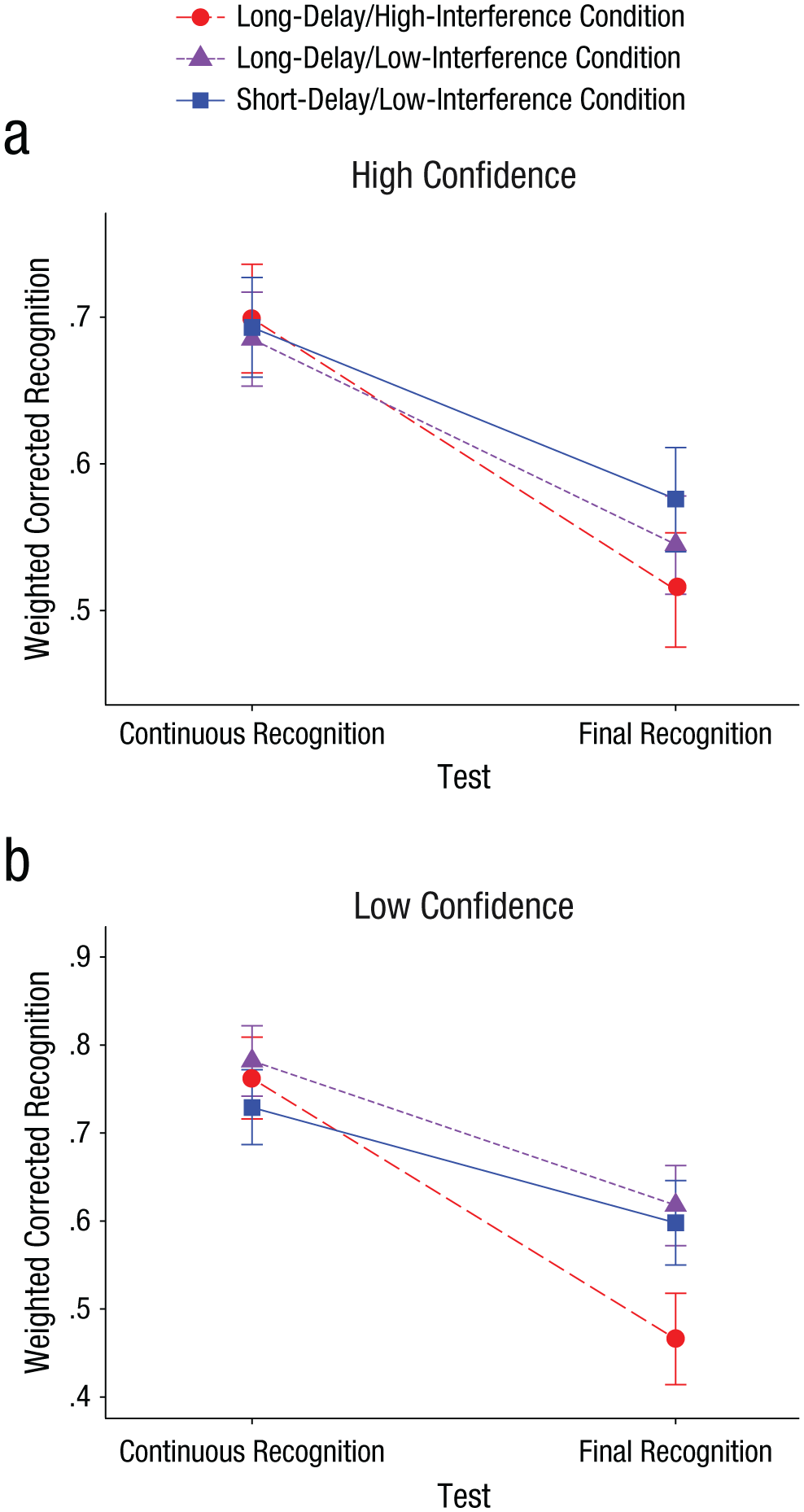
Corrected recognition as a function of test, weighted by high-confidence memories (a) and low-confidence memories (b), presented separately by condition. Error bars represent 95% confidence intervals around the means.
Analysis of Variance Results: Effects of Condition, Test, and Response on Weighted Corrected Recognition
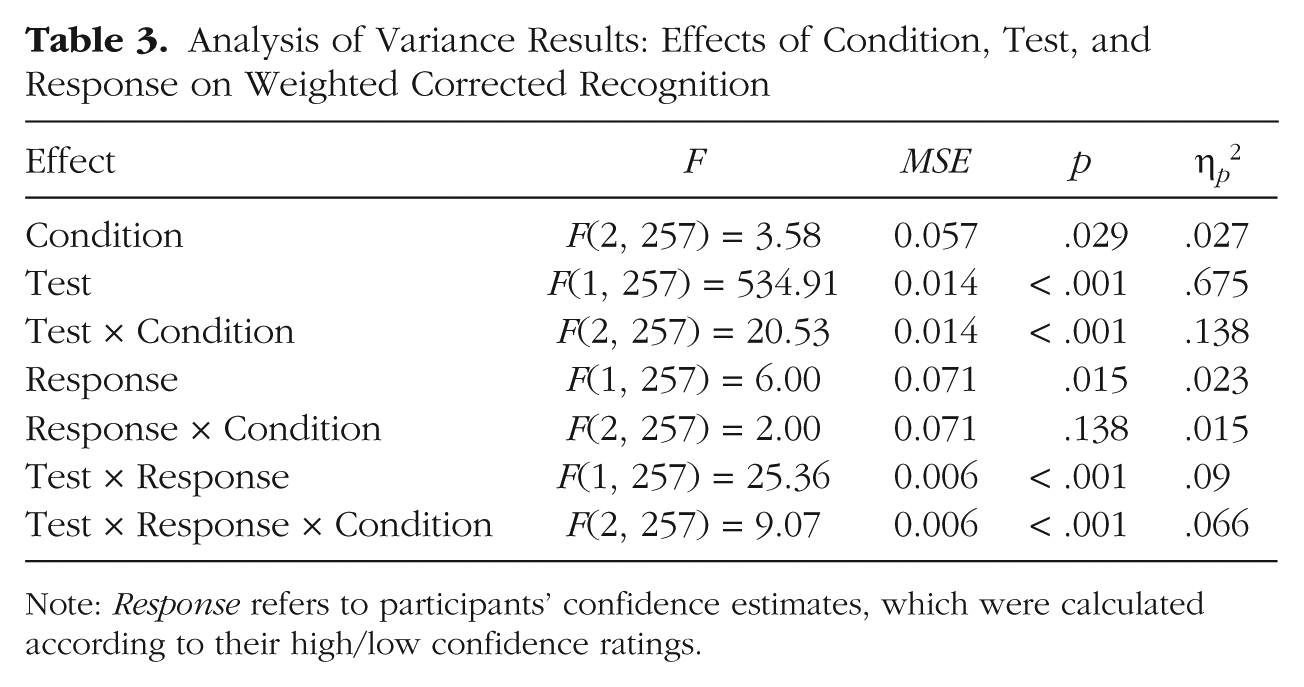
Note: Response refers to participants’ confidence estimates, which were calculated according to their high/low confidence ratings.
Note, however, that corrected recognition weighted by the high-confidence percentage was considerably larger than corrected recognition weighted by the recollection estimates because the percentage of high-confidence responses in the continuous recognition test was considerably higher than the percentage of recollection. As in the analysis of recollection estimates and familiarity estimates, we tested the union of the following three outcomes: (a) differential effects of decay and interference on high-confidence and low-confidence responses, (b) decay effect on high-confidence responses, and (c) interference effect on low-confidence responses. An analysis parallel to that described earlier resulted in a combined p value of less than .004. Last, as in the analysis of recollection estimates and familiarity estimates, our prediction that high-confidence responses would be less sensitive to interference than low-confidence responses was confirmed by a significant three-way interaction between condition (long-delay/high- interference, long-delay/low-interference), response (high confidence, low confidence), and test (continuous recognition, final recognition), F(1, 170) = 12.78, MSE = 0.08, p < .001, η p 2 = .07 (see the descriptive pattern in Fig. 3).
Because the responses in Phase 2 for all conditions were self-paced, we could not fully control for the duration of this stage. Thus, although we had aimed to equate the retention intervals of the long-delay/high-interference and long-delay/low-interference conditions, there was, in fact, a significant difference between them: The former lasted a mean of 15.46 min and the latter a mean of 13.82 min, t(170) = 2.85, p = .005, 95% confidence interval of the difference = [0.51, 2.78]). Therefore, to alleviate the concern that the effects we found were due to differences in duration between the two conditions, we reran the main analysis (the results of which are given in Figs. 2 and 3 and in Tables 2 and 3) and equated the durations of retention intervals between the two conditions. To this end, we excluded from the analysis 10 participants from the long-delay/high-interference condition for whom the duration of the retention interval was longer than 20 min. With these participants excluded from the analysis, there were no longer significant differences between the retention intervals of both conditions (p > .250). Note that the analysis examining the differential effects of decay and interference on recollection and familiarity estimates and on high- and low-confidence responses revealed the same pattern with the same significant effects, even after the exclusion of these participants.
Discussion
As we predicted, forgetting patterns depended on the type of memory representation. Relative to familiarity-based memories, recollection-based memories were more prone to decay than to interference. Likewise, relative to recollection-based memories, familiarity-based memories were more prone to interference than to decay. The pattern was the same for the other analysis: Relative to low-confidence memories, high-confidence memories were more prone to decay than to interference, and relative to high-confidence memories, low-confidence memories were more prone to interference than to decay. This pattern of results is all the more telling because, to date, most experimental manipulations (e.g., levels of processing, attention) have been found to exert their effects more strongly on recollection-based memories or high-confidence memories than on familiarity-based memories or low-confidence memories (Yonelinas, 2002). Our results thus provide the first direct and clear evidence for our representation theory of forgetting according to which the causes of forgetting depend on the nature of the underlying memory representation.
As mentioned in the introduction, there is a debate in the literature concerning the manner in which hippocampal and extrahippocampal memories are distinguished from each other. Whereas the distinction between recollection and familiarity emphasizes differences in the reliance on context between the two forms of memory, another alternative is that the differences relate to the strength of the memories. The purpose of the current study was not to distinguish between these alternatives but to maximize the potential to distinguish between hippocampal memories and extrahippocampal memories by using both approaches. Still, our findings support qualitative (rather than quantitative) differences between the two forms of memory, regardless of whether they are distinguished by the recollection/familiarity division or by confidence-ratings division. Quantitative differences, such as those we report, are consistent with certain strength models, according to which the pattern of activity elicited in the hippocampus at retrieval may be markedly different from that elicited in extrahippocampal structures. Specifically, a nonlinear relationship has been proposed between functional MRI-measured activity in medial-temporal-lobe structures and memory strength: In the hippocampus, there is a steep increase in activity at the higher levels of the strength scale, whereas for extrahippocampal structures, there is a steep increase in activity at the lower levels of the strength scale (Squire, Wixted, & Clark, 2007).
Our theory posits that recollection-based memories and high-confidence memories should be relatively protected from interference because of pattern separation in the hippocampus. Indeed, as our analyses focusing on the long-delay/high-interference and long-delay/low-interference groups show, interference exerts weaker effects on recollection-based memories than on familiarity-based memories, and it exerts weaker effects on high-confidence memories than on low-confidence memories. An interesting possibility is that these weaker effects of interference may, in fact, be the result of some recollected words converting to familiar words before the second phase of the experiment, whereas the rest of the words, which maintained their recollected status, are highly resistant to interference. This pattern could also be as true for high-confidence memories and low-confidence memories as it is for recollection-based and familiarity-based memories, respectively.
In our analysis, estimates of recollection and familiarity (as well as of confidence) were derived from the continuous recognition test. Our underlying assumption was that differences in memory between that continuous recognition test and the final test reflected the effects of the manipulations on these recollection and familiarity estimates. It is likely, however, that these estimates are slightly skewed because the continuous recognition test served as an additional encoding opportunity for each item, thereby possibly altering the nature of the items’ representations. Indeed, it has been found that manipulations that involve reencoding, including reminders (Hintzman, 2010), multiple presentations (Jacoby, Jones, & Dolan, 1998; Yonelinas, 2002), and multiple testing sessions (Chan & McDermott, 2007; Roediger & Karpicke, 2006), have noticeable effects on memory. In the context of the current experiment, it is possible that the continuous recognition test involved the addition of contextual information to an item classified as a familiar word, or, for other reasons, a conversion from low to high confidence. Conversely, as discussed in the previous paragraph, conversion of a recollected word to a familiar word (or a conversion from high confidence to low confidence) may have occurred. Unfortunately, we could not test for the potential effects of reencoding before the manipulation without introducing a test that would have involved yet another opportunity to reencode the items, and so on, ad infinitum.
Any change in the recollection estimate introduced by reencoding should not differ among the three conditions, because the estimation occurred before the manipulation (or, along the same lines, to the familiarity estimate or the confidence ratings). Nevertheless, we ran two simulations on our data to explore the possible effects of reencoding, the full details and results of which are reported in the Supplemental Material. The pattern of results in both simulations did not differ from the original data set in terms of differential effects of decay and interference on recollection and familiarity.
As mentioned, a plausible neural manifestation of decay in the hippocampus is the weakening of unstable memory traces, as is the case, for example, during consolidation or reconsolidation (by “consolidation,” we mean reorganization of neural systems supporting long-term memory retention; Hardt et al., 2013). On the other hand, consolidation and reconsolidation processes may also provide an opportunity for strengthening a memory trace. Thus, during consolidation or reconsolidation, items may be restored to their previous strength levels before decay sets in. This could possibly be the mechanism underlying testing effects (i.e., memory enhancement after testing; Roediger & Karpicke, 2006). Presumably, retesting offers an avenue for preserving memory from decay by strengthening the memory trace during reconsolidation. Notwithstanding this issue, what causes memory traces to either weaken or strengthen during consolidation and reconsolidation is an intriguing question for future research.
Finally, our results indicate that forgetting is not a singular concept; rather, it arises from both decay and interference. Although we showed that these two factors can be teased apart, it is important to note that interference and decay are inherently confounded. Interfering activity occurs over time and therefore necessarily includes a delay. By the same token, a delay period most likely involves interfering activity or thoughts. Nevertheless, by comparing memory performance across varying degrees of interference and decay, our study reveals that interference and decay differ with regard to the memory representations on which they exert their primary effects. These results provide evidence toward a resolution of the long-standing debate about interference and decay as causes of forgetting and support the representational theory of forgetting.
Footnotes
Acknowledgements
We are grateful to John Wixted for insightful comments and suggestions. We thank Erica Caron, Christa Dang, Catharina Englert, Nick Hoang, Sigal Gat-Lazer, and Xiaowei Wang for help in data collection. We thank John Anderson for technical assistance.
Action Editor
Stephen D. Lindsay served as action editor for this article.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by Natural Sciences and Engineering Research Council of Canada Grant A8347 (to M. Moscovitch), by Canadian Institutes of Health Research Grant MOP 125958 (to M. Moscovitch and G. Winocur), and by a Natural Sciences and Engineering Research Council of Canada Postdoctoral Fellowship award (to J. D. Ozubko).
Open Practices
All materials have been made publicly available via Open Science Framework and can be accessed at https://osf.io/tfhxm/. We are conducting additional analyses on the data set that are extraneous to the focus of the current article. We will make the raw data publicly available after these analyses are completed. The plan for Study 2 was registered at Open Science Framework and can be accessed at https://osf.io/ca5vt/. Results for that experiment, which was combined with Study 1 in this article, are reported separately in the Supplemental Material. The complete Open Practices Disclosure for this article can be found at http://pss.sagepub.com/content/by/supplemental-data. This article has received the badges for Open Materials and Preregistration. More information about the Open Practices badges can be found at https://osf.io/tvyxz/wiki/1.%20View%20the%20Badges/ and ![]() .
.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.