
Abstract
To investigate whether dogs could recognize contingent reactivity as a marker of agents’ interaction, we performed an experiment in which dogs were presented with third-party contingent events. In the perfect-contingency condition, dogs were shown an unfamiliar self-propelled agent (SPA) that performed actions corresponding to audio clips of verbal commands played by a computer. In the high-but-imperfect-contingency condition, the SPA responded to the verbal commands on only two thirds of the trials; in the low-contingency condition, the SPA responded to the commands on only one third of the trials. In the test phase, the SPA approached one of two tennis balls, and then the dog was allowed to choose one of the balls. The proportion of trials on which a dog chose the object indicated by the SPA increased with the degree of contingency: Dogs chose the target object significantly above chance level only in the perfect-contingency condition. This finding suggests that dogs may use the degree of temporal contingency observed in third-party interactions as a cue to identify agents.
Contingent reactivity—or, in other words, consistent and predictable relations between a subject’s actions and the occurrence of a partner’s responses—is an inherent feature of social interactions in both humans (Bigelow, 2001; Watson, 1979) and nonhuman animals (Carazo & Font, 2010). Matching the timing of actions has the potential to create a sense of connectedness among actions in both the interacting partners and the observers of those interactions. A high degree of temporal contingency is assumed to be an indicator of agency (Movellan & Watson, 2002), and it could also be crucial in the identification of communicative interactions (Csibra, 2010; Csibra & Gergely, 2006). In accordance with these accounts, 2-month-old human infants were found to react socially (with smiling and cooing) if a hanging toy above their crib moved contingently with their head movement on a pressure-sensitive pillow. This finding possibly indicates that they attributed agency to the mobile object (Watson, 1972). Moreover, after self-induced contingent interaction with an unfamiliar animated figure (Deligianni, Senju, Gergely, & Csibra, 2011), a robot (Movellan & Watson, 2002), or a furry puppet without a face (Johnson, Slaughter, & Carey, 1998), infants followed the orientation changes of these agents and looked at the distal target that was aligned with the front part of these agents. These findings have been explained as being due to the attribution of agency to these objects through the ascription of attention and perception (Johnson, 2000, 2003), and these results have also been used to argue that infants interpret the orientation change as a referential signal (Csibra, 2010).
However, little is known about whether nonhuman animals also use contingent reactivity as a cue to identify agency. Domestic dogs are promising candidates for studying this issue given that they have evolved a wide range of refined social skills (Hare & Tomasello, 2005). Dogs are sensitive to ostensive signals (e.g., eye gaze or hearing their own names) that might allow them to interact with humans (Topál, Kis, & Oláh, 2014). Converging evidence also shows that dogs are capable of correctly responding to several verbal commands (Kaminski, Call, & Fischer, 2004; Pilley, 2013; Pilley & Reid, 2011) and to nonverbal referential signals, such as pointing (Kaminski, Bräuer, Call, & Tomasello, 2009; Lakatos, Gácsi, Topál, & Miklósi, 2012) or gazing (Téglás, Gergely, Kupán, Miklósi, & Topál, 2012). However, human and canine interpretations of some of these signals are different (Kaminski et al., 2011; Tauzin, Csík, Kis, & Topál, 2015). Then again, these findings do not imply that dogs can identify the contingent structure of social interactions or that they can attribute agency on the basis of this information.
Although dogs’ previous experiences could help them to identify unfamiliar entities as agents (e.g., in a helping scenario; see A. Gergely et al., 2015), this does not necessarily entail that dogs attribute agency because they recognized the contingent reactivity of an unfamiliar agent. They might simply use other agency cues, such as goal directedness (G. Gergely & Csibra, 2003) or differences in the variability of motion (A. Gergely, Petró, Topál, & Miklósi, 2013). Thus, it remains unknown whether dogs are sensitive to the role that temporal contingency plays in interactions among agents and whether they can attribute agency on the basis of the observed contingent reactivity.
To investigate these questions, we adapted the method of Hare and Tomasello (1999). They found that dogs in a two-alternative forced-choice task preferred to choose the object that was approached by a human or a conspecific as opposed to the object not approached (for a review, see Miklósi & Soproni, 2006). We assumed that, in a similar setup, dogs would choose the object that had been approached by an unfamiliar, self-propelled agent (SPA) if this entity had been seen previously to take part in third-party contingent interactions because such interactions could elicit agency attribution.
Thus, in the present study, dogs were provided with different degrees of temporal contingency in three conditions. In the perfect-contingency condition, dogs heard verbal commands, each of which was consistently followed by a single response action of the SPA. It has been previously hypothesized that human infants can perceive high but imperfect degrees of contingency as communicative (Bigelow, 1998; Watson, 1994); therefore, in the high-but-imperfect-contingency condition, two thirds of the verbal commands were followed by action responses. In contrast, in the low-contingency condition, only one third of the verbal commands were followed by response actions. During the test phase, the SPA approached one of two identical tennis balls, thereby marking the target object.
The aim of this study was to test whether dogs could recognize contingent reactivity as a marker of third-party interactions and agency. More specifically, we investigated the effect of different degrees of temporal contingency on dogs’ tendency to attribute agency to an inanimate SPA, and we measured whether dogs showed an agent-congruent choice bias toward a target object. We assumed that if dogs attributed agency to an unfamiliar entity merely because of some morphological cues (eyespots, etc.) or self-propulsion, then they would exhibit a preference toward the target object in all conditions. However, if the observed degree of contingent reactivity was the primary indicator of agency, then dogs would show a significant agent-congruent preference in their choices when the degree of contingency was high enough.
Method
Subjects
Adult pet dogs (N = 74) and their owners were recruited from the Family Dog Research Database at the Department of Ethology, Eötvös Loránd University. 1 Two dogs in the low-contingency condition were excluded from the data analysis because they choose between the target objects in the test phase fewer than three times in the four trials. Additional dogs were excluded because of the technical failure of the radio-controlled car (4 in the perfect-contingency condition, 2 in the high-but-imperfect-contingency condition, and 2 in the low-contingency condition), because of experimenter error (1 in the high-but-imperfect-contingency condition), or because the owner did not follow the instructions (3 dogs, 1 in each of the three conditions).
The dogs that were included in the final analyses (N = 60) were randomly but evenly distributed over the three conditions, so that there were 20 dogs in each condition—perfect contingency: 11 males, mean age = 4.2 years, SD = 2.67 (4 mixed breeds and 16 purebreds from 13 different breeds); high-but-imperfect contingency: 10 males, mean age = 2.95 years, SD = 2.14 (3 mixed breeds and 17 purebreds from 10 different breeds); and low contingency: 12 males, mean age = 5.15 years, SD = 2.87 (7 mixed breeds and 13 purebreds from 10 different breeds). Sample size was defined a priori on the basis of previous research (for a review, see Miklósi & Soproni, 2006).
Audio stimuli
We used three different prerecorded verbal commands as cues in the observational-learning phase. The commands were recorded by a female assistant. The three commands were “Look, plush toy! Go!” (in Hungarian, “Figyelj, plüss! Indulj”), “Look, plush toy! Move back!” (“Figyelj, plüss! Tolass!”), and “Look, plush toy! Turn!” (“Figyelj, plüss! Fordulj!”). Note that although dogs have good spoken-word discrimination skills (e.g., see Griebel & Oller, 2012), the instructions we used in the present study (“go,” “turn,” and “move back”) are not part of a typical pet dog’s obedience-training vocabulary. Obviously, this fact does not exclude the possibility that these words were more or less familiar to some dogs. However, we investigated dogs’ sensitivity to temporal contingency; thus, their understanding of these instructions was not a precondition.
Commands were presented in series of three; each of the three types of commands (go, turn, and move back) was presented once within each triplet. To control for the order of commands, each of the six possible triplet orders (e.g., “go,” “turn,” “move back”; “go,” “move back,” “turn”) was presented once to each subject. The order of presentation of these six triplets was consistent across dogs and conditions. The presentation of verbal stimuli was preceded by a 16-s interval during which a ticking sound was presented (one tick per second). Then, a verbal command approximately 2 s long (“go”: 1.99 s; “turn”: 2.10 s; “move back”: 2.10 s) was played, followed by an 8-s break during which the previous ticking sound was presented. The dogs were presented with the commands and the breaks (duration: 9.99–10.1 s) 18 times altogether, with the same timing. However, the ticking sound was not presented in the last 8-s break; this indicated the end of the observational learning phase to the experimenters. The total duration of the audio-stimulus presentation, therefore, was 189.14 s: 16 s + (6 × 9.99 s) + (6 × 10.1 s) + (6 × 10.1 s) – 8 s.
Procedure
The experiment took place in a room (5.0 × 3.0 m) at the Department of Ethology, Eötvös Loránd University. A loudspeaker (SB-M300M2; Technics, Osaka, Japan; 33.5 × 21.4 × 38.6 cm) placed on the floor in one corner of the room was used for sound playback during the observational learning phase. Two 15-cm-tall plush toys, a crocodile (green) and an owl (beige), were used to demonstrate more contingent and less contingent interactions during the observational learning phase (see next section for more details). For half of the dogs, the crocodile was the SPA, and for the other half, the owl was the SPA. The toy acting as the SPA was mounted on a 16- × 10- × 10-cm radio-controlled car (U-Convertor; Nikko R/C, Tokyo, Japan).
Before the experiment started, the owner (who was present throughout the whole procedure) let the dog freely explore the testing room. Then the owner was asked to sit in a chair and to hold the dog by its collar; the dog sat in front of the owner, and both of them faced the loudspeaker (from a distance of 2.5–2.8 m), which was in the front right corner from the dog’s point of view.
Observational-learning phase
Experimenter 1 placed the SPA on the floor in the midline of the testing room, facing the loudspeaker, and placed the other plush toy directly in front of the loudspeaker. Thus, the two plush toys were approximately 1 m from each other. Experimenter 2, who stood to the left of the dog, placed an opaque occluder (1 × 0.75 m) between the dog and the setup each time Experimenter 1 moved or changed the setup. The occluder prevented the dog from observing the rearrangement of the objects. Then Experimenter 1, who controlled the SPA, stood in the front left corner of the room, opposite the dog, with his head facing down, and Experimenter 2 removed the occluder. Owners were not allowed to speak throughout the procedure.
In the perfect-contingency condition, the SPA responded to the 18 prerecorded verbal commands (six command triplets) by performing the corresponding action during the 8-s breaks. That is, in response to the command “Look, plush toy! Go!” the agent moved approximately 30 cm straight forward (i.e., toward the speaker and the other toy) and returned to the starting point. In response to the command “Look, plush toy! Move back!” the SPA moved 30 cm back and then straight forward on the same route. Finally, in response to the command “Look, plush toy! Turn!” the SPA turned 45° to the right while proceeding forward (again moving approximately 30 cm); this movement resulted in an arc-like trajectory. The SPA then moved back to its starting point on the same trajectory. There was no delay between the cue and the response beyond the natural variance caused by the latency of Experimenter 1 and the radio-controlled car, which was approximately the same in all conditions (0.2–0.4 s) because Experimenter 1 and the apparatus were the same in each condition.
The high-but-imperfect-contingency condition was similar to the perfect-contingency condition except that the SPA moved in response to only two thirds of the command playbacks. That is, on one third of the trials, the SPA ignored the command; it did not respond to the command playbacks and remained motionless during the 8-s breaks. On one third of the trials, the SPA responded to the auditory signal by performing a single action. On the remaining third of the trials, the SPA performed two different actions. Note that the number and the order of verbal commands, as well as the number and order of actions performed by the SPA, were the same as in the perfect-contingency condition. Consequently, correspondence among commands and actions was not perfect.
The dogs observed while the six command triplets were played and the SPA responded or did not respond to the commands. In each triplet, there were commands with 0, 1, or 2 response actions. The order of vocal commands with 0, 1, or 2 response actions was counterbalanced within subjects. Consequently, the dogs were presented with six possible combinations of 0, 1, and 2 response actions in a predetermined order (0, 1, 2; 0, 2, 1; 1, 0, 2; 1, 2, 0; 2, 0, 1; 2, 1, 0). On the basis of previous findings with human infants (Deligianni et al., 2011; Johnson et al., 1998; Movellan & Watson, 2002) and theoretical claims about agency attribution (Watson, 1994), we supposed that most social agents—in contrast to physical objects—produce variable and delayed responses. Therefore, beyond response uncertainty, the delay between the uttering of the command and the action of the SPA was either 0, 1, 2, 3, or 4 s (in addition to the aforementioned response latency of Experimenter 1 and the SPA) during the 8-s break between commands. The average delay was 1.16 s (SD = 0.22).
The low-contingency condition was basically the same as the perfect-contingency condition except that the SPA responded only to every third verbal cue (i.e., on two thirds of the trials, the SPA ignored the command and remained motionless). However, in the trials in which the SPA did respond to the command playbacks, it performed each of the three corresponding actions (i.e., go, move back, turn) during the 8-s breaks without intentional delay between the cues and responses. The number and order of the commands and actions were the same as in the other two conditions. This ensured that the number and the variability of actions were similar to those presented in the perfect-contingency and in the high-but-imperfect-contingency conditions (see Fig. 1).
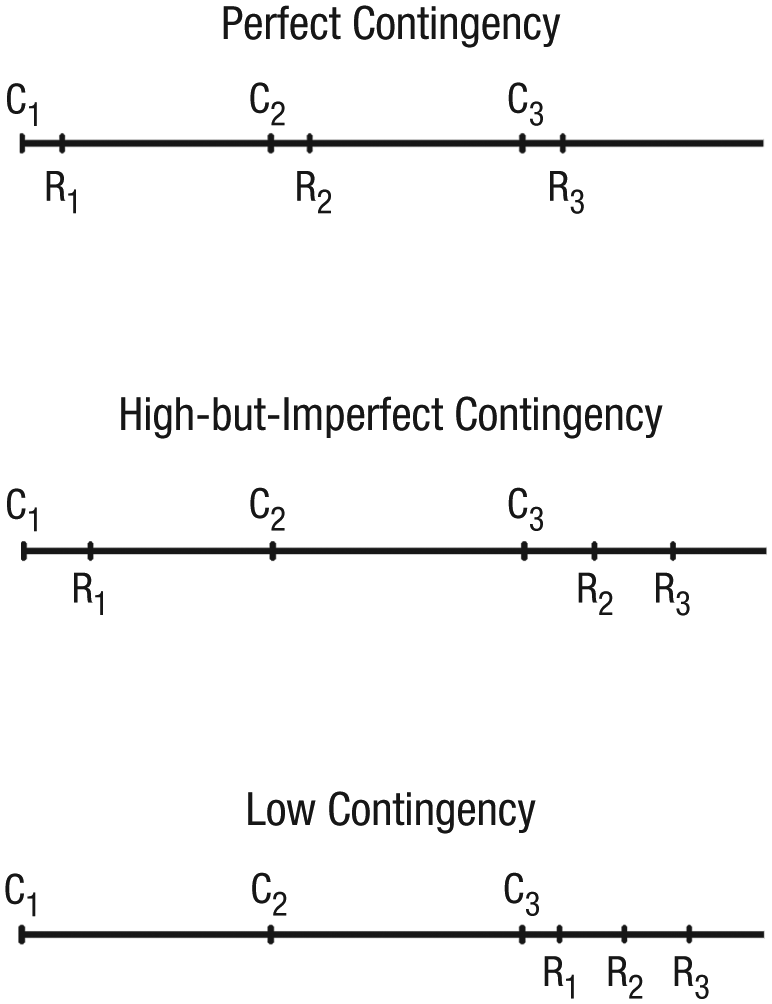
Examples of temporal patterns of commands (C) and responses (R) for the three conditions.
Test phase
The observational-learning phase was followed immediately by the test phase. Experimenter 2 placed the occluder between the dog and the setup; behind the occluder, Experimenter 1 placed two identical tennis balls on the floor 2 m apart. The tennis balls were equidistant (2 m) from the dog. Experimenter 1 also placed the SPA on the floor halfway between the two tennis balls, facing toward one of the balls (i.e., the target object). Experimenter 2 removed the occluder, and then the SPA indicated the target object by approaching it in a straight line. The SPA stopped before it could bump into the target object so that it was marked but without manipulation. After that, the occluder was placed again between the dog and the apparatus. Experimenter 1 removed the SPA and then left the room.
Finally, Experimenter 2 removed the occluder, the owner released the dog, and the dog made its choice. After choosing, the dog was allowed to play with the selected ball for a few seconds, then the owner called the dog and made it sit down. The whole procedure was repeated four times in total; the side of the target object was counterbalanced in an ABBA order (see Fig. 2).
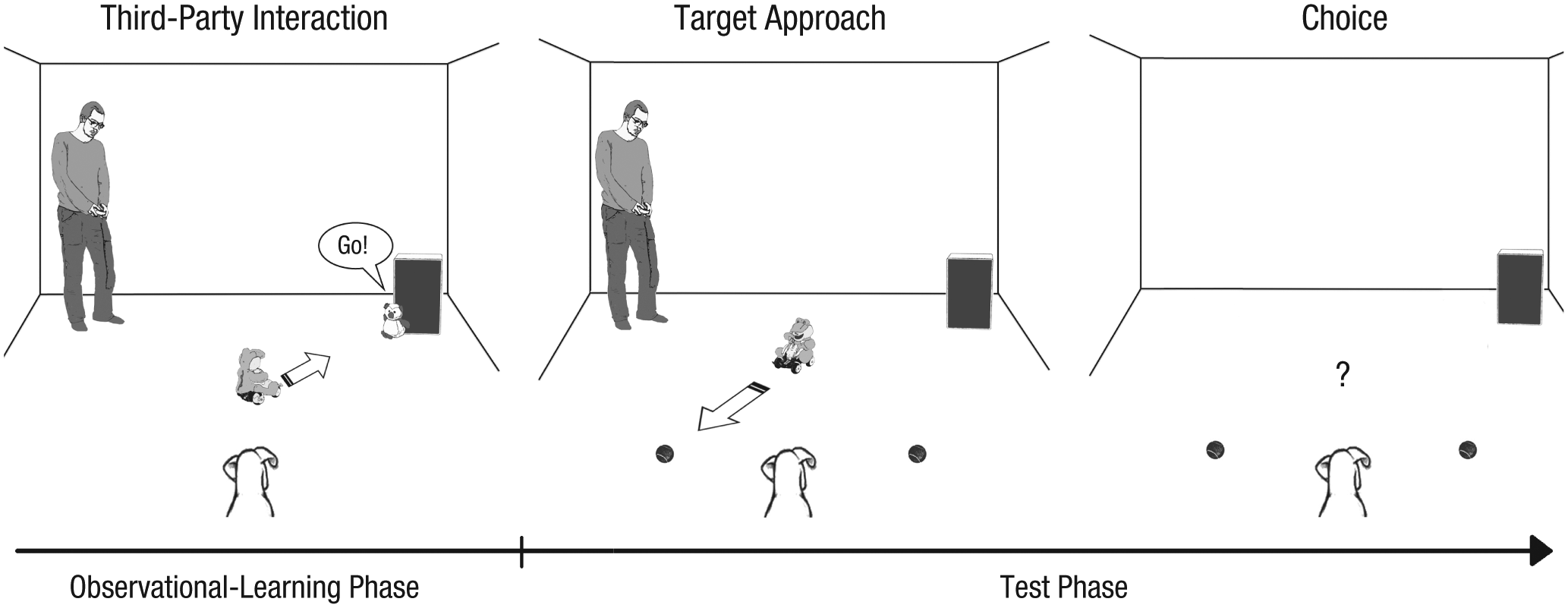
The observational-learning and test phases of the experiment from the dog’s point of view. In the observational-learning phase, the dogs observed that the self-propelled agent (SPA) responded to the verbal commands in a more or less contingent manner (depending on the condition); in the test phase, the dogs observed the SPA approach one tennis ball, and then they chose one of the tennis balls.
Data collection and analysis
Test events were videotaped and analyzed later. A tennis ball was regarded as chosen if the dog touched it or the dog’s paw or muzzle was closer than approximately 0.15 m to the ball within 30 s. All of the test-trial videotapes were first coded by author T. Tauzin; a second coder blind to the demonstration also coded all of the test trials. Interrater reliability was perfect (Cohen’s κ = 1.0 in all conditions).
We included in the analyses all dogs that made a choice on at least three of the four trials. We used the proportion of trials on which the dog selected the target object to measure each dog’s performance. To compare the dogs’ performance with chance-level performance, we used Wilcoxon signed-rank tests; to compare the performance of dogs in different groups, we used Mann-Whitney U tests. To test whether the degree of contingency could predict the dogs’ performance in the test phase, we performed a simple linear regression analysis. We used IBM SPSS Statistics (Version 21) to conduct our analyses.
Results
In the perfect-contingency condition, the dogs chose the target object (i.e., the tennis ball indicated by the SPA) significantly more often than expected by chance (z = −2.44, p = .016, r = .39). However, the dogs did not choose the target object significantly more often than expected by chance in the high-but-imperfect-contingency condition (z = −1.72, p = .117, r = .27) or in the low-contingency condition (z = −0.44, p = .800, r = .07). Moreover, the mean proportion of trials on which the dog chose the target object was higher in the perfect-contingency condition (M = 67.08, SD = 26.96, 95% CI = [55.27, 78.90]) than in the high-but-imperfect-contingency condition (M = 60.00, SD = 23.50, 95% CI = [49.70, 70.30]) or in the low-contingency condition (M = 48.75, SD = 24.96, 95% CI = [37.81, 59.69]). The difference between the perfect-contingency and low-contingency conditions was significant (U = 124, p = .037, r = .34). However, the difference between the perfect-contingency condition and the high-but-imperfect-contingency condition (U = 174.5, p = .458, r = .02) was not significant; also, the difference between the high-but-imperfect-contingency condition and the low-contingency condition (U = 141.5, p = .101, r = .26) was not significant (Fig. 3). A linear regression revealed a significant effect of the level of contingency on the frequency with which the dogs chose the target object, β = 0.094, t(58) = 2.39, p = .020, R2 = .089. More details about the dogs’ choices are available at Open Science Framework (http://osf.io/b6xv4).
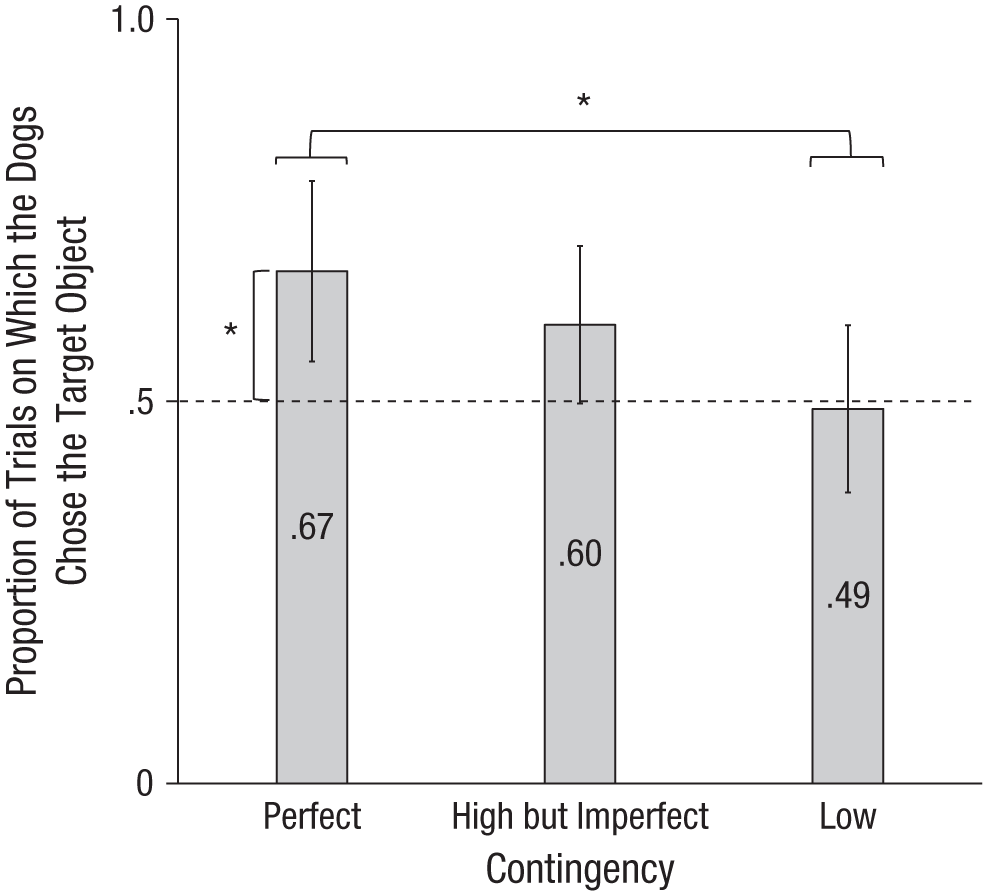
Proportion of trials on which the dogs chose the target object in each condition. An asterisk indicates a significant difference (p < .05) between conditions or from chance (i.e., .5, indicated by the dashed line). Error bars represent 95% confidence intervals.
Discussion
We hypothesized that dogs would attribute agency to the SPA and that, consequently, they would be biased to choose the target object as a function of the previously observed temporal contingency. Our results support this hypothesis, because the dogs’ performance was affected by the different degrees of temporal contingency. They chose the tennis ball that was approached by the SPA significantly above chance level only in the perfect-contingency condition, and they chose the target object significantly more often in the perfect-contingency condition than in the low-contingency condition. Moreover, the finding that the dogs showed intermediate performance in the high-but-imperfect-contingency condition suggests that they were sensitive to the differences in the degree of contingency as well.
The present finding also supports the notion that dogs tend to perceive an object’s perfectly contingent motor responses to auditory cues as an interaction; in the current study, this perception would have had the potential to elicit agency attribution to the SPA. However, this finding does not necessarily imply that the dogs interpreted the SPA’s target-approaching action referentially (i.e., they did not interpret the action as being similar to pointing or gazing) or that the dogs selectively attributed communicative intent to the SPA. It is reasonable to assume that the dogs in the test phase simply paid more attention to agents than to nonagents and that the varying degrees of choice bias across conditions stemmed from the dogs’ selective attention to the contingently responding SPA, which could direct their attention to the target object through local enhancement (Thorpe, 1956). Note that our results also indicate that the dogs’ behavior in the three conditions cannot be interpreted purely in terms of the attention-grabbing potential of self-propelled objects (Hoppitt & Laland, 2013), because such an account would imply a significant target-object preference in the low-contingency condition as well. We therefore assume that the dogs’ performance in the present study reflected their sensitivity to third-party contingency, which made them capable of attributing agency to self-moving entities.
Although the dogs did not show a significant choice bias in the high-but-imperfect and low-contingency conditions, the observed degree of contingency significantly predicted their test performance in these conditions according to the results of the linear regression model. However, a conclusion that there is a linear relationship between dogs’ choice bias and the observed degree contingency would be premature, because the dogs were tested in only three different conditions. This pattern of results might mean that in an observational context, dogs could interpret a dyadic event as interactive and attribute agency to the interactive objects if the event involved very high degrees of temporal contingency. Human infants, in contrast, attribute agency on the basis of high but imperfect contingency in social situations (Watson, 1994) and have a preference toward middle or high contingencies in nonsocial contexts (Kidd, Piantadosi, & Aslin, 2012, 2014). This difference may be rooted in species-specific sets of innate abilities in dogs and humans (e.g., language-related skills in humans). However, it is plausible that, in addition to these innate factors, ontogenetic factors also have an effect on the expectations about communicative interactions in dogs. Because pet dogs are quite often exposed to human commands, they might simply find the perfect-contingency condition the most similar to the command-response interactions in which pet dogs often take part. In contrast, human infants are supposed to form expectations on the basis of less predictable, conversation-like interactions (Bigelow, 2001; Watson, 1979, 1994). Note, however, that even if the dogs’ previous experience shaped their reactions in the present study, the results imply that they are sensitive to the different degrees of contingency and that they attribute agency on that basis.
In summary, our findings support the notion that dogs, like infants (Bigelow, 1998, 2001), can grasp some fundamental perceptual characteristics of interactions as well as the importance of contingency in them. This suggests that dogs are sensitive to the temporal structure of events that can indicate to the dogs the presence of agents, which might mean that dogs’ social skills are more similar to those of humans than previously supposed (Hare & Tomasello, 2005). Thus, besides their ability to correctly respond to an agents’ gazing (Téglás et al., 2012), pointing (Kaminski et al., 2009; Lakatos et al., 2012) or to verbal commands (Kaminski et al., 2004; Pilley, 2013; Pilley & Reid, 2011), dogs might be able to identify an agent whose behavior should be monitored, even if that agent is unfamiliar to them. Agent identification, therefore, might help dogs to differentiate between social and nonsocial interactions, a skill that could also help them to determine whether an action is relevant.
However, it is crucial to note that dogs might use these cues differently than humans. There is some indication that, in dogs, the degree of contingent reactivity has a linear relation with agency attribution; consequently, dogs attribute agency if the agent shows perfect or near-perfect contingent reactivity. In contrast, in humans, a nonlinear (i.e., inverted-U-shaped) function is supposed to describe the same relation; thus, humans attribute agency when the degree of contingency is high but imperfect (Watson, 1994). Therefore, the superficial similarity between the two species in the sensitivity to contingencies might involve a crucial difference that can reflect an interspecies divergence in the representation of agents or in the use of contingency as an agency cue. As opposed to humans, whose representations about the presence or absence of agency are more categorical, dogs might represent level of agency as a continuous function of the degree of contingency.
Footnotes
Acknowledgements
We thank Andor Csík, Fruzsina Elekes, Gabriella Felhősi, Ágnes Melinda Kovács, Ádám Miklósi, Eszter Szabó, and John Watson for their help, and we are grateful to all participating dog owners.
Action Editor
Gretchen B. Chapman served as action editor for this article.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This research was supported by Hungarian Science Foundation Grants OTKA K112138 and OTKA NK83997 and by the European Research Council (ERC) under the European Union’s Seventh Framework Programme (FP7/2007-2013)/ERC Grant 609819.